

Abstract
The adaptor molecule SAP plays a critical role during natural killer T (NKT) cell development in humans and mice. In CD4+ T cells, SAP interacts with the tyrosine kinase Fyn to deliver signals required for TCR-induced TH2-type cytokine production. To determine whether the SAP-dependent signals controlling NKT cell ontogeny rely on its binding to Fyn, we used the OP9-DL1 system to initiate structure function studies of SAP in murine NKT cell development. In cultures containing wild type (WT) hematopoietic progenitors, we noted the transient emergence of cells that reacted with the NKT cell-specific agonist α-galactosyl ceramide (α-GC) and its analogue PBS57. Sap−/− cells failed to give rise to NKT cells in vitro; however, their development could be rescued by re-expression of WT SAP. Emergence of NKT cells was also restored by a mutant version of SAP (SAP R78A) that cannot bind to Fyn, but with less efficiency than WT SAP. This finding was accentuated in vivo in SapR78A knock-in mice as well as SapR78A competitive bone marrow chimeras, which retained NKT cells but at significantly reduced numbers compared to controls. Unlike SapR78A CD4+ T cells, which produce reduced levels of IL-4 following TCR ligation, α-GC-stimulated NKT cells from the livers and spleens of SapR78A mice produced TH2 cytokines and activated NK cells in a manner mimicking WT cells. Thus, SAP appears to use differential signaling mechanisms in NKT cells, with optimal ontogeny requiring Fyn binding, while functional responses occur independent of this interaction.
Keywords: T cells, cell differentiation, signal transduction
Introduction
Natural killer T (NKT) cells comprise a rare lymphocyte subpopulation with roles in tumor rejection, control of infection and protection from autoimmunity. These cells express markers specific to NK cells as well as markers found on T cells (1). Unlike conventional αβ T cells, however, most NKT cells express an invariant TCR that recognizes glycolipid antigens presented in complex with the MHC class I-like molecule CD1d. Via this invariant TCR, NKT cells bind with high affinity to the model antigen α-galactosyl ceramide (α-GC), which results in their rapid and robust secretion of TH1 and TH2-type cytokines and increased expression of co-stimulatory molecules (1). Through these mechanisms, it is proposed that NKT cells modulate the functions of other immune cell lineages, including T and B lymphocytes and dendritic cells, and thereby link the initial innate and subsequent adaptive arms of the immune response.
Signaling lymphocytic activation molecule (SLAM)-associated protein (SAP) is a cytoplasmic adaptor molecule encoded by the SH2D1A gene that is defective in patients with X-linked lymphoproliferative disease (XLP), a rare immunodeficiency disorder associated with increased susceptibility to EBV infection, development of lymphoma and progressive hypogammaglobulinemia (2). SAP is comprised primarily of a Src homology 2 (SH2) domain, through which it binds to tyrosine residues present within the cytoplasmic domains of several structurally related receptors now known as the SLAM family, including SLAM itself (CD150, encoded by the SLAMF1 gene), Ly9 (CD229; SLAMF3), 2B4 (CD244, SLAMF4), CD84 (SLAMF5) and NTB-A (Ly108 in the mouse; SLAMF6). SAP also binds to the Src family tyrosine kinase Fyn, which it recruits to these receptors (3, 4). In so doing, SAP promotes the generation of ligand-dependent tyrosine phosphorylation signals that support specific cell functions, including CD4+ T cell TH2-type cytokine production (5, 6) as well as NK cell (7–10) and CD8+ T cell cytotoxicity (11, 12).
