

Abstract
Genes located on the mammalian Y chromosome outside of the pseudoautosomal region do not recombine with those on the X and are predicted to either undergo selection for male function or gradually degenerate because of an accumulation of deleterious mutations. Here, phylogenetic analyses of X-Y homologues, Zfx and Zfy, among 26 felid species indicate two ancestral episodes of directed genetic exchange (ectopic gene conversion) from X to Y: once during the evolution of pallas cat and once in a common predecessor of ocelot lineage species. Replacement of the more rapidly evolving Y homologue with the evolutionarily constrained X copy may represent a mechanism for adaptive editing of functional genes on the nonrecombining region of the mammalian Y chromosome.
The eutherian Y chromosome is unusual because conventional recombination with the X is limited to genes within the small pseudoautosomal region. Comparative analyses of mammalian sex chromosomes suggest that the Y acquires genes through recombination with the X in the pseudoautosomal region, and that these genes subsequently undergo extensive repositioning into the nonrecombining region of the Y (NRY) via inversion and intrachromosomal translocation (1, 2). There, X-Y recombination ceases, and genes in the NRY are predicted to either become specialized for male function (3, 4) or gradually degenerate from the accumulation of deleterious mutations through processes such as Muller's ratchet (5), hitchhiking with favorable mutations at other Y genes (6), background selection (6), and insertion of retroposons (7).
However, additional factors may influence the genetic composition and evolution of genes within the NRY. For example, during hominid evolution 4–5 million years ago, a major portion of the X long arm was incorporated onto the Y (8) and subsequently was repositioned into the NRY via intrachromosomal recombination between repetitive LINE (long-interspersed element) retroposons (9). This is evidence that the occasional translocation of large chromosome fragments into the NRY is possible. Another example is Zfx and Zfy, a pair of gene homologues located in the sex-specific regions of chromosomes X and Y, respectively. A sequence comparison of coding regions of these genes between rodent and primate taxa (10–12) revealed lower than expected divergence estimates between these X and Y homologues. Thus, some form of genetic exchange was indicated in the NRY.
Here we address this potential evolutionary paradox by a comprehensive sequence analysis of the terminal Zn-finger exon of Zfy and Zfx in 26 species of the cat family Felidae. Unusual patterns of diversification between these homologues demonstrate a novel mechanism of genetic exchange, termed ectopic gene conversion, occurs within the nonrecombining regions of the X and Y. Because the felid phylogeny has been well established by using multiple molecular genetic markers (13–20), we were able to characterize precisely two distinct episodes of ectopic gene conversion of Zfy by Zfx during the evolution of modern-day cat species. Considered together with a reassessment of Zfy and Zfx diversification across mammalian orders represented by human, rat, and cat, we offer additional insights on the evolutionary fate of functional genes within the NRY.
Materials and Methods
DNA Specimens.
DNA was purified from either blood or skin cell fibroblasts from a male and female from 26 species of felids representing the eight major evolutionary groups. Species from the ocelot lineage included ocelot (Leopardus pardalis), tigrina (L. tigrinas), margay (L. weidii), pampas cat (Lynchailurus colocolo), Geoffroy's cat (Oncifelis geoffroyi). Representatives from the domestic cat lineage were domestic cat (Felis catus), jungle cat (F. chaus), and European wildcat (F. silvestris). Within the Panthera group, lion (Panthera leo), leopard (P. pardus), tiger (P. tigris), jaguar (P. onca), and clouded leopard (Neofelis neofelis) were included. Three species comprising the puma group were puma (Puma concolor), cheetah (Acinonyx jubatus), and jaguarundi (Herpailurus yagouaroundi). The lynx genus included bobcat (Lynx rufus) and Siberian lynx (Lynx lynx). The Asian leopard cat group was Asian leopard cat (Prionailuris bengalensis), flat-headed cat (Ictailurus planiceps), and fishing cat (P. viverrinus). The caracal group was represented by caracal (Caracal caracal), African golden cat (Profelis aurata) and serval (Leptailurus serval). One species, Asian golden cat, (Profelis temmincki) was used from the bay cat group. Lastly, the phylogenetically unaligned species of pallas cat (Otocolobus manul) was included in the analysis. GenBank accession numbers for exons in human ZFY and ZFX were J03134 and M30607. Rat Zfy and Zfx exon sequences were X75172 and X75171, respectively. Intron GenBank accession numbers were X58926 and X58925 for human ZFY and ZFX. Intron sequences from rat were X58934 and X58933 for Zfy and Zfx, respectively. Domestic cat intron sequences are available at http://lgd.nci.nih.gov.
PCR Amplification of Exon.
Amplification of the Zn-finger exon used chromosome-specific sets of primers (Table 1). The same approach was repeated for both Zfy and Zfx whereby an initial segment (generated by chromosome-specific primers) encompassing a portion of the 5′ adjacent intron and spanning the exon was used as the template in nested PCR. For Y genes, chromosome specificity was verified by the presence of the Zfy gene PCR product in male individuals and absent from female individuals for each felid species. For Zfy, two primer pairs were used for first-round PCR: Zfy2F/olw111r, δZfyF/Exon6R (Table 1). Selecting the appropriate first-round product as template, a series of overlapping Zfy fragments were amplified by several primer pairs: Zfy1F/Exon1R, Zfy1F/Exon4R, δZfyF/Exon4R, Exon4F/Exon6R. Likewise, Zfx was generated by using nested PCR with first-round products amplified by primer pairs Zfx2F/δZfxR and δZFxF/olw143R. Second-round nested PCR for Zfx used the appropriate first-round Zfx product as template and the primer pairs of Zfx1F/δZfxR and δZfx1F/Exon6R. Other primer pairs (Table 1) were used as needed for additional sequence amplification and verification.
