

Abstract
Climate differences across latitude can result in seasonal constraints and selection on life history characters. Since Aedes albopictus (Skuse) invaded North America in the mid-1980s, it has spread across a range of ≈14° latitude and populations in the north experience complete adult mortality due to cold winter temperatures that are absent in the south. Life table experiments were conducted to test for differences in the adult survival and reproductive schedules of Ae. albopictus females from two populations from the northern (Bloomington, IN [BL] and Manassas, VA [VA]; ≈39° N) and southern (Tampa, FL and Fort Myers, FL; ≈27–28° N) extremes of the species distribution in North America. Regardless of population origin, age-specific hazard rate increased with reproductive output and decreased with number of bloodmeals. Larger females took fewer bloodmeals, and they had greater hazard rates than did smaller females. There were no consistent differences between northern versus southern populations in resource allocation between reproduction and maintenance, reproduction over time, and reproductive investment among offspring, suggesting that latitudinal variation in climate is probably not a main selective factor impinging on adult mortality and reproductive schedules. One possible effect of climate on geographic differences in life history was detected. BL had lower survivorship, lower lifetime reproductive output, and lower adult reproductive rate than did all other populations. This result may be an indirect result of lower egg survivorship due to the severity of winter in BL compared with other populations, including VA at approximately the same latitude. Such a scenario may make the BL population more prone to extinction, irregularly recolonized from more favorable sites, and thus more susceptible to founder effects, genetic drift, and inbreeding, resulting in lower mean values of fitness-related traits.
Keywords: life history evolution, survival, fecundity, reproductive investment
The Asian tiger mosquito, Aedes albopictus (Skuse), is considered medically important because it is a vector of arboviruses, including dengue, La Crosse encephalitis, and West Nile virus (Ibañez-Berñal et al. 1997, Gerhardt et al. 2001, Turell et al. 2005). Although its native range is centered on the Oriental region and India, Ae. albopictus has invaded North and South America, Europe, and Africa in the past two decades (see Juliano and Lounibos 2005, and references therein) to represent a “third wave” of worldwide spread of human disease-vector mosquitoes, after two prior waves by Aedes aegypti (L.) in the sixteenth and seventeenth centuries and Culex quinquefasciatus Say in the nineteenth century (Lounibos 2002). It is increasingly clear that understanding the processes involved in the invasion and spread of Ae. albopictus is crucial for accurate assessment of new health risks (Lounibos 2002).
Of fundamental importance to the invasion and spread of an organism are the temporal patterns of reproduction and survival that make up its life history (Juliano and Lounibos 2005). In many instances, energy available to organisms is limiting; thus, trade-offs occur among traits when energy is partitioned among growth, maintenance, and reproduction. Trade-offs can manifest as costs of reproduction, such as decreases in parental longevity or reduced ability to reproduce later in life with early reproduction (Bell 1980). Fundamental trade-offs are also likely to occur within the reproductive budget, including negative relationships between fecundity and investment per offspring. Because of these trade-offs among fitness-related traits, natural selection favors individuals with trait combinations that attain greatest fitness under local regimes of mortality and reproductive phenology (Michod 1979, Bell 1980, Roff 2002).
Intraspecific variation in life history traits across latitude has long been a focus in insect ecology (Stearns 1992, Roff 2002). Individuals from unfavorable or harsh environments may inherently commit a high proportion of available resources (e.g., time and energy) to reproduction to ensure that at least some offspring reach maturity, despite high mortality, resulting in relatively high lifetime reproductive output. For species that inhabit a range of climates, early reproduction could increase the fitness of individuals in regions subject to seasonal time constraints, such as onset of an unfavorable climate (e.g., Rowe and Ludwig 1991). Unfavorable environments also may favor production of higher investment (i.e., larger) offspring because greater offspring size often increases offspring probability of surviving to adulthood (Fox and Czesak 2000). Thus, individuals from populations at different latitudes may become locally adapted via selection on allocation of resources to survival, current reproduction, and future reproductive investment among offspring. In this study, we test for reproductive life history differentiation and physiological costs of reproduction for four populations of Ae. albopictus from the extremes of the species’ latitudinal range in North America.
Ae. albopictus invaded continental United States from Japan through a port in Houston Texas in the mid-1980s (Sprenger and Wuithiranyagool 1986). Since then, it has spread rapidly eastward and northward over a range of ≈14° latitude, and it has become one of the most common human-biting mosquitoes in its new range (Moore 1999, Lounibos et al. 2002). In the eastern United States, the northern range limit of the species corresponds approximately to daily mean January temperatures of −5°C (Nawrocki and Hawley 1987, Hanson and Craig 1995), indicating severe winter temperatures. In contrast the southern limit of the species in North America includes the southernmost counties of Florida (O’Meara et al. 1995), which have a subtropical climate. In temperate zones, Ae. albopictus experiences high larval and almost complete adult mortality during winter and almost exclusively survives adverse winter temperatures as diapausing eggs (Sota and Mogi 1992, Swanson et al. 2000), whereas in subtropical areas, larvae and adults are often active during the winter (Rios and Maruniak 2004; Lounibos and Escher 2008). In the Northern Hemisphere, the duration of favorable summer temperatures permitting survival of adults and larvae declines with increasing latitude. Hence, high larval, and more importantly, almost complete adult mortality due to cold winter temperatures in the north are absent in the south, where Ae. albopictus is likely limited by factors impinging solely on immature survival, such as interspecific competition or predation at the larval stage and the desiccation and thermal tolerance of eggs (Juliano et al. 2002, Kesavaraju et al. 2008).
Ae. albopictus has shown differentiation of life history traits subsequent to its invasion of North America. Armbruster and Conn (2006) found weak but consistent trends toward increased rates of larval growth traits in northern relative to southern populations from Florida, Texas, and New Jersey. Lounibos et al. (2003) showed the incidence of diapause in Ae. albopictus eggs differed among populations over a smaller latitudinal range, from Jacksonville to Miami, FL, suggesting more rapid differentiation. Isozyme analyses conducted shortly after the eastward and northward range expansion of Ae. albopictus in the United States were interpreted as indicating rapid local differentiation (Black et al. 1988). Similar patterns of isozyme variation were detected at the heart of its native range in the tropics, suggesting that local genetic differentiation is characteristic of this species (Black et al. 1989). Hence, many North American populations seem sufficiently isolated in climatically different habitats to allow for local adaptation if appropriate selection exists. Components of adult fitness are important aspects of life history. In this study, we expand on earlier work to investigate geographic differentiation of adult longevity and reproduction of North American Ae. albopictus.
Life History Predictions for Ae. Albopictus
Intraspecific variation in life history traits across latitude has long been a focus in ecology because climate differences across latitude can result in seasonal constraints and selection on life history characters. Populations at the northern and southern extremes of the North American distribution of Ae. albopictus are within temperate versus subtropical zones respectively, and they are likely to experience vastly different climates. However, no prior studies that we are aware of have evaluated the role of interpopulation variation in the adult life history of North American Ae. albopictus. We used life table experiments to compare survival, longevity, preoviposition reproductive investment (blood-feeding), and postoviposition reproductive allocation of Ae. albopictus from two populations from the northern and southern extremes of the species distribution in North America. We also test for the effect of body size on the longevity and postoviposition reproductive allocation of Ae. albopictus.