We and others previously showed that Sap−/− mice and XLP patients lack NKT cells (13–15). Interestingly, Fyn−/− mice also exhibit a reduction in NKT cell number, although not to the same extent as Sap−/− animals (16, 17). The similar phenotypes of Sap−/− and Fyn−/− mice led us to hypothesize that SAP and Fyn must cooperate to control NKT cell development. Here, we use cells expressing wild type (WT) or a mutant version of SAP (R78A) that cannot bind to Fyn to examine the requirement for the SAP-Fyn interaction during NKT cell ontogeny and function. Consistent with our initial hypothesis, we demonstrate that this interaction is critical for efficient NKT cell development in vitro and in vivo. In contrast, it is much less important for NKT cell functions, such as α-GC-induced cytokine production and activation of bystander lymphocytes. These data are the first to examine the signaling properties of SAP in NKT cells and suggest that there is a differential requirement for the SAP-Fyn interaction during ontogeny versus activation.
Materials and Methods
Mouse strains and cell lines
Mice were backcrossed to C57BL/6 for ≥ 7 generations and maintained in specific pathogen-free conditions. All animal procedures were performed according to protocols approved by the Children’s Hospital of Philadelphia. OP9 cell lines were maintained as previously described (18).
Antibodies and flow cytometry
NKT cells were examined using CD24 (M1/69), NK1.1 (PK 136), Thy 1.2 (53-2.1), CD8 (53-6.7), CD4 (RM4.5), TCRβ (H57-597), CD122 (TM-b1), CD69 (FN50), CD44 (IM7), TCRvβ7 (TR-310) and TCRvβ8 (MR5-2) antibodies (BD Pharmingen, San Jose, CA) and CD1d tetramers unloaded or loaded with PBS57, an analogue of α-GC (NIH Tetramer Core Facility, Atlanta, GA). SLAM receptor expression was evaluated using monoclonal antibodies specific for mouse SLAM (TC15-12F12.2), Ly9 (Ly9/ab3) and CD84 (mCD84.7) from Biolegend (San Diego, CA), as well as CD244.2 (2B4; BD Pharmingen) and Ly108 (R & D Systems, Minneapolis MN). Data were acquired using FACSCalibur or FACSVantage flow cytometers and analyzed using CellQuest (BD Biosciences, San Jose, CA) or Flowjo software (Tree Star, Inc., Ashland, OR).
Generation of NKT cells in the OP-9-DL1 system
Cells were isolated from day 13–15 fetal livers (FL). Heat stable antigen (HSA)/CD24low/neg cells were enriched by anti-CD24 antibody depletion and seeded at 0.5–2 × 105 cells in 24-well plates containing Delta-like 1 (DL1)-expressing OP9 cells (OP9-DL1). Cells were maintained in Alpha MEM medium (Invitrogen Gibco, Carlsbad, CA) with 20% fetal bovine serum (FBS), 2 mM glutamine, antibiotics, 2.2 g/L sodium bicarbonate, 5 ng/ml IL-7 and 5 ng/ml Fms-like tyrosine kinase-3 (Flt3) ligand (Peprotech, Rocky Hill, NJ). Every 2 days, cells were fed by adding fresh medium and cytokines. Every 4 days, FL cells were harvested and plated onto a new OP9 monolayer. In indicated experiments, 5 μg/ml of anti-CD1d antibody 3C11 (BD Pharmingen) or an isotype control antibody were added to the cultures.
Detection of α-GC-induced cytokine production by in vitro derived NKT cells
α-GC (KRN7000) and vehicle were provided by Kirin Brewery Company Limited (Tokyo, Japan). FL cells from day 17–18 OP9-DL1 cultures (2 × 105) were placed in ELISPOT plates (R & D systems) in the presence of 2–5 × 105 Sap−/− or Ja281−/− splenocytes with or without 100 ng/ml of α-GC for 6 hours at 37°C. Plates were processed as recommended by the manufacturer, and spots were counted using a stereomicroscope (40 × magnification) or an automated plate reader (Carl Zeiss Microimaging Inc. and K.S. ELISPOT 4.0 software).