Table 1.
List of primers, gene specificity, nucleotide position, sequence, and source
| Primer | Gene | Position | Nucleotide position | 1st Round | 2nd Round | Sequence | Source |
|---|---|---|---|---|---|---|---|
| Zfy2F | Zfy | 5′ Intron | −459–439 | X | 5′-tgtcagcataagcaggct | 2 | |
| Zfy1F | Zfy | 5′ Intron | −166–149 | X | 5′-tggagtttgctgttacct | 2 | |
| Zfx2F | Zfx | 5′ Intron | −731–717 | X | 5′-gcttctgtgactga | 2 | |
| Zfx1F | Zfx | 5′ Intron | −403–386 | X | 5′-tgggtttaagctctactg | 2 | |
| Zf2F | Zfy/x | 5′ Intron | −804–783 | X | 5′-ttccaggcagtac | 2 | |
| δZfyF | Zfy | Exon | 292–315 | X | X | 5′-tgcataaggaaaaagccaaca | 1 |
| δZfxF | Zfx | Exon | 292–315 | X | X | 5′-tgcataaggagaaaggagccaaca | 1 |
| δZfyR | Zfy | Exon | 315–292 | X | 5′-tgttggctttttctttatgca | 1 | |
| δZfxR | Zfx | Exon | 315–292 | X | X | 5′-tgttggctcctttctccttatgca | 1 |
| Exon4F | Zfy/x | Exon | 885–900 | X | 5′-cgccatattctctcag | 1 | |
| Exon4R | Zfy/x | Exon | 900–885 | X | 5′-ctgagagaatatggcg | 1 | |
| olw111F | Zfy/x | Exon | 1005–1028 | X | 5′-agtgtaccagtgtgagtactgtgagtat | 3 | |
| olw111R | Zfy/x | Exon | 1028–1005 | X | 5′-atactcacagtactcacactgata | 3 | |
| Exon5F | Zfy/x | Exon | 1057–1071 | X | 5′-ggcacgttatctcca | 1 | |
| Exon5R | Zfy/x | Exon | 1071–1057 | X | 5′-tggagataacgtgcc | 1 | |
| Exon6R | Zfy/x | Exon | 1181–1164 | X | X | 5′-gggcaggccaacttcctt | 1 |
| olw144 | Zfy | 3′ Flank | (+)195–165 | X | 5′-agaaactgaataggattcagaacagcg | 3 | |
| olw143 | Zfx | 3′ Flank | (+)195–165 | X | 5′-aacaaactgaatagaattcagaacacac | 3 |
Conditions for PCR amplification in the first stage consisted of 50 μl reactions containing 50–100 ng/μl total genomic DNA, 50 mM KCl, 10 mM Tris (pH 8.3), 1.5 mM MgCl2, 0.01% gelatin, 0.01% NP-40, 0.01% Tween-20, 0.2 μM of each primer, 0.2 mM dNTP, and 2.5 units Taq polymerase. For Zfy, thermocycling conditions for the first round consisted of a hot start of 3′ at 94°C, followed by 40 cycles of 15 sec at 94°C, 15 sec at 48°C, and 30 sec of 72°C and ending with a final extension of 72°C for 5 min. The second round consisted of a 2 μl volume of the first-round product with PCR conditions identical to the first round, except for an annealing temperature of 52°C. With Zfx exon amplification, conditions were identical except that the annealing temperature was 55°C for both first and second rounds of nested PCR. All PCRs were performed with an ABI Perkin-Elmer 9700 thermocycle sequencing machine.
PCR products were visualized on a 1% agarose gel. Sequences were generated by BIGDYE sequencing kits (Applied Biosystems) and analyzed by using an automated sequencer (model 373; Applied Biosystems).
Sequence Analyses.
Sequences encompassing the final exon of Zfy and Zfx were aligned by using the algorithm of Needleman and Wunsch (21) with the GCG computer package version 8 (22) and verified visually. The complete exon multiple sequence alignment is deposited at the web site of http://lgd.nci.nih.gov. Additional alignments were derived for the genetic analysis of cat, human, and mouse Zfy and Zfx sequences by using intron and exon sequences. The alignment of intron sequences used clustal x (23). Nucleotide substitution models were selected based on empirically derived estimates of the following parameters: frequency of nucleotides, transition/transversion ratio, and among-site rate variation. Therefore, exon genetic distances used the Kimura two-parameter model of substitution (24). Intron sequences were analyzed by using Tajima-Nei (25) distances. Exon sequences were translated into amino acids by using mega version 1.01 (26).
Phylogenetic Analysis.
Phylogenetic analysis of exon sequences in Felidae was performed by paup* (27). Specific conditions for the heuristic search are presented in Fig. 1. The reliability of the data to recapitulate the same tree was assessed by a bootstrap analysis. In the cat-human-rat phylogenetic analyses of exon and intron segments, exhaustive searches that examined all possible trees to identify the most optimal phylogeny were implemented by paup* with specific conditions presented in Fig. 3.