Greater seasonal mortality of the adult stage is predicted to select for greater reproductive investment when the average contribution of each offspring to adult fitness increases (Michod 1979). Reproduction has survival costs, but greater investment in reproduction (particularly early reproduction) may outweigh these costs if seasonal constraints are severe. For Ae. albopictus and other blood-feeding mosquitoes, higher reproductive investment is likely to manifest in two nonexclusive ways: increased blood intake (i.e., number and size of bloodmeals) and a greater total mass of eggs (i.e., the product of increased individual egg mass and number of eggs). Blood feeding by mosquitoes represents a phenotypic expression of reproductive investment. It is the acquisition of resources specifically for reproduction and can increase the risk of death for the female mosquito (Roitberg et al. 1993) and diverts time and energy from maintenance. Reproductive output represents the energy allocated to egg production and oviposition that could otherwise be allocated to maintenance of somatic function, and a risk to survival during the act of oviposition. We predict that Ae. albopictus from northern populations will invest greater time and energy in blood feeding and thus take more and larger bloodmeals. Furthermore, after controlling for both body size and number of bloodmeals, we predict that females from northern populations will produce a greater total mass of eggs.
Unfavorable climatic effects that decrease the survival of both adult and juvenile mosquitoes are also likely to alter how reproduction is allocated over time and among offspring. For individuals constrained by a short active season, early onset of reproduction is likely to increase fitness despite associated costs of reduced longevity and residual reproductive output. Conversely, for individuals that experience a longer active season greater reproductive output may result from apportioning energy to reproduction more evenly over time. We predict that Ae. albopictus from northern populations will have greater early reproductive investment, which is likely to manifest in three ways: increased size of initial bloodmeals, decreased time until first oviposition, and greater egg production in the first gonotrophic cycle. Greater juvenile mortality also is predicted to select for higher investment offspring, specifically larger eggs at the cost of fewer of these eggs. Producing fewer larger eggs may increase the number of offspring that successful complete development in a short season and ultimately to increase the likelihood of reaching a diapause state in which successful overwintering is possible. We therefore predict that Ae. albopictus from northern populations will produce fewer, larger eggs than will Ae. albopictus from southern populations.
Materials and Methods
Collection and Maintenance of Mosquitoes
Laboratory colonies of Ae. albopictus were established from larvae and pupae collected from each of four discrete field sites in the United Sates in 2005. Each colony consisted of 300–400 individuals. Sites were chosen to represent replicate populations near the extremes of the distribution of Ae. albopictus in North America (northern versus southern latitudes). Two southern populations were collected from sites in central Florida where Ae. albopictus has been established since the early 1990s: Memorial Gardens Cemetery, Fort Myers (26.7° N, 81.8° W) (FM); and Rose Hill Cemetery, Tampa (28.0° N, 82.4° W) (TA). One northern population was collected from two separate tire piles ≈2 km apart near Bloomington, IN (39.1° N, 86.6° W and 39.1° N, 86.5° W) (BL). A second northern population was collected from a tire pile in Manassas, VA (38.6° N, 77.4° W) (VA), 40 km southwest from downtown Washington, DC. Ae. albopictus has been collected in Bloomington since the early 1990s and the Washington, DC, metropolitan area since the late 1990s. All populations were >170 km apart. Northern populations experience longer and more severe winters, based on climate data from local meteorological stations (<14.9 km from collection sites) of the National Climate Data Center (http://www.ncdc.noaa.gov/oa/mpp/freedata.html). These data represent the long-term (1971–2000) mean conditions at each site. The lengths of winters, estimated as the number of days between the mean dates of first and last frost, were as follows: FM, 0 d; TA, 0 d; BL, 174 d; and VA, 191 d. Previous studies examining the latitudinal limits of Ae. albopictus in the eastern United States and its native regions in China and Korea have shown that January temperature, as a measure of the severity of winter, is a good predictor of Ae. albopictus distribution (Nawrocki and Hawley 1987, Hanson and Craig 1995). January had the coldest mean daily temperature among all months at all the sites in this study: BL, −2.3°C; VA, −0.2°C; FM, 18.3°C; and TA, 16.3°C. All sites had similar mean annual precipitation (1,062–1,366 mm) but slightly different patterns of monthly rainfall. Southern populations, but not northern populations, experienced 4 mo (January–April) per year with dry conditions (<60 mm precipitation per mo).
Colony adults were kept in cages in an insectary at 26°C at 90% RH under a photoperiod of 16:8 (L:D) h, and they had continuous access to 20% sugar solution. Females took bloodmeals from anesthetized guinea pigs, and they laid eggs on paper towels in water-containing cups. Eggs were hatched synchronously in a solution of 0.44 g of nutrient broth per liter of deionized water. Experimental mosquitoes were the first generation progeny from individuals that were field collected as larvae. They were reared as larvae at low densities (20 larvae per 200 ml) in 250-ml cups provisioned with either 20 or 30 mg of bovine liver power with the goal of providing variable submaximal nutritional levels to produce a wide range of adult sizes. Each day, we collected pupae and put them in individual vials until adult eclosion.
Experimental Setup
As adults eclosed they were placed in 20-liter see-through nylon screen cages that had a sleeve for access. Adults were provided with daily access to 20% sugar solution. Depending on mosquito availability, three to 49 adults, always with more males than females, were released into each cage over 1–5 d. Because males typically eclosed before females and thus were released into cages first, the number of males in a cage dictated the number of females added to that cage.
Ten days after the first females were added in a cage, all females in the cage were given the opportunity to feed to repletion. Females were given 10 min to commence feeding on an anesthetized mouse. Females that did not feed within 10 min were not given the opportunity to feed again, and they were excluded from the experiment. By limiting the time that females were allowed to commence feeding, we potentially selected the most “aggressive” feeders and those females prone to invest more resources into early reproduction. This helps to minimize heterogeneity of vigor and initial condition, ensuring that we are comparing physiologically similar individuals across populations, and potentially giving us a greater chance to detect trade-offs among survivorship, reproduction, and reproductive allocation among populations. We determined that a female had completed blood feeding once she had removed her proboscis from the mouse. Usually, females rested on the mouse for some time after feeding or they flew off the mouse and rested on the side of the screen cage, and they could be easily observed. Although females usually fed to repletion, we nevertheless categorized observed meal sizes into one of three a priori groups: 1) small, one-third repletion or less; 2) medium, ca. one-half repletion; and 3) high, 100% repletion. Mosquitoes were observed (P.T.L.) mating frequently, and it is highly likely that all females were mated. Sample sizes for BL, VA, FM, and TA were 16, 18, 12, and 18 females, respectively (64 total), which are small compared with their relative field populations but similar to other studies of adult survival and reproduction of Ae. albopictus (e.g., Braks et al. 2006).
Upon completion of feeding, each female was collected in a plastic capsule, removed from the 20-liter cage, and isolated into a 600-ml plastic container with a screen top. Within each container, females were provided with an oviposition cup (50-ml black plastic beaker containing ≈25 ml of water and 5 ml of inoculation of 14-d-old oak leaf infusion, and lined with a paper towel) and continuous access to 20% sugar solution. Containers were placed in an environmental chamber at 26°C, 90% humidity, and a photoperiod of 16:8 (L:D) h, with a graduated dawn–dusk period designed to simulate conditions under which Aedes oviposit in the field (Clements 1999). Females were given the opportunity for a bloodmeal every 4 d, by laying an anesthetized mouse on the screen top and extending its tail through the screen and into the cup. We recorded when females took a bloodmeal. All additional bloodmeals seemed to be to repletion, so later meal sizes were not categorized.
Plastic containers were checked daily for eggs and female survivorship. Any eggs were counted and the mean dry mass of each clutch was calculated from a sample of three randomly chosen eggs that were immediately dried for 24 h at 50°C, stored, and later weighed together to 0.1 μg. At death, females were dissected and the numbers of mature eggs (stages 4 and 5, Detinova 1962) were counted. Wings were removed, dried (24 h at 50°C), and measured. Wings of 12 females were sufficiently worn that they could not be measured. The experiment continued until all mosquitoes had died.