Retrovirus infections
Retroviral supernatants were generated as described (19). Sap−/− FL cells were incubated 1 day before infection in Alpha MEM medium with 20% FBS, 2 mM glutamine, antibiotics, 2.2 g/L sodium bicarbonate, 20 ng/ml IL-3, 10 ng/ml IL-6 and 100 ng/ml stem cell factor (Peprotech). Cells were infected with 1 ml of retroviral supernatant and cultured in the same medium (13). On day 2, cells were infected again and placed in culture with OP9-DL1 cells, as described above.
Evaluation of the NKT cell compartment and in vivo NKT cell functions
Quantification of NKT cells, assessment of α-GC-induced cell functions, expression of Sap gene transcripts, detection of Vα14-Jα18 gene transcripts and measurement of CD1d expression and function were performed as previously reported (15, 17, 20).
Measurement of T cell cytokine production
CD4+ T cells were purified from mouse spleens by negative selection using StemSet magnetic columns (StemCell Technologies, Vancouver B.C., Canada). NKT cells were depleted by adding anti-NK1.1 and anti-CD49b (DX-5; BD Pharmingen) antibodies at the time of initial purification. Cell preparations were analyzed by flow cytometry and typically > 95% CD4+ cells and devoid of NKT cells. Cells were stimulated on plates coated with an agonistic anti-CD3 antibody with or without soluble anti-CD28 antibody, in the presence of IL-2 (100 Units/ml). Cells were also stimulated with PMA (100 ng/ml) plus ionomycin (1μM; Sigma Aldrich, Oakville, Ontario, Canada). After 48 hours, supernatants were harvested and cytokine production determined by ELISA (R&D Systems).
Generation of competitive BM chimeras
BM cells were obtained from CD45.1+ C57BL/6.SJL mice (Taconic Laboratories, Germantown, NY), as well as Sap−/− or SapR78A CD45.2+ mice. Cells were washed, mixed at a ratio of 1:1, and 1 × 106 cells were injected intravenously into Rag2−/− recipients (Taconic) that had been treated with 550 rads of γ irradiation (13). Mice were maintained on sterile water containing the antibiotic Bactrim for 2–3 weeks and then switched to sterile water alone. After 6–8 weeks, percent donor chimerism and reconstitution of lymphocyte populations was determined.
Statistical analysis
Statistical analyses were completed using a 2-tailed Student’s t-Test or an analysis of variance, as appropriate (ANOVA, Excel software).
Results
Generation of NKT cells using the OP9 co-culture system
Although SAP is essential for NKT cell ontogeny, the underlying mechanisms remain unknown. Prior studies of mice deficient in the expression of SLAM, SAP or Fyn indicate that each of these molecules is required to enable optimal production of TH2-type cytokines by TCR-stimulated CD4+ T cells (5, 6). Furthermore, the physical interaction of SAP and Fyn is critical, as T cells expressing a mutant version of SAP (R78A) that cannot bind to Fyn exhibit defective IL-4 production and altered Gata-3 upregulation (5, 6). Based on these data and on our initial prediction that SAP and Fyn cooperate during NKT cell ontogeny, we speculated that a SAP-Fyn signaling complex is important for NKT cell development. To address this possibility, we took advantage of an in vitro system of lymphocyte development to model NKT cell ontogeny that would also allow for specific manipulations of Sap gene expression.