Figure 1.

Phylogenetic analysis using 1,181 bp of the Zn-finger exon of Zfx and Zfy for 26 species of Felidae. Shown is the 50% majority rule consensus (length = 171; consistency index = 0.672) of 10,850 trees generated by maximum parsimony analysis. Numbers on branches are length/homoplasies. Numbers in italics are bootstrap proportions in support of adjacent node that exceeded 50%. Bootstrap analysis consisted of 100 iterations (43). Note that the Zfx and Zfy sequences are respectively monophyletic with the exception of the pallas cat Y, which resembles X chromosomes Zfx genes and more precisely, the pallas cat Zfx homologue (see text). The resolution of species divergence hierarchies within the Zfx and Zfy groups are not robust because of slow-moving evolution of these genes in the period of Felidae evolution (10–15 million years). The derived tree is significantly different from a constrained tree representing established species lineages as listed in Fig. 2 (Kishino-Hasegawa test; t = 2.59; P < 0.0095). For this reason, the species topologies should not be considered to reflect Felidae phylogenetic divergence accurately.
Figure 3.

(A) Phylogenetic tree of exon sequences from cat, human, and rat Zfx and Zfy genes. The tree is derived from minimum-evolution using neighbor-joining derived from an exhaustive search using Kimura 2P model. Branch lengths are % sequence divergence. (B) Phylogenetic tree of adjacent 5′ intron of Zfy and Zfx from cat, human, and rat. Shown is the minimum evolution-neighbor joining tree derived from an exhaustive search of all possible trees based on the Tajima-Nei model of substitution. Numbers on branches represent % sequence divergence.
Expression of Zfy and Zfx.
Nine tissues were dissected from a euthanized male domestic cat kitten and flash-frozen into liquid nitrogen. Total RNA was extracted from frozen tissue each from kidney, liver, thymus, spleen, heart, lung, skin, brain, and testes by using the protocol of Rneasy minikit (Qiagen, Chatsworth, CA). cDNA was synthesized by using the procedure of Superscript Preamplification System (GIBCO/BRL). Hemi-nested PCR was used for the amplification of cDNA sequences by using primers ZF1F and Exon1R for the first round. Second-round primers consisted of primers ZF2F and the Y-specific primer δZfyR for Zfy, and ZF2F with X-specific primer δZfxR for Zfx. These primers span the final intron and 315 bp into the final exon. In both rounds, each 50-μl reaction contained 50–100 ng/μl total cDNA, 50 mM KCl, 10 mM Tris (pH 8.3), 1.0 mM MgCl2, 0.01% gelatin, 0.01% NP-40, 0.01% Tween-20, 0.2 μM of each primer, 0.2 mM dNTP, and 1.25 units of Taq polymerase. Using a Perkin–Elmer 9700 thermocycling machine, first-round cycling conditions consisted of an initial hot start of 3′ at 94°C, followed by 40 cycles of 15 sec at 94°C, 15 sec at 50°C, and 30 sec at 72°C followed by a final extension of 5′ at 72°C. Second-round conditions were identical except the annealing temperature was changed to 52°C.
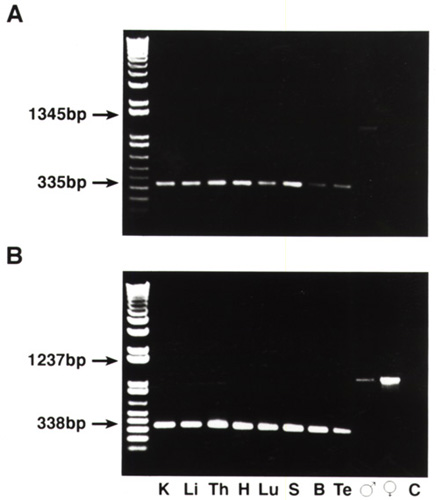
This same fragment, using the primers listed above for the cDNA second round, was amplified from genomic DNA for Zfy and Zfx. Each 50-μl reaction used the same concentration of reagents, with the appropriate sex-specific primer pair, but adjusted MgCl2 between 1.0 mM for Zfx and 2.5 mM for Zfy. Thermocycling conditions were identical to those used above with following exceptions: the annealing temperature was 50°C for Zfx and 60°C for Zfy; and amplification occurred with 35 cycles rather than 40. Genomic and cDNA were identified by their expected differences in product size (338 bp cDNA, 1,237 bp genomic DNA for Zfx, and 335 bp cDNA versus 1,345 bp genomic for Zfy).
Results
We sequenced the terminal Zn finger exon (1,181 bp) of Zfx and Zfy from individuals of 26 Felidae species to extend previous research on the molecular evolution of X and Y genes. Our initial analysis (20) of the adjacent 5′ intron of Zfx and Zfy among cat species revealed differences in nucleotide substitution rates between X and Y chromosome introns and demonstrated that these genes recapitulated previously described felid phylogenetic relationships. In contrast, the present study indicated that Zfx and Zfy exons evolved more slowly compared with intron sequences and were phylogenetically uninformative in establishing expected relationships among cat species (Fig. 1). Nonetheless, there was considerable genetic distance between Zfx and Zfy sequences supporting the notion that Zfx/Zfy divergence predated the evolution of modern-day felids. An unusual exception to this Zfx/Zfy exon sequence distinctiveness occurred in one species, pallas cat, where Zfy clustered with pallas cat Zfx within the monophyletic lineage of all other Zfx genes (Fig. 1). It was this unexpected observation that stimulated the following analysis.