Statistical Analyses
For each female, longevity, number of bloodmeals, reproductive output, age of first oviposition, daily reproductive output, blood-feeding frequency, number of days from last oviposition to death, and number of retained eggs at death were all determined. Longevity was measured as the number of days from initial bloodmeal to death. Reproductive output was calculated as the product of a female’s mean egg mass and cumulative number of eggs laid. Our statistical conclusions were the same whether we analyzed eggs oviposited (fecundity) or eggs oviposited + eggs retained at death (investment). The reproductive output for every clutch of eggs was calculated and summed for each individual. Daily reproductive output was calculated as total reproductive output divided by longevity. Blood-feeding frequency was calculated as the total number of meals a female took divided by the number of meals offered. Reproductive output was divided into two parts: early reproduction and residual reproduction. Early reproduction represented egg clutches laid before a second bloodmeal (i.e., the first gonotrophic cycle), whereas residual reproduction represented all later output. If females did not take a second bloodmeal, early reproduction equaled total reproductive output. Females can retain eggs into a future oviposition cycle, but egg retention from the first oviposition cycle into the second cycle is probably minimal (Packer and Corbet 1989); thus, the number of eggs laid before the second bloodmeal is likely to be the full complement of eggs derived from protein obtained from the first bloodmeal.
Estimated age-specific hazard rate was compared among populations by using multivariable Cox proportional hazard model (SAS PROC TPHREG, SAS Institute 2003). To test for a physiological cost of reproduction in our laboratory, cumulative number of bloodmeals and cumulative reproductive output were included in the model as time-dependent covariates (i.e., their values can change over the course of the experiment, SAS Institute 2003). Population, wing length, initial bloodmeal size, and date of initial bloodmeal were included in the model as time-independent variables. Hazard functions (mean hazard rate at age x) were calculated for each population in TPHREG by using the BASELINE statement. For qualitative comparison with the hazard functions, life tables were prepared and data on the expected reproductive output (mx = mean reproductive output for each daily age x), adult reproductive rate (Σlxmx,, where lx represents the probability of being alive at day x), and cumulative number of bloodmeals were calculated and graphed.
Overall associations between longevity, bloodmeal number, and reproductive output, and between early reproduction and residual reproduction, were examined using partial correlations, after controlling for wing length and date by using SAS PROC CORR (SAS Institute 2003). Patterns of life history traits were compared among populations by multiple analyses of covariance (MANCOVAs) and analyses of covariance (ANCOVAs), by using SAS PROC GLM (SAS Institute 2003). In all MANCOVAs, population and initial bloodmeal size were class variables and date of initial bloodmeal and wing length were covariates. The first MANCOVA, on the number of days from last oviposition to death and the number of retained eggs at death, quantified the status of females at death. The second MANCOVA was on longevity, number of bloodmeals, and reproductive output and was used to examine the allocation of resources to maintenance versus reproduction. Follow-up ANOVAs on daily reproductive output and blood-feeding frequency were done to test the effect of population on adult reproductive rates. A third MANCOVA on age at first oviposition, early reproduction, and residual reproduction was used to test for differences in allocation of reproduction over time. A fourth MANCOVA on fecundity and egg mass was used to test for a tradeoff in allocation of resources among offspring. For all MANCOVAs, we used F statistics derived from Pillai’s trace (SAS Institute 2003). We interpreted the contributions of individual dependent variables to significant multivariate effects by using standardized canonical coefficients (Scheiner 2001). To meet assumptions of normality and homogeneity of variances, days from last oviposition to death, number of retained eggs, number of bloodmeals, age at oviposition, daily reproduction rate, and residual reproduction were log-transformed, and blood-feeding frequency was arcsine square-root transformed.
Preliminary tests indicated no significant difference among populations for slopes relative to wing length (results not presented). Interactions with population of both initial bloodmeal size and date could not be tested because not all populations were represented at each bloodmeal size or date of initial bloodmeal. For hazard analyses, preliminary tests also indicated no significant differences among populations for slopes relative to cumulative bloodmeals and cumulative reproductive output. For all analyses, we ran reduced models without interactions involving population to test the main effects of date of initial bloodmeal, wing length, and initial bloodmeal size, and eliminated those main effects when they were nonsignificant. Regardless of whether date of initial bloodmeal, wing length, or initial bloodmeal size, were significant and retained in the reduced model, we report their tests of significance.
Populations in this study were not a randomly selected sample of all possible populations in the north or south. Therefore, population was treated as a fixed effect and statistical inferences extend only to the populations selected. This strategy follows that of previous studies on geographic variation of life history traits (e.g., Reznick et al. 2001). For all analyses, we tested for significant differences among populations by using multivariate pairwise contrasts (Scheiner 2001), with sequential Bonferroni correction for all possible comparisons (six) within each analysis. We tested for a region effect using the a priori contrast comparing average values of northern versus southern sites. Population effects on frequency data were tested using two-tailed Fisher exact tests (SAS Institute 2003, PROC FREQ). Because of the relatively low numbers of mosquitoes in the study and because our emphasis was on detecting biological patterns, we used a more liberal experiment-wise α = 0.10. Main population effects were evaluated at α = 0.05.
Results
Initial Bloodmeal and Oviposition
Of the 64 females that took a bloodmeal and were included in the experiment, 57 fed to repletion, seven took a medium-sized meal, and none took a small meal. One quarter of females from BL took a medium meal (4/16), whereas only 2/18 (11.1%), 1/18 (5.6%), and 0/12 (0%) from VA, TA, and FM, respectively, took medium meals. Most females that took a bloodmeal oviposited (57/64; 89.1%), including six of the seven females that took a medium-sized meal, indicating that one medium-sized bloodmeal was usually sufficient for oogenesis. BL had the greatest proportion of females that did not oviposit, with nearly one third of its total females (5/16; 31.3%) dying without laying eggs. One and two females did not oviposit from VA (1/18; 5.6%) and FM (2/12; 16.7%), respectively, whereas all females from TA laid eggs. Associations of population with bloodmeal size (two-tailed Fisher exact test: expected P = 0.0162, observed P = 0.1904) and oviposition (two-tailed Fisher exact test: expected P = 0.0024, observed P = 0.0237) were both nonsignificant, likely due to low sample sizes.
MANCOVA on the number of days from last oviposition to death and number of retained eggs showed no significant effects of date of initial bloodmeal and initial bloodmeal size (Table 1); thus, these effects were dropped from the reduced model. Wing length was significant, and it strongly affected days from last oviposition to death but not number of retained eggs (see standardized canonical coefficients in Table 1). The parameter estimate (±SE) for the effect of wing length on days from last oviposition to death was −1.18 ± 0.47, indicating that larger females survived for fewer days after their last oviposition than did smaller females. The reduced model showed no significant effect of population (Table 1).
Table 1.
MANCOVA results and standardized canonical coefficients for the number of days from last oviposition until death and the number of retained eggs
| Source of variation | Pillai’s trace (F) | df | P | Canonical variates
|
Standardized canonical coefficients
|
|||
|---|---|---|---|---|---|---|---|---|
| Variate no. | % variation | P | Days from oviposition to death | Retained eggs | ||||
| Pop | 0.21 | 6, 80 | 0.9722 | 1 | 89.4 | 0.9736 | 0.33 | 0.96 |
| 2 | 10.6 | 0.9347 | 1.01 | −0.17 | ||||
| Initial bloodmeal size | 0.90 | 3, 37 | 0.4167 | 1 | 100 | 0.4167 | −0.36 | 0.86 |
| Wing length | 3.14 | 2, 37 | 0.0548 | 1 | 100 | 0.0548 | 1.03 | 0.18 |
| Date of initial bloodmeal | 0.11 | 2, 37 | 0.8964 | 1 | 100 | 0.8964 | 0.89 | −0.40 |
Multivariate pairwise contrasts of populations are not shown as the population effect was nonsignificant. Significant effects are indicated in bold.