Presence of the Notch ligand DL1 on OP9 bone marrow (BM) stromal cells induces T lineage commitment in vitro in a manner similar to T cell differentiation in the thymus (18). Since NKT cells develop in the thymus, we reasoned that they could be generated using this system under appropriate conditions. Therefore, we cultured FL hematopoietic progenitors from WT mice on OP9 cells expressing DL1 (OP9-DL1). Consistent with prior reports, OP9-DL1 cells induced a normal program of early T cell differentiation, as demonstrated by the generation of CD4+CD8+ [double positive (DP)] thymocytes (Fig. 1A, lower plots). Between days 14–18 of culture, we also detected the transient emergence of a subpopulation of cells that reacted with anti-TCRβ antibodies and with CD1d tetramers loaded with the α-GC analogue PBS57 (Tet; Fig. 1A, upper plots) but not unloaded tetramers (Unl; Fig. 1A, middle plots), which suggested the presence of invariant NKT cells. In further support of this possibility, the development of these tetramer-reactive cells required CD1d-mediated intercellular interactions, as they failed to appear in cultures of WT FL cells that contained a blocking anti-CD1d antibody (Fig. 1B, middle plots) or in cultures of CD1d−/− FL cells (Fig. 1B, lower plots). Similar to the development of thymocytes in this system, emergence of tetramer-reactive NKT cells required the presence of a Notch ligand on OP9 stromal cells, since they did not appear when WT FL cells were cultured on native OP9 cells that did not express DL1 (data not shown). In vitro generated NKT cells expressed rearranged Vα14-Jα281 gene transcripts characteristic of the invariant TCRα chain (Fig. 1C). Furthermore, they exhibited a pattern of surface marker expression similar to WT thymic NKT cells, including positivity for TCR Vβ8/7, NK1.1, CD4 and CD122 (Fig. 1D; top histograms – in vitro derived NKT cells, bottom histograms – thymic NKT cells). Together, these data indicate that it is possible to generate what appear to be invariant NKT cells, albeit transiently, using the OP9-DL1 system.
FIGURE 1. NKT cells arise in OP9-DL1 co-cultures.

(A) WT FL cells were cultured on OP9-DL1 cells as described in the methods, stained with PSB57-loaded (Tet) or unloaded (Unl) CD1d tetramers and anti-TCRβ, CD4 or CD8 antibodies and analyzed for the percentage of NKT or conventional T cells by flow cytometry. (B) WT FL cells were placed into OP9-DL1 cultures with or without the CD1d blocking antibody 3C11. Alternatively, cultures were set up using CD1d−/− FL cells. The percentage of NKT cells was examined by flow cytometry. (C) Real-time RT-PCR quantification of Vα14-Jα281 gene transcripts was performed using FL cells from day 17 OP9-DL1 co-cultures shown in (B). Vα14-Jα281 transcript levels were normalized to GAPDH and shown relative to normalized Vα14-Jα281 transcript levels of WT thymocytes. (D) Tet+ TCRβ+ NKT cells from WT thymus (top histograms) and OP9-DL1 co-cultures (bottom histograms) were evaluated for surface marker expression by flow cytometry. (E) WT splenocytes or a combination of cells from day 17 OP9-DL1 co-cultures and Sap−/− splenocytes were stimulated for 6 hours with medium or 100 ng/mL of α-GC, as described in the methods. IL-4 and IFN-γ-producing cells were detected by ELISPOT assay. The data shown represent > 7 (A), 2 (B), 2 (C), 3 (D) or 2 (E) experiments. Abbreviations: n.d. – no cytokines detected.
OP9-DL1-derived NKT cells produce cytokines following short-term exposure to α-GC
To assess whether these in vitro generated NKT cells were functional, we examined their ability to produce cytokines in response to α-GC. We mixed loosely adherent cells from day 17 WT OP9-DL1 co-cultures with splenocytes from Jα281−/− or Sap−/− mice, which lacked NKT cells but retained CD1d+ APCs that could present α-GC to the putative NKT cells. Cells were then cultured in ELISPOT plates in the presence or absence of α-GC, and the number of cytokine-producing cells was quantified after 6 hours. Using this approach, we observed no cytokine-producing cells in the presence of medium alone (n.d. – none detected; Fig. 1E). In contrast, there was a marked increase in IL-4- and IFN-γ-producing cells after short-term exposure to α-GC (Fig. 1E). The increase in number of cytokine-producing cells was similar to what was observed when we added α-GC to wells containing only WT splenocytes, which served as a positive control. We conclude from these studies that in vitro OP9-DL1-derived NKT cells respond to TCR stimulation and produce cytokines in a manner resembling WT splenic NKT cells.