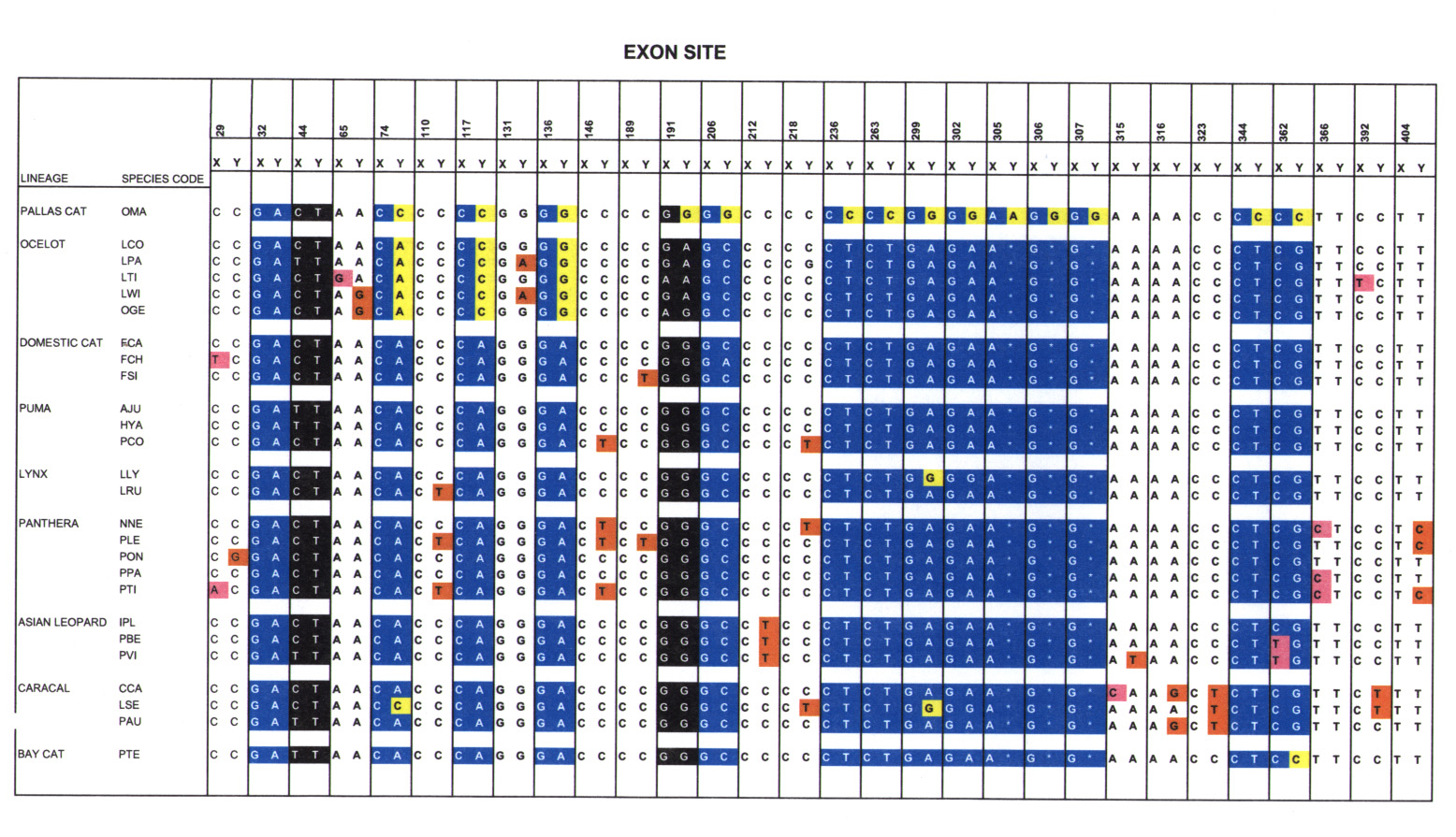
Alignment of Zfx and Zfy exons revealed 55 nucleotide site positions that were variable in at least two species of Felidae (Fig. 2). Of these, several (19/55) represented fixed differences that discriminated between Zfx and Zfy exons (blue). For example, site 32 was diagnostic as G (Zfx) or A (Zfy) in all cat species (see Fig. 4, which is published as supplementary material on the PNAS web site, www.pnas.org). Therefore, these substitutions likely occurred in a common ancestor for all modern felids. A second category of substitutions (5/55) included sites that exhibited the same polymorphisms in both Zfy and Zfx exons (black). For instance, site 512 alternated randomly between A and G in both Zfx and Zfy across Felidae (see Fig. 4).
Figure 2.