Population Differences and Physiological Costs of Reproduction
The effects on age-specific hazard rate of initial bloodmeal size and date of initial bloodmeal were nonsignificant (Table 2); thus, they were dropped from the model. Wing length was significant, with a hazard ratio indicating that hazard rate increased with body size (Table 2). In the reduced model, age-specific hazard rate was significantly affected by the population origin of Ae. albopictus (Table 2). The hazard curve of BL inclines steeply, before all other populations, at ≈30 d (Fig. 1A). However, multiple pairwise contrasts revealed that the hazard rate for BL was only significantly higher than that of VA. All other pairwise contrasts among populations were nonsignificant (sequential Bonferroni correction, Table 2). The hazard curves for all populations curved upward (Fig. 1A), indicating that hazard rate increased with time for Ae. albopictus, regardless of origin.
Table 2.
Results for survival analysis using multivariable Cox proportional hazard model
| Parameter | χ2 | df | P | Hazard ratio |
|---|---|---|---|---|
| Pop | 11.66 | 3, 45 | 0.0087 | |
| Northern vs. southern | 0.22 | 1, 45 | 0.6411 | 0.413 |
| BL vs. FM | 4.09 | 1, 45 | 0.0432 | 0.409 |
| BL vs. TA | 3.89 | 1, 45 | 0.0487 | 0.413 |
| BL vs. VA | 11.60 | 1, 45 | 0.0007 | 0.225 |
| FM vs. TA | 0.00 | 1, 45 | 0.9803 | 1.011 |
| FM vs. VA | 1.98 | 1, 45 | 0.1589 | 0.551 |
| TA vs. VA | 2.01 | 1, 45 | 0.1589 | 0.546 |
| Cumulative no. of bloodmeals | 4.37 | 1, 45 | 0.0377 | 0.421 |
| Cumulative reproductive output (μg) | 4.17 | 1, 45 | 0.0411 | 1.005 |
| Initial bloodmeal size (R vs. M) | 0.08 | 1, 43 | 0.7763 | 0.871 |
| Wing length (mm) | 3.74 | 1, 43 | 0.0532 | 7.619 |
| Date of initial bloodmeal | 0.58 | 1, 43 | 0.8093 | 1.003 |
Multivariate pairwise contrasts of populations were made with significance at experimentwise α = 0.10 (sequential Bonferroni correction, six comparisons). Significant effects and pairwise comparisons are indicated in bold. For parameters corresponding to continuous variables, the hazard ratios are ratio of hazard rates for an increase of 1 unit of the variable. For parameters of categorical variables, the hazard ratio is the ratio of the hazard rates between the given category and the reference category. The reference category is listed first.
Fig. 1.
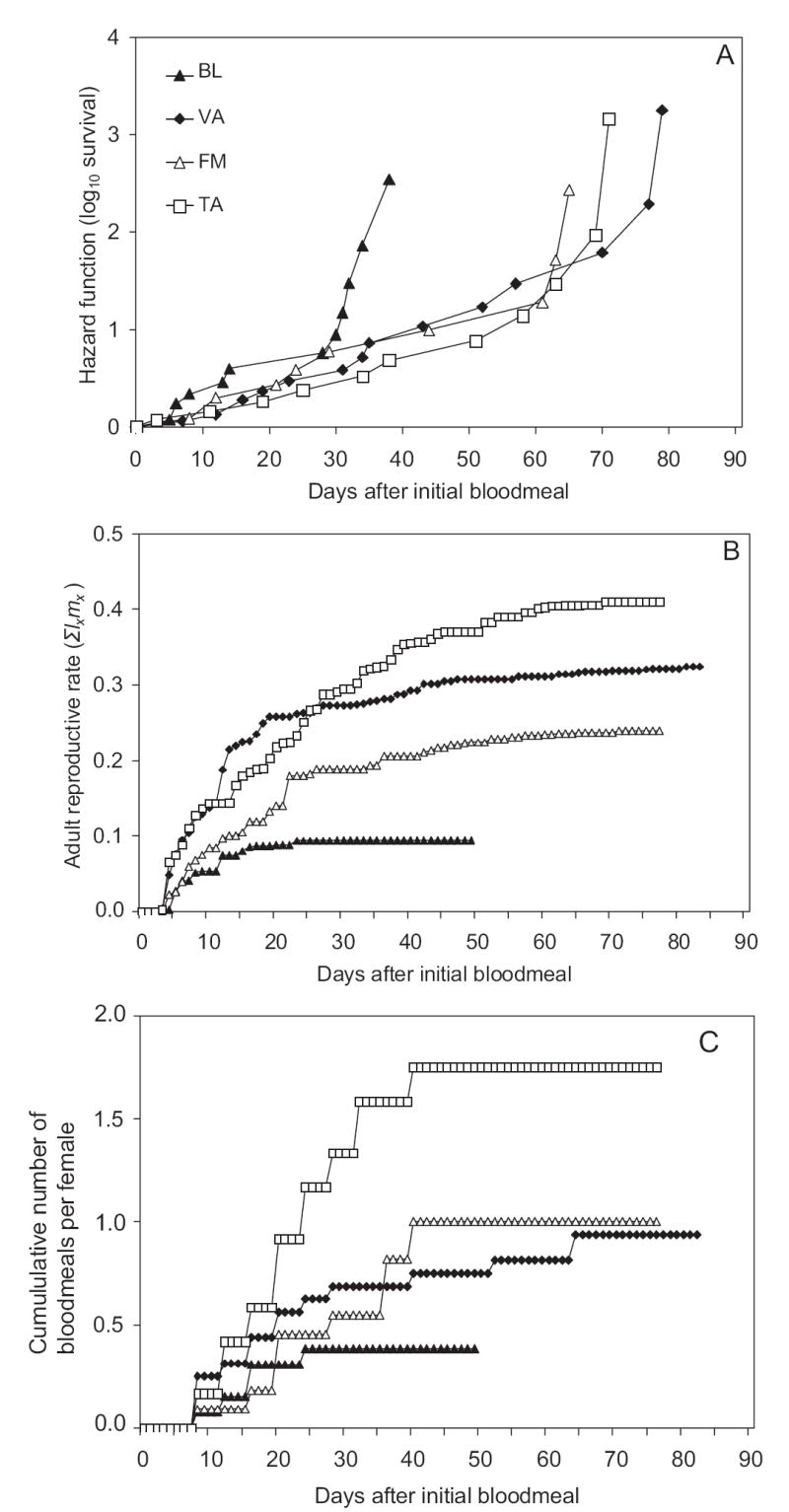
Hazard (log-survivor) function (A), expected reproductive output (mx) (B), cumulative adult reproductive rate (Σlxmx) (C), and cumulative number of bloodmeals from northern (BL and VA) and southern (FM and TA) sites.
Age-specific hazard rate was negatively affected by cumulative number of bloodmeals and positively affected by cumulative reproductive output (Table 2). Overall, hazard rate decreased by >40% with every bloodmeal, and it rose with increasing reproductive output at the average rate of 0.5% per 1 μg of egg mass. The average mass of a single egg was 2.8 μg, indicating that the average overall hazard rate increased by 1.4% for every egg oviposited.