Distribution of nucleotide sites that vary in two or more felid species in the Zn-finger exon of Zfy and Zfx. Except for the phylogenetically unaligned pallas cat, all other species are listed within the eight monophyletic evolutionary groups within Felidae (11–18). Five categories of substitutions are depicted. Blue: substitution diagnostic between Zfy and Zfx and fixed in the majority of species from all lineages. Black: Zfx-Zfy polymorphism, not sex specific. Orange: Newly derived Zfy substitution shared by some, or all, of the corresponding species group. Pink: Newly derived Zfx substitution shared by some, or all, of the corresponding species group. Yellow: Site where the Zfy-specific nucleotide is replaced by that from Zfx in some, or all of the corresponding species group. The yellow continuum for pallas cat includes those sites that are identical between Zfy and Zfx in most felid species (including pallas cat). In addition, the clearest evidence of gene conversion is that each of the fixed diagnostic sites (blue) shared by all other felid lineages are not present in the yellow region of Zfy exons from pallas cat or ocelot lineage species. Caracal group includes serval. β = Substitution that is unique and shared by Zfy and Zfx within a single species. *: Sites that encode amino acid substitutions distinguishing between Zfy and Zfx in all felids. For specific alignment details of polymorphic sites, please see Fig. 4 at PNAS web site http://www.pnas.org and http://lgd.nci.nih.gov.
A continuum of yellow blocks over multiple, adjacent sites (Fig. 2) indicated a historic gene conversion event where the Zfy gene segment was replaced by an exact copy of the counterpart sequence from the Zfx exon. Two such motifs were apparent in Zfy (Fig. 2), one spanning sites 74–1046 in pallas cat and the second in positions 74–136 in the five ocelot lineage species. In the pallas cat, with the exception of the first four variable sites (sites 29, 32, 44, 65), the Zfy exon was identical to the corresponding Zfx exon. For example, position 236 was Zfx-C and Zfy-T in all species except in pallas cat where both Zfx and Zfy were C. This pattern is repeated throughout the pallas cat Zfy fragment starting from site 74. For this reason, the direction of gene conversion was clearly from Zfx to Zfy and not vice versa. The second putative gene conversion event occurred in a common ancestor of the monophyletic ocelot lineage. In this case, the motif encompassed approximately 62 bp (positions 74–136). Three of the sites within this region (positions 74, 117, and 136) were diagnostic between Zfy and Zfx in all other species of Felidae. However, within the ocelot lineage the Zfy sequence at these sites was identical to Zfx and indicated directed gene conversion from X to Y. In addition, two of these species, ocelot (Lpa) and margay (Lwi) exhibited a derived Y substitution that evolved after the initial gene conversion event.
The greater number of shared derived substitutions (synapomorphies) observed for Zfy (24/55) than Zfx (9/55) indicated unequal rates of nucleotide substitution between Y and X homologues during Felidae evolution. Excluding the ocelot lineage species and pallas cat, the empirical substitution rate ratio estimated between Zfy and Zfx exons was Y/X = 1.96. Using the equation Y/X = 3 αm/(αm + 2), which accounts for the pattern of inheritance between Y and X chromosomes (28), the estimated male/female mutation rate ratio (αm) was 3.76. This value was consistent with the hypothesis of male-driven evolution that states enhanced rates of mutation observed in Y genes are a consequence of the higher number of cell divisions that take place in spermatogenesis versus oogenesis (28).
Translation of nucleotides into amino acid residues indicated the protein domain encoded by the Zn finger exon (393 aa) was highly conserved between Zfy and Zfx in Felidae. Of the 55 sites examined, only three (positions 117 and 136 and a 3-bp deletion of 305–307), encoded amino acid changes between Zfy and Zfx in all felid species. Two of these sites, 117 and 136, were converted from the Zfy version (117 isoleucine; 136 histidine) to the Zfx version (117 leucine; 136 arginine) in both pallas cat and a common ancestor of the ocelot lineage. The third amino acid difference, encoded by the nucleotide insertion/deletion at sites 305–307, resulted in a lysine deletion from Zfy in all felids. However, in pallas cat, the gene conversion of Zfy by Zfx resulted in the restoration of lysine at this site in Zfy.
Zfx and Zfy Expression in Domestic Cat.
To assess the tissue distribution of Zfx and Zfy in cats, we extracted RNA from nine tissues of domestic cat, prepared cDNA, and assayed for Zfx/Zfy transcripts by using reverse transcription–PCR methodology. The results show abundant RNA transcripts for both genes in every tissue (Fig. 5, which is published as supplementary material on the PNAS web site). Because the PCR primers spanned an intron of both Zfy (995 bp) and Zfx (844 bp), the cDNA predicted size of 338 bp (Zfx) and 335 bp (Zfy) were easily distinguished from genomic DNA product of 1,237 bp for Zfx and 1,345 bp for Zfy (see Fig. 5). DNA sequence analysis confirmed each cDNA PCR product as Zfy or Zfx.
Genetic Distance of Zfx and Zfy in Human, Mouse, and Cat Homologues.
Provocative insights on sex chromosome evolution relative to gene evolution were derived by sequence comparison of Zfy and Zfx in different mammalian orders. In Table 2 (which is published as supplementary material on the PNAS web site), we present genetic distances between Zfx and Zfy gene sequences in human, rat, and domestic cat species with available intron and exon data. Considered together with phylogenetic trees of these exon and intron sequences (Fig. 3), three principle observations were revealed. First, intron and exon sequences of Zfx diverged more slowly than their respective counterparts from Zfy. Most likely, this was a reflection of the reduced substitution rates for X chromosome genes. Second, genetic distances (exon or intron) between cat and human for both Zfx and Zfy genes (i.e., cat Zfx vs. human Zfx and cat Zfy vs. human Zfy) were considerably closer than the comparable distance of either gene from either species to rat Zfx and Zfy. This difference might be explained by the more ancient divergence of rodents (estimated at 112 million years ago) in comparison to the split between carnivores and primates (approximately 80 million years ago) (29). Third, distance estimates between Zfx and Zfy in cat (1.8% for exon and 29.8% for intron) are less than Zfy divergence among mammal orders. The same is true for human (i.e., X-Y distances are 5.3% for exon and 40.1% for intron). Thus, the closest sequence to cat and human Y sequence was its corresponding X homologue within each order. Because the original placement of Zfx and Zfy onto the sex chromosomes occurred early within eutherian mammal divergence (2, 30, 31), this similarity implied ancient episodes of gene conversion too old to allow character analysis such as is presented in Fig. 2. By contrast, the rat does not display this increased X-Y similarity. Our analysis was unable to determine whether the rat Zfy and Zfx results were a consequence of a lower frequency of conversion, generation time differences (32), or sampling errors.
Discussion
Comparative analysis of the terminal Zn-finger exon of Zfy and Zfx in 26 species of Felidae depicts a novel form of genetic exchange between mammalian sex chromosomes. Located in the sex-specific regions, and therefore excluded from conventional recombination during chromosomal pairing of X and Y, these gene homologues demonstrate unique, precise episodes of ectopic gene conversion. The proposed mechanism of ectopic gene conversion is thought to be an inadvertent outcome of homology searches preceding, and essential to, synapsis of homologous chromosomes (33). Thus, ectopic gene conversion may be particularly relevant to nonrecombining regions of the genome like the NRY because it does not require the formation of the synaptonemal complex accompanied by reciprocal exchange (34, 35) and also may take place during both meiosis and mitosis (36–38). Considered together with our comparisons of Zfy and Zfx segments between carnivore (cat), primate (human), and rodent (rat), our results illustrate the potential importance of ectopic gene conversion in the maintenance of Y gene function in eutherian mammals.
Using a phylogenetic analysis of Zfy and Zfx exons, the presence of ectopic gene conversion was indicated by the unexpected placement of the pallas cat Zfy sequence clearly within the monophyletic lineage of Zfx sequences from all other felid species. Additional analysis of variable sites within aligned Zfy and Zfx sequences confirmed gene conversion of nearly the entire Zfy exon by that of Zfx in pallas cat and also revealed a second event involving the replacement of a smaller fragment (62 bp) in multiple species within the ocelot lineage.
The frequency of fixation of ectopic conversion events between sex chromosomes can be estimated by using the well-characterized evolutionary history of the Felidae. The ocelot lineage and pallas cat results indicate ectopic gene conversion plus fixation occurred twice in the evolution of nine lineages tested. As the divergence of modern felid species likely occurred 10–15 million years ago (18–20), the frequency of fixation for ectopic gene conversion is 0.0148 events/million years. Assuming an average generation time of 5 years in felids, an estimate of 7.4 × 10−8 ectopic gene conversion events/generation is obtained. This estimate reflects only those events that become fixed within the species and is thus a fraction of number of ectopic gene conversion possible within each generation. Therefore, it is consistent with the estimate of 1 × 10−7/generation, computed from experimental studies of mitotic ectopic gene conversion not associated with crossing over in yeast (39).
The observed X to Y direction of gene conversion (Fig. 2) suggests that a more constrained version of the gene, preserved by comparatively lower nucleotide substitution rates of the X chromosome, is reintroduced into the paternal lineage. Genes on the X chromosome evolve more slowly than those on the Y, presumably because of male-driven evolution [i.e., sex-biased mutation rates arising from germ-cell division differences (28, 40)] or selection leading to a mutation rate reduction on the X chromosome (41). In felids, the estimated ratio of male/female mutation rate per generation for Zn-finger exon is αm = 3.76. This is consistent with the estimated ratio for the adjacent intron (αm = 4.38) (20). Considered together, these estimates support the prediction of higher mutation rates of Y chromosome genes compared with those from the X.
Ectopic gene conversion may prove to be a mechanism for adaptive correction of Y genes whose X homologues do not undergo X inactivation in females. Unlike the mouse, in which Zfy is expressed in testes only (42), domestic cat and humans express Zfy and Zfx ubiquitously, implying both copies of the gene are required for normal function. Yet, genes located in the nonrecombining region have a greater probability of accumulating deleterious mutations and undergoing loss of function. The predicted enhanced mutation rates on the Y are supported by our data that depict a greater number of newly derived substitutions in Zfy (orange) relative to Zfx (pink) during felid radiation (Fig. 2). Of those that were nonsynonymous, three encoded diagnostic amino acid differences between Zfy and Zfx in all felids. The subsequent removal of these differences in the ocelot lineage (two of three sites) and pallas cat (three of three sites) via ectopic gene conversion reverts Zfy to a more conserved state. Therefore, the apparent nonrandom distribution of gene conversion sites relative to codon-altering substitutions may indicate targets of selective pressures responsible for driving gene conversion events to fixation.