Life Time Allocation of Resources to Maintenance and Reproduction
After controlling for wing length, correlations indicated that bloodmeal number (r = 0.65; P < 0.0001) and reproductive output (r = 0.55; P < 0.0001) were both significantly positively associated with longevity and with each other (r = 0.70; P < 0.0001). MANCOVA on the allocation of resources to maintenance and reproduction across populations showed that the effects of initial bloodmeal size and date of initial bloodmeal were nonsignificant (Table 3); thus, they were dropped from the model. Wing length was significant, and it was retained in the model. Wing length most strongly affected number of bloodmeals and moderately affected longevity, with the parameter estimates (bloodmeals, −0.42 ± 0.20 [mean ± SE:]; longevity −0.41 ± 0.20) indicating that larger females took fewer bloodmeals and did not live as long as smaller females. Wing length only weakly affected reproductive output (Table 3). The reduced model MANCOVA showed a significant effect of population, with population most strongly affecting reproductive output (Table 3; Fig. 2A). Multivariate pairwise contrasts revealed BL was significantly different from both TA and VA. Other pairwise contrasts were nonsignificant (sequential Bonferroni correction) (Table 3). BL differed from TA and VA primarily in reproductive output (Table 3; Fig. 2A and B); females from TA and VA sustained a higher adult reproductive rate throughout their lifetime and especially after 20 d (Fig. 1B). BL females also moderately differed from TA in number of bloodmeals and from VA in number of bloodmeals and longevity (Table 3). Among all populations, females from TA and VA had the greatest reproductive rates and females from TA fed more frequently (Fig. 1B and C). Follow-up ANCOVAs showed a significant population effect on daily fecundity (F3, 60 = 3.33; P = 0.0253), but not on blood-feeding frequency (F3, 60 = 0.80; P = 0.4969). Multiple pairwise contrasts on daily fecundity revealed that the mean for TA was significantly higher than that for BL (F3, 60 = 3.33; P = 0.0253). All other pairwise contrasts were nonsignificant (sequential Bonferroni correction). All other factors showed no significant effects on either daily fecundity or blood-feeding frequency (data not shown).
Table 3.
MANCOVA results and standardized canonical coefficients for the allocation of resources to maintenance and reproduction
| Source of variation | Pillai’s trace (F) | df | P | Canonical variates
|
Standardized canonical coefficients
|
||||
|---|---|---|---|---|---|---|---|---|---|
| Variate no. | % variation | P | No. bloodmeals | Reproductive output | Longevity | ||||
| Pop | 2.66 | 9, 141 | 0.0101 | 1 | 78.7 | 0.0079 | −0.04 | 1.12 | 0.11 |
| 2 | 14.2 | 0.2526 | 1.56 | −0.63 | −0.76 | ||||
| 3 | 7.1 | 0.1789 | 0.21 | −0.80 | 1.12 | ||||
| Northern vs. southern | 1.66 | 3, 45 | 0.1882 | 1 | 100 | 0.1882 | 1.08 | 0.09 | −0.09 |
| BL vs. FM | 1.01 | 3, 45 | 0.3967 | 1 | 100 | 0.3967 | 0.47 | 0.01 | 0.71 |
| BL vs. TA | 5.54 | 3, 45 | 0.0025 | 1 | 100 | 0.0025 | 0.27 | 0.93 | 0.02 |
| BL vs. VA | 3.92 | 3, 45 | 0.0143 | 1 | 100 | 0.0143 | −0.45 | 1.09 | 0.51 |
| FM vs. TA | 2.86 | 3, 45 | 0.0474 | 1 | 100 | 0.0474 | 0.60 | 0.96 | 0.30 |
| FM vs. VA | 1.99 | 3, 45 | 0.1283 | 1 | 100 | 0.1283 | −0.95 | 1.46 | 0.16 |
| TA vs. VA | 1.42 | 3, 45 | 0.2483 | 1 | 100 | 0.2483 | 1.28 | 0.14 | −0.79 |
| Initial bloodmeal size | 0.03 | 3, 43 | 0.9913 | 1 | 100 | 0.9913 | 0.89 | −1.47 | 0.25 |
| Wing length | 2.25 | 3, 43 | 0.0960 | 1 | 100 | 0.0960 | 1.00 | −0.52 | 0.71 |
| Date of initial bloodmeal | 1.48 | 3, 43 | 0.2339 | 1 | 100 | 0.2339 | 1.43 | −0.27 | −0.35 |
Multivariate pairwise contrasts of populations were made at experiment-wise α = 0.10 (sequential Bonferroni correction, six comparisons). Significant effects and pairwise comparisons are indicated in bold.
Fig. 2.

Bivariate plots of the allocation of resources to maintenance and reproduction by population. Data were statistically tested using MANCOVA. Least squares means have been adjusted for the variables used in each analysis. Number of bloodmeals has been backtransformed. Error bars are ±SE.
Reproductive Allocation across Time
MANCOVA on the allocation of reproduction over time showed no significant effects of initial bloodmeal size and wing length, and these effects were dropped from the reduced model (Table 4). Date of initial bloodmeal was significant and retained (Table 4). The reduced model MANCOVA showed a significant effect of population (Table 4). Population most strongly affected residual reproduction, secondarily affected early reproduction, and weakly affected age at first oviposition (Table 4; Fig. 3A and B). Multivariate pairwise contrasts showed that, on average, northern populations were significantly different from southern populations, and that BL differed from both TA and VA. BL females clearly had the lowest cumulative reproductive output across their entire lifetime compared with all other populations (Fig. 1B and C), including VA and thus seemed to drive the significant northern versus southern difference. The clearest distinction between BL and TA was that BL females had lower residual reproduction. The clearest distinction between BL and VA was that BL females had lower early reproduction (Fig. 3A). All other pairwise contrasts were nonsignificant (Table 4). For the first canonical variate for the overall population effect from MANCOVA, and for contrasts of BL versus VA and BL versus TA, the coefficients for both early and residual reproduction were of the same sign, indicating that populations that invested greater resources in early reproduction also invested greater resources in reproduction after the first bloodmeal. A similar positive but weak relationship between early and late reproduction was present within groups (partial correlation, r = 0.085).
Table 4.
MANCOVA results and standardized canonical coefficients for the allocation of reproduction over time
| Source of variation | Pillai’s trace (F) | df | P | Canonical variates
|
Standardized canonical coefficients
|
||||
|---|---|---|---|---|---|---|---|---|---|
| Variate no. | % variation | P | Age at first oviposition | Early reproduction | Residual reproduction | ||||
| Pop | 2.18 | 9, 153 | 0.0265 | 1 | 66.8 | 0.0262 | 0.42 | 0.57 | 0.88 |
| 2 | 28.7 | 0.1597 | 0.54 | 0.51 | −0.58 | ||||
| 3 | 4.5 | 0.3352 | 0.77 | −0.67 | 0.31 | ||||
| Northern vs. southern | 2.35 | 3, 49 | 0.0839 | 1 | 100 | 0.0839 | 0.16 | 0.03 | 1.09 |
| BL vs. FM | 1.09 | 3, 49 | 0.3641 | 1 | 100 | 0.3641 | 0.88 | 0.32 | 0.69 |
| BL vs. TA | 3.65 | 3, 49 | 0.0188 | 1 | 100 | 0.0188 | 0.14 | 0.38 | 1.01 |
| BL vs. VA | 3.78 | 3, 49 | 0.0163 | 1 | 100 | 0.0163 | 0.60 | 0.72 | 0.60 |
| FM vs. TA | 1.36 | 3, 49 | 0.2660 | 1 | 100 | 0.2660 | −0.34 | 0.29 | 0.86 |
| FM vs. VA | 0.72 | 3, 49 | 0.5425 | 1 | 100 | 0.5425 | 0.02 | 0.99 | 0.25 |
| TA vs. VA | 2.02 | 3, 49 | 0.1234 | 1 | 100 | 0.1234 | 0.40 | 0.18 | −0.85 |
| Initial bloodmeal size | 0.10 | 3, 36 | 0.9607 | 1 | 100 | 0.9607 | −0.50 | 0.17 | 0.94 |
| Wing length | 0.99 | 3, 36 | 0.4106 | 1 | 100 | 0.4106 | −0.19 | −0.30 | 1.05 |
| Date of initial bloodmeal | 3.38 | 3, 36 | 0.0287 | 1 | 100 | 0.0287 | −0.57 | 0.17 | −0.88 |
Multivariate pairwise contrasts of populations were made with significance at experimentwise α = 0.10 (sequential Bonferroni correction, six comparisons). Significant effects and pairwise comparisons are indicated in bold.
Fig. 3.

Bivariate plots of the allocation of reproduction over time. Data were statistically tested using MANCOVA. Least squares means have been adjusted for the variables used in each analysis. Residual reproduction and age at reproduction have been backtransformed. Error bars are ±SE.