Our results revise and expand previous research on the evolution of genes located in the NRY of the Y chromosome. Our reassessment of substitution patterns in the Zn finger exon across mammalian orders of carnivore, primate, and rodent consistently depicts a closer association between Zfy and its corresponding Zfx homologue. Under the hypotheses that these two homologues diverged early within eutherian radiation, the expected phylogenetic pattern should have demonstrated a major bifurcation between a monophyletic lineage composed of Zfy sequences and one of all Zfx. However, the opposite is observed whereby Zfy and Zfx cluster with each other within each order. Thus, a process of ancestral recombination likely occurred that may have consisted of the following: (i) ancient reciprocal exchange occurred between these homologues originally in the pseudoautosomal region, (ii) intrachromosomal shuffling of Zfy into the nonrecombining region at different times in each mammalian order, and (iii) episodes of ectopic gene conversion that occurred after the genes were situated within the sex-specific regions of Y and X. Because of this process, ancient recombination patterns will be evident only by comparisons across higher-order taxonomic divisions. In contrast, more recent evolutionary trends can be discerned by phylogenetic analysis of Zfy and Zfx homologues within mammalian orders. In Felidae, we find evidence for both processes. Zfy and Zfx separate into monophyletic gene-specific lineages that reflect recent diversification and precise episodes of ectopic gene conversion between these genes in modern species.
In conclusion, based on the evolution of Zfy and Zfx in multiple species of Felidae, we postulate a plausible third alternative for the fate of genes located in the NRY. The conventional view that most of the Y chromosome, excluding the pseudoautosomal region, cannot benefit from recombination is challenged by the clear incidence of ectopic gene conversion in felid evolution. This form of recombination may be an effective mechanism for the adaptive editing of functional genes within the NRY and thus ameliorate the effects of Y chromosome degeneration.
Supplementary Material
Acknowledgments
We thank A. J. Pearks-Wilkerson and S. Cevario for excellent laboratory assistance. We thank our colleague Dr. Warren Johnson for helpful comments and discussions. We acknowledge the National Cancer Institute for allocation of computer time and assistance at the Frederick Biomedical Supercomputing Center. All tissue samples were collected in full compliance with specific Federal Fish and Wildlife permits, Convention and International Trade in Endangered species of Wild Flora and Fauna (CITES); Endangered and Threatened Species, Captive Bred issued to the National Cancer Institute-National Institutes of Health (S.J.O., principal officer) by the U.S. Fish and Wildlife Service of the Department of the Interior.
Abbreviation
- NRY
nonrecombining region of the Y
Footnotes
References
- 1.Ellison J W, Li X, Francke U, Shapiro L J. Mamm Genome. 1996;7:25–30. doi: 10.1007/s003359900007. [DOI] [PubMed] [Google Scholar]
- 2.Lahn B, Page D. Science. 1999;286:964–967. doi: 10.1126/science.286.5441.964. [DOI] [PubMed] [Google Scholar]
- 3.Graves J A M. BioEssays. 1995;17:311–321. doi: 10.1002/bies.950170407. [DOI] [PubMed] [Google Scholar]
- 4.Graves J A M. J Exp Zool. 1998;281:472–481. [PubMed] [Google Scholar]
- 5.Charlesworth B, Charlesworth D. Genet Res. 1997;70:63–73. doi: 10.1017/s0016672397002899. [DOI] [PubMed] [Google Scholar]
- 6.Charlesworth B. Curr Biol. 1996;6:149–162. doi: 10.1016/s0960-9822(02)00448-7. [DOI] [PubMed] [Google Scholar]
- 7.Charlesworth B. Science. 1991;251:1030–1033. doi: 10.1126/science.1998119. [DOI] [PubMed] [Google Scholar]
- 8.Page D C, Harper M E, Love J, Botstein D. Nature (London) 1984;311:119–123. doi: 10.1038/311119a0. [DOI] [PubMed] [Google Scholar]
- 9.Schwartz A, Chan D C, Brown L G, Alagappan R, Pettay D, Disteche C, McGillivray B, de la Chapelle A, Page D. Hum Mol Genet. 1998;7:1–11. doi: 10.1093/hmg/7.1.1. [DOI] [PubMed] [Google Scholar]
- 10.Hayashida H, Kuma K, Miyata T. J Mol Evol. 1992;35:181–183. doi: 10.1007/BF00183228. [DOI] [PubMed] [Google Scholar]
- 11.Pamilio P, Bianchi N O. Mol Biol Evol. 1993;10:271–281. doi: 10.1093/oxfordjournals.molbev.a040003. [DOI] [PubMed] [Google Scholar]
- 12.Shimmin L C, Chang B H-J, Li W-C. J Mol Evol. 1994;39:569–578. doi: 10.1007/BF00160402. [DOI] [PubMed] [Google Scholar]
- 13.Collier G E, O'Brien S J. Evolution. 1985;39:437–487. doi: 10.1111/j.1558-5646.1985.tb00389.x. [DOI] [PubMed] [Google Scholar]
- 14.Modi W S, O'Brien S J. In: Chromosome Structure and Function. Gustafson J P, Arpels R, editors. New York: Plenum; 1988. pp. 215–242. [Google Scholar]
- 15.Pecon Slattery J, Johnson W E, Goldman D, O'Brien S J. J Mol Evol. 1994;39:296–305. doi: 10.1007/BF00160153. [DOI] [PubMed] [Google Scholar]
- 16.Janczewski D, Modi W S, Stephens J S, O'Brien S J. Mol Biol Evol. 1995;12:690–707. doi: 10.1093/oxfordjournals.molbev.a040232. [DOI] [PubMed] [Google Scholar]
- 17.Masuda R M, Lopez J V, Pecon Slattery J, Yuhki N, O'Brien S J. Mol Phylogenet Evol. 1996;6:351–365. doi: 10.1006/mpev.1996.0085. [DOI] [PubMed] [Google Scholar]
- 18.Johnson W E, Dratch P A, Martenson J S, O'Brien S J. J Mamm Evol. 1996;3:97–120. [Google Scholar]
- 19.Johnson W E, O'Brien S J. J Mol Evol. 1997;44:S98–S116. doi: 10.1007/pl00000060. [DOI] [PubMed] [Google Scholar]
- 20.Pecon Slattery J, O'Brien S J. Genetics. 1998;148:1245–1255. doi: 10.1093/genetics/148.3.1245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Needleman S B, Wunsch C D. J Mol Evol. 1970;48:443–453. doi: 10.1016/0022-2836(70)90057-4. [DOI] [PubMed] [Google Scholar]
- 22.Genetics Computer Group. Program Manual for the Wisconsin Package Version 8. Madison, WI: Genetics Computer Group; 1994. [Google Scholar]
- 23.Thompson J D, Gibson T J, Plewniak F, Jeanmough F, Higgins D G. Nucleic Acids Res. 1997;25:4876–4882. doi: 10.1093/nar/25.24.4876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kimura M. J Mol Evol. 1980;16:11–120. doi: 10.1007/BF01731581. [DOI] [PubMed] [Google Scholar]
- 25.Tajima F, Nei M. Mol Biol Evol. 1984;1:269–285. doi: 10.1093/oxfordjournals.molbev.a040317. [DOI] [PubMed] [Google Scholar]
- 26.Kumar S, Tamura K, Nei M. mega: Molecular Evolutionary Genetics Analysis. University Park: Pennsylvania State Univ.; 1993. , Version 1.0. [Google Scholar]
- 27.Swofford D L. paup*: Phylogenetic Analysis Using Parsimony (*And Other Methods) Sunderland, MA: Sinauer; 1998. , Version 4. [Google Scholar]
- 28.Miyata T, Hayashida H, Kuma K, Mitsuyasu K, Yasanaga T. Cold Spring Harbor Symp Quant Biol. 1987;52:863–867. doi: 10.1101/sqb.1987.052.01.094. [DOI] [PubMed] [Google Scholar]
- 29.Kumar S, Hedges B. Nature (London) 1998;392:917–918. doi: 10.1038/31927. [DOI] [PubMed] [Google Scholar]
- 30.Sinclair A H, Foster J W, Spencer J W, Page D C, Palmer M, Goodfellow P N, Graves J A M. Nature (London) 1988;333:780–783. doi: 10.1038/336780a0. [DOI] [PubMed] [Google Scholar]
- 31.Watson J M, Frost C, Spencer J A, Marshall Graves J A. Genomics. 1993;15:317–322. doi: 10.1006/geno.1993.1063. [DOI] [PubMed] [Google Scholar]
- 32.Li W-H, Graur D. Molecular Evolution. Sunderland, MA: Sinauer; 1999. [Google Scholar]
- 33.Steele D F, Morris M E, Jinks-Robertson S. Genetics. 1991;127:53–60. doi: 10.1093/genetics/127.1.53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Jinks-Robertson S, Sayeed S, Murphy T. Genetics. 1997;146:69–78. doi: 10.1093/genetics/146.1.69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.McKim K S, Green-Marroquin B L, Sekelsky J J, Chin G, Steinberg C, Khodosh R, Hawley R S. Science. 1998;279:876–878. doi: 10.1126/science.279.5352.876. [DOI] [PubMed] [Google Scholar]
- 36.Jinks-Robertson S, Petes T D. Genetics. 1986;114:731–752. doi: 10.1093/genetics/114.3.731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Lichten M, Goldman A S H. Annu Rev Genetics. 1995;29:423–444. doi: 10.1146/annurev.ge.29.120195.002231. [DOI] [PubMed] [Google Scholar]
- 38.Hogstrand K, Bohne J. Mol Biol Cell. 1997;8:2511–2517. doi: 10.1091/mbc.8.12.2511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Petes T D, Hill C W. Annu Rev Genet. 1988;22:147–168. doi: 10.1146/annurev.ge.22.120188.001051. [DOI] [PubMed] [Google Scholar]
- 40.Haldane J B S. Ann Eugen. 1947;13:262–271. doi: 10.1111/j.1469-1809.1946.tb02367.x. [DOI] [PubMed] [Google Scholar]
- 41.McVean G T, Hurst L D. Nature (London) 1997;386:388–392. doi: 10.1038/386388a0. [DOI] [PubMed] [Google Scholar]
- 42.Koopman P, Ashworth A, Lovell-Badge R. Trends Genet. 1991;7:132–136. doi: 10.1016/0168-9525(91)90458-3. [DOI] [PubMed] [Google Scholar]
- 43.Hillis D M, Bull J J. Syst Biol. 1993;42:182–192. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}
{kind=link}
{kind=link}