Reproductive Allocation among Offspring
For the 56 females that oviposited, MANCOVA on allocation of reproduction among offspring showed date, initial bloodmeal size, and wing length were all nonsignificant (Table 5); thus, they were dropped from the model. Egg sizes were similar across populations, but fecundity of BL females tended to be lower than that of all other populations (Fig. 4). Despite this trend, the reduced MANCOVA model showed no significant effect of population (Table 5).
Table 5.
MANCOVA results and standardized canonical coefficients for the allocation of reproduction across offspring
| Source of variation | Pillai’s trace (F) | df | P | Canonical variates
|
Standardized canonical coefficients
|
|||
|---|---|---|---|---|---|---|---|---|
| Variate no. | % variation | P | Egg no. | Egg size | ||||
| Pop | 1.72 | 6, 104 | 0.1364 | 1 | 99.0 | 0.1239 | 1.05 | 0.27 |
| 2 | 1 | 0.9671 | −0.20 | 0.94 | ||||
| Initial bloodmeal size | 0.15 | 2, 37 | 0.8604 | 1 | 100 | 0.8604 | 0.88 | 0.85 |
| Wing length | 0.31 | 2, 37 | 0.7322 | 1 | 100 | 0.7322 | 0.93 | −0.28 |
| Date of initial bloodmeal | 0.89 | 2, 37 | 0.4175 | 1 | 100 | 0.4175 | 1.07 | −0.03 |
Multivariate pairwise contrasts of populations are not shown since population effect was nonsignificant.
Fig. 4.
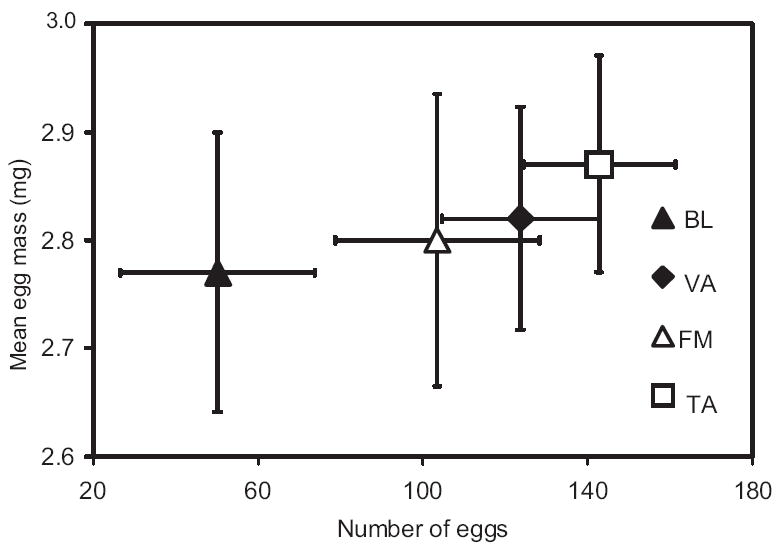
Bivariate plot of the allocation of reproduction among offspring. Data were statistically tested using MANCOVA. Least squares means have been adjusted for the variables used in each analysis. Error bars are ±SE.
Discussion
Our data show innate differences among populations in reproduction and survival of Ae. albopictus. However, all populations showed consistent patterns of resource allocation between reproduction and maintenance, reproduction over time, and reproductive investment among offspring. We found no consistent differences between northern versus southern populations in the mean patterns of life history traits, refuting our predictions based on climate, and they provide evidence that latitudinal variation in climate is probably not driving the evolution of interpopulational differences in reproductive life histories. To our knowledge, this is the first demonstration of geographic variation in adult patterns of survivorship and reproductive tactics of invasive Ae. albopictus in North America.
Regardless of population origin, the age-specific hazard rate of Ae. albopictus increased with increasing reproductive output but decreased with cumulative blood feeding; the latter result being opposite to what we predicted. One hypothesis for this result is that some protein or other nutrients from bloodmeals can be used for maintenance. Briegel and Timmermann (2001) have shown that Ae. albopictus may use only 35–50% of blood protein for oogenesis, and Ae. albopictus, like most anautogenous mosquitoes, divert some protein for maintenance. Therefore, it is possible that protein use by females in this study offset the costs (e.g., energy and time) associated with securing the meal and extended their longevity. An alternative hypothesis is that this relationship is not causal: longer lived females may take more bloodmeals simply because they are offered more meals. However, we found no significant difference among populations in blood-feeding frequency, which does not support this alternative explanation. Another hypothesis is that females that have inherently robust physiology are likely both to live longer and to take more bloodmeals. It is impossible to determine the direction of the causal relationship, if any, between survival and bloodmeal number by using our data, and experiments manipulating timing and number of bloodmeals offered to females are needed.
We found that larger females took fewer bloodmeals, died sooner after last oviposition, and had higher daily hazard rates and lower longevity than did smaller females. These results contrast with the findings from prior studies on several species that have shown body size to be positively correlated with both blood-feeding frequency (Xue et al. 1995) and survival (Packer and Corbet 1989, Lounibos et al. 1990, Briegel and Timmermann 2001), likely because of greater energetic reserves at emergence (Briegel and Timmermann 2001). We found no relationship of wing length with initial bloodmeal size and reproductive output, which also contrasts with most other studies of mosquitoes that have shown body size to be highly positively correlated with both bloodmeal volume (Frankino and Juliano 1999, Briegel and Timmermann 2001) and fecundity (Blackmore and Lord 2000, Armbruster and Hutchinson 2002, Lounibos et al. 2002). We measured initial bloodmeal size relative to body size; thus, the absolute mass of a meal of the same nominal size for a large female exceeded that of a small female. Therefore, although large females took fewer bloodmeals than did small females, they may attain similar overall levels of protein from larger initial (and possibly later) bloodmeals.
To our knowledge, all the studies that have examined the relationship between body size and fecundity in Ae. albopictus have restricted their analysis to the first gonotrophic cycle (Blackmore and Lord 2000, Armbruster and Hutchinson 2002, Lounibos et al. 2002), which is an effective measure of reproductive potential (Clements 1992). Our study suggests that for Ae. albopictus, body size may not be correlated with the lifetime fecundity, or the actual realization of reproductive potential. Most workers make the implicit assumption that there is a direct relationship between the number of eggs in the first gonotrophic cycle and lifetime fecundity. Data on the parous rate of field caught Ae. albopictus also suggest that the average female only matures a single batch of eggs in the wild (Hawley 1988). Although other species have shown a positive relationship between the number of eggs from the first gonotrophic cycle and lifetime fecundity (Shuleman 1990), early fecundity (first gonotrophic cycle) and residual reproduction (all other reproduction) in our study were very weakly correlated (r = 0.085), indicating that initial reproductive investment is a very poor predictor of lifetime fecundity. Although reproduction of laboratory populations cannot be easily extrapolated to field conditions, and there are potential artifacts of laboratory environments (e.g., egg retention due to limited choice of oviposition sites, accidental deaths, restricted movement); nevertheless, the weak relationship between early fecundity and lifetime fecundity of Ae. albopictus raises questions about using results from only the first gonotrophic cycle to estimate reproductive success.
Our study showed nonsignificant interaction effects involving population for both reproductive output and blood feeding, indicating that the physiological stress of oviposition and net effect (positive or negative) of bloodmeals are consistent across populations. BL had the highest hazard rate of all populations (significantly higher than that of VA). If protein from bloodmeals is used for maintenance, this situation may arise because BL females usually took smaller initial meals and rarely took a second meal compared with females from other populations. Compared with BL, TA females had a higher lifetime reproductive output and daily fecundity, which may be the result of a high cumulative number of bloodmeals (see standardized coefficients in Table 3). VA females also had a higher lifetime reproductive output and longevity than BL, which also could be due to a high cumulative number of bloodmeals.
In this experiment, phenotypic measurements on females were made in a common environment so that observed differences in population mean phenotypes probably reflect underlying genetic differences that imply local life history differentiation. Our data suggest that latitudinal variation in climate is probably not driving the evolution of interpopulation differences in life histories. The primary response of North American Ae. albopictus to unfavorable climatic conditions may not involve quantitative variation in the allocation to reproductive output, but rather via the phenotypically plastic alternative life history strategy of producing diapausing eggs rather than developing eggs. Incidence of photoperiodically induced diapause among populations was positively correlated with latitude, from East St. Louis, IL, to the Caribbean (Lounibos et al. 2003). If females from northern populations lay diapausing eggs in response to shorter daylengths well before winter conditions lower adult and larval survival, there may be insufficient selection for greater total and early reproductive investment and larger eggs. We conducted our experiment under summer conditions with a photoperiod of 16:8 (L:D) h. Replicating our experiment under a short daylength (e.g., a photoperiod of 10:14 [L:D] h, which induces diapause in Ae. albopictus), would yield a test of whether latitudinal differences in diapause affect survival and reproductive schedules.
BL females clearly had a different suite of life history traits compared with other populations, having lower survival, feeding frequency, lifetime reproductive output, and reproductive rate. Such differences may arise via nonadaptive forces, such as founder effects or genetic drift. Although our VA site is at approximately the same latitude as the site in BL, the mean daily January temperature at BL (−2.3°C) is lower than that at VA (−0.2°C), and very close to the lowest mean daily January temperatures of overwintering sites for Ae. albopictus in its native areas of China and Korea (−3°C) (Nawrocki and Hawley 1987). Given that BL’s regional January temperature is close to the apparent limit for overwintering for Ae. albopictus, the population from BL may be a sink, maintained by irregular summer colonization from more favorable sites. Thus, the population at BL may be of relatively recent origin and relatively small, and therefore susceptible to founder effects and genetic drift. Unique haplotypes occur in low frequencies in some United States populations indicating that the breeding structure of Ae. albopictus is characterized by drift in local populations (Birungi and Munstermann 2002).
If the BL population has experienced bottlenecks due to low overwinter survival and it has little gene flow from other established populations, another hypothesis for their lower survival and reproduction in this study could be inbreeding depression. In insects, inbreeding can reduce stress resistance (e.g., Armbruster and Reed 2005), increase susceptibility to parasitic infection (e.g., Stevens et al. 1997), negatively affect life history traits such as fecundity (Takaka 1993, Roff 1998), and adult longevity (e.g., van Oosterhout et al. 2000). In wild and laboratory populations of the mosquito Aedes geniculatus (Olivier), estimated net replacement rate was negatively affected by inbreeding across three populations (Armbruster et al. 2000).
This study has shown interpopulational differentiation in the survival and reproductive schedules of North American Ae. albopictus that are not strongly associated with the latitudinal extremes of the species, but that they are associated with an extreme winter climate at one of the sites. Comparing the life history of Ae. albopictus from a larger sample of populations among regions with varying seasonal climates and photoperiods is needed to support this result. Population genetic studies that help describe the phylogeography and patterns of dispersal, bottlenecks, extinction, and recolonization of Ae. albopictus may shed light on this and other interpopulational differences in the life history of this invasive species.
Acknowledgments
We thank Deborah O’Donnell, Peter Armbruster, Michelle Tseng, Courtney Janiec, and Ebony Murrell for field collections or help maintaining the experiment, and Kavitha Damal, Phil Lounibos, and Sabine Loew for useful discussion. This experiment was funded by National Institute of Allergy and Infectious Diseases grant R01-(AI)-44793.
References Cited
- Armbruster P, Hutchinson RA. Pupal mass and wing length as indicators of fecundity in Aedes albopictus and Aedes geniculatus (Diptera: Culicidae) J Med Entomol. 2002;39:699–704. doi: 10.1603/0022-2585-39.4.699. [DOI] [PubMed] [Google Scholar]
- Armbruster P, Reed DH. Inbreeding depression in benign and stressful environments. Heredity. 2005;95:235–242. doi: 10.1038/sj.hdy.6800721. [DOI] [PubMed] [Google Scholar]
- Armbruster P, Conn JE. Geographic variation of larval growth in North American Aedes albopictus (Diptera: Culicidae) Ann Entomol Soc Am. 2006;99:1234–1243. [Google Scholar]
- Armbruster P, Hutchinson RE, Linvell T. Equivalent inbreeding depression under laboratory and field conditions in a tree-hole-breeding mosquito. Proc R Soc Lond B Biol Sci. 2000;267:1939–1945. doi: 10.1098/rspb.2000.1233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bell G. The costs of reproduction and their consequences. Am Nat. 1980;116:45–76. [Google Scholar]
- Birungi J, Munstermann LE. Genetic structure of Aedes albopictus (Diptera: Culicidae) populations based on mitochondrial ND5 sequences: evidence for an independent invasion into Brazil and United States. Ann Entomol Soc Am. 2002;95:125–132. [Google Scholar]
- Black WC, IV, Hawley WA, Rai KS, Craig GB. Breeding structure of a colonizing species: Aedes albopictus (Skuse) in peninsular Malaysia and Borneo. Heredity. 1988;61:439–446. doi: 10.1038/hdy.1988.136. [DOI] [PubMed] [Google Scholar]
- Black WC, IV, Karamjit SR, Turco BJ, Arroyo DC. Laboratory study of competition between United States strains of Aedes albopictus and Aedes aegypti (Diptera: Culicidae) J Med Entomol. 1989;26:260–271. doi: 10.1093/jmedent/26.4.260. [DOI] [PubMed] [Google Scholar]
- Blackmore MS, Lord CC. The relationship between size and fecundity in Aedes albopictus. J Vector Ecol. 2000;35:212–217. [PubMed] [Google Scholar]
- Braks MAH, Juliano SA, Lounibos LP. Superior reproductive success on human blood without sugar is not limited to highly anthrophilic mosquito species. Med Vet Entmol. 2006;20:53–59. doi: 10.1111/j.1365-2915.2006.00612.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Briegel H, Timmermann SE. Aedes albopictus (Diptera: Culicidae): physiological aspects of development and reproduction. J Med Entomol. 2001;38:566–571. doi: 10.1603/0022-2585-38.4.566. [DOI] [PubMed] [Google Scholar]
- Clements AN. Development, nutrition and reproduction. I. Chapman & Hall; London, United Kingdom: 1992. The biology of mosquitoes. [Google Scholar]
- Clements AN. Sensory reception and behaviour. II. CABI Publishing; New York: 1999. The biology of mosquitoes. [Google Scholar]
- Detinova TS. Age-grouping methods in Diptera of medical importance. World Health Organization; Geneva, Switzerland: 1962. [PubMed] [Google Scholar]
- Fox CW, Czesak ME. Evolutionary ecology of progeny size in arthropods. Annu Rev Entmol. 2000;45:341–369. doi: 10.1146/annurev.ento.45.1.341. [DOI] [PubMed] [Google Scholar]
- Frankino WA, Juliano SA. Costs of reproduction and geographic variation in the reproductive tactics of the mosquito Aedes triseriatus. Oecologia (Berl.) 1999;120:59–68. doi: 10.1007/s004420050833. [DOI] [PubMed] [Google Scholar]
- Gerhardt RR, Gottfried KL, Apperson CS, Davis BS, Erwin PC, Smith AB, Panella NA, Powell EE, Nasci RS. The first isolation of La Crosse virus from naturally occurring infected Aedes albopictus. Emerg Infect Dis. 2001;7:807–811. doi: 10.3201/eid0705.017506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanson SM, Craig GB., Jr Aedes albopictus (Diptera: Culicidae) eggs: field survivorship during northern Indiana winters. J Med Entomol. 1995;32:599–604. doi: 10.1093/jmedent/32.5.599. [DOI] [PubMed] [Google Scholar]
- Hawley WA. The biology of Aedes albopictus. J Am Mosq Control Assoc. 1988;4(Suppl):1–40. [PubMed] [Google Scholar]
- Ibañez-Berñal SB, Briseño JP, Mutebi EA, Rodriguez G. First record in America of Aedes albopictus naturally infected with dengue virus during the 1995 outbreak at Reynosa, Mexico. Med Vet Entomol. 1997;11:305–309. doi: 10.1111/j.1365-2915.1997.tb00413.x. [DOI] [PubMed] [Google Scholar]
- Juliano SA, Lounibos LP. Ecology of invasive mosquitoes: effects on resident species and on human health. Ecol Lett. 2005;8:558–574. doi: 10.1111/j.1461-0248.2005.00755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Juliano SA, O’Meara GF, Morrill JR, Cutwa MM. Desiccation and thermal tolerance of eggs and the coexistence of competing mosquitoes. Oecologia (Berl.) 2002;130:458–469. doi: 10.1007/s004420100811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kesavaraju B, Damal K, Juliano SA. Do natural container habitats impede invader dominance? Predator-mediated coexistence of invasive and native container-dwelling mosquitoes. Oecologia (Berl.) 2008 doi: 10.1007/s00442-007-0935-4. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lounibos LP. Invasions by insect vectors of human disease. Annu Rev Entomol. 2002;47:233–266. doi: 10.1146/annurev.ento.47.091201.145206. [DOI] [PubMed] [Google Scholar]
- Lounibos LP, Escher RL. Sex-ratios of mosquitoes from long-term censuses of Florida treeholes. J Am Mosq Control Assoc. 2008 doi: 10.2987/5656.1. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lounibos LP, Larson VL, Morris CD. Parity, fecundity and body size of Mansonia dyari in Florida. J Am Mosq Control Assoc. 1990;6:121–126. [PubMed] [Google Scholar]
- Lounibos LP, Escher RL, Lourenço-de-Oliveira R. Asymmetric evolution of photoperiodic diapause in temperate and tropical invasive populations of Aedes albopictus (Diptera: Culicidae) Ann Entomol Soc Am. 2003;96:512–518. [Google Scholar]
- Lounibos LP, Suárez S, Menéndez Z, Nishimura N, Escher RL, O’Connell SM, Rey JR. Does temperature affect the outcome of larval competition between Aedes aegypti and Aedes albopictus. J Vector Ecol. 2002;27:86–95. [PubMed] [Google Scholar]
- Michod RE. Evolution of life histories in response to age-specific mortality factors. Am Nat. 1979;113:531–550. [Google Scholar]
- Moore CG. Aedes albopictus in the United States: current status and prospects for further spread. J Am Mosq Control Assoc. 1999;15:221–227. [PubMed] [Google Scholar]
- Nawrocki SJ, Hawley WA. Estimation of the northern limits of distribution of Aedes albopictus in North America. J Am Mosq Control Assoc. 1987;3:314–317. [PubMed] [Google Scholar]
- O’Meara GF, Evans LF, Jr, Gettman AD, Cuda JP. Spread of Aedes albopictus and decline of Ae. aegypti (Diptera: Culicidae) in Florida. J Med Entomol. 1995;32:554–562. doi: 10.1093/jmedent/32.4.554. [DOI] [PubMed] [Google Scholar]
- Packer MJ, Corbet PS. Size variation and reproductive success of female Aedes punctor (Diptera: Culicidae) Ecol Entomol. 1989;14:297–309. [Google Scholar]
- Reznick D, Butler MJ, IV, Rodd H. Life history evolution in guppies. VII. The comparative ecology of high-and low-environments. Am Nat. 2001;157:12–26. doi: 10.1086/318627. [DOI] [PubMed] [Google Scholar]
- Rios L, Maruniak JE. Asian Tiger Mosquito, Aedes albopictus (Skuse) (Insecta: Diptera: Culicidae) Publication EENY-319, Featured Creatures series, Entomology and Nematology Department, Florida Cooperative Extension Service, Institute of Food and Agricultural Sciences, University of Florida. 2004 April; http://creatures.ifas.ufl.edu.
- Roff DA. Inbreeding on morphological and life history traits of the sand cricket, Gryllus firmus. Evolution. 1998;55:111–121. [Google Scholar]
- Roff DA. Life history evolution. Sinauer; Sunderland, MA: 2002. [Google Scholar]
- Roitberg BD, Sircom J, Roitberg CA, Vanalphen JJM, Mangel M. Life expectancy and reproduction. Nature (Lond.) 1993;364:108. doi: 10.1038/364108a0. [DOI] [PubMed] [Google Scholar]
- Rowe L, Ludwig D. Size and time of metamorphosis in complex life cycles: time constraints and variation. Ecology. 1991;72:413–427. [Google Scholar]
- SAS Institute. SAS user’s guide: statistics, version 9.1 computer program. SAS Institute; Cary, NC: 2003. [Google Scholar]
- Scheiner SM. MANOVA: multiple response variables and multispecies interactions. In: Scheiner SM, Gurevitch J, editors. Design and analysis of ecological experiments. 2. Oxford University Press; Oxford, United Kingdom: 2001. pp. 99–115. [Google Scholar]
- Shuleman M. Intraspecific variation in the reproductive capacity of Anopheles stephensi (Diptera: Culicidae) J Med Entomol. 1990;27:819–828. doi: 10.1093/jmedent/27.5.819. [DOI] [PubMed] [Google Scholar]
- Sota T, Mogi M. Interspecific variation in desiccation survival time of Aedes (Stegomyia) mosquito eggs is correlated with habitat and egg size. Oecologia (Berl.) 1992;90:353–358. doi: 10.1007/BF00317691. [DOI] [PubMed] [Google Scholar]
- Sprenger D, Wuithiranyagool T. The discovery and distribution of Aedes albopictus in Harris County, Texas. J Am Mosq Control Assoc. 1986;2:217–219. [PubMed] [Google Scholar]
- Stearns SC. The evolution of life histories. Oxford University Press; Oxford, United Kingdom: 1992. [Google Scholar]
- Stevens L, Yan GY, Pray LA. Consequences of inbreeding on invertebrate host susceptibility to parasitic infection. Evolution. 1997;51:2032–2039. doi: 10.1111/j.1558-5646.1997.tb05126.x. [DOI] [PubMed] [Google Scholar]
- Swanson J, Lancaster M, Anderson J, Crandell M, Haramis L, Grimstad P, Kitron U. Overwintering and establishment of Aedes albopictus (Diptera: Culicidae) in an urban La Crosse virus enzootic site in Illinois. J Med Entomol. 2000;37:454–460. doi: 10.1093/jmedent/37.3.454. [DOI] [PubMed] [Google Scholar]
- Takaka Y. A genetic mechanism for the evolution of senescence in Callosobruchus chinensis (the azuki bean weevil) Heredity. 1993;70:318–321. [Google Scholar]
- Turell MJ, Dohm DJ, Sardelis MR, O’Guinn ML, Andreadis TG, Blow JA. An update on the potential of North American mosquitoes (Diptera: Culicidae) to transmit West Nile virus. J Med Entomol. 2005;42:57–62. doi: 10.1093/jmedent/42.1.57. [DOI] [PubMed] [Google Scholar]
- van Oosterhout C, Zijlstra WG, van Heuven MK, Brakefield PM. Inbreeding depression and genetic load in laboratory metapopulations of the butterfly Bicyclus anynana. Evolution. 2000;54:218–225. doi: 10.1111/j.0014-3820.2000.tb00022.x. [DOI] [PubMed] [Google Scholar]
- Xue R, Barnard DR, Schreck CE. Influence of body size and age of Aedes albopictus on human host attack rates and the repellency of deet. J Am Mosq Control Assoc. 1995;11:50–53. [PubMed] [Google Scholar]