In vitro NKT cell development involves SLAM receptor expression on hematopoietic cells
Murine NKT cell development is controlled in large part by homotypic interactions mediated by SLAM and Ly108, which are co-expressed on CD1d+ DP cells in the thymus (21). To better understand the signals supporting NKT cell development in vitro, we examined the expression of SLAM family receptors on WT mouse thymocytes and OP9 cells. As reported (21), SLAM, Ly108, Ly9 and CD84 were robustly expressed on DP as well as CD4+ or CD8+ single positive (SP) cells. 2B4, on the other hand, was expressed at only very low levels (Fig. 2, thymocytes). In contrast to DP or SP cells, SLAM receptors were not present on most lineage negative CD4−CD8− double negative (DN) cells (Fig. 2, CD4−CD8−, lineage gated). OP9 cells did not express CD1d or the SAP-associated SLAM receptors (Fig. 2, OP9 cells and not shown). Thus, similar to NKT cell development in the thymus, generation of NKT cells on OP9-DL1 cells requires interaction between CD1d+ hematopoietic cells that also express specific SLAM receptors.
FIGURE 2. SLAM family receptors are expressed on thymocytes but not on OP9 stromal cells.

WT mouse thymocytes were stained with isotype control antibodies, anti-CD4 and -CD8 antibodies, and antibodies recognizing specific SLAM receptors, followed by analysis of thymocyte populations by flow cytometry. SLAM receptor expression was further examined on CD4−CD8− cells by staining with anti-B220, -NK1.1, -γδTCR and -Gr-1 antibodies. Non-T cells were excluded from the analysis by electronic gating. SLAM receptor expression on OP9 cells was examined by flow cytometry after gently lifting cells off plates and staining with the indicated antibodies. Filled histograms – cells stained with isotype control antibodies.
Expression of WT SAP or SAP R78A enables NKT cell development in vitro
To determine whether the OP9-DL1 system would be useful for SAP structure function studies in NKT cell development, we next evaluated whether Sap−/− FL cells could give rise to NKT cells in vitro. In agreement with prior reports of XLP patients and Sap−/− mice (14, 15, 20), Sap−/− FL cells did not produce NKT cells when grown on OP9-DL1 cells (Fig. 3A, upper plots). They did, however, generate DP thymocytes at percentages comparable to control cells (Fig. 3A, lower plots). Prior studies have shown that arginine 78 (R78) of SAP mediates its interaction with Fyn, and mutation of this residue to alanine (R78A) severely disrupts the binding of SAP to Fyn (4). Consistent with this property, the SAP R78A mutant protein fails to support optimal TCR-induced IL-4 production by purified CD4+ T cells (5, 6). To establish whether SAP and Fyn must interact to promote NKT cell ontogeny, we set out to test the ability of SAP R78A to support the differentiation of Sap−/− FL cells into NKT cells using the OP9-DL1 system.
FIGURE 3. Sap−/− FL cells fail to produce NKT cells in OP9-DL1 cultures; however, NKT cell development can be restored by expression of WT SAP or the SAP R78A Fyn-binding mutant.

(A) The upper dot plots depict the percentage of Tet+ TCRβ+ NKT cells in day 17 OP9-DL1 cultures or WT and Sap−/− thymi. The lower plots show the percentage of cells staining with CD4 and CD8 antibodies in day 17 OP9-DL1 cultures. (B) Sap−/− FL cells were left uninfected or were infected with retroviruses encoding GFP alone (MIGR), GFP and WT SAP (SAP) or GFP and SAP R78A (R78A). The percent GFP+ cells among gated Thy1.2+ cells is shown (histograms). GFP+ Thy1.2+ cells were analyzed for the percentage of Tet+ TCRβ+ NKT cells (dot plots). The difference between the percentages of Tet+ TCRβ+ NKT cells in WT or SAP R78A cultures, averaged from 3 independent experiments, was not statistically significant (p = 0.07). (C) RT-PCR was used to measure expression of transcripts encoding WT or mutant SAP using cDNA derived from C57BL/6 thymocytes or cells obtained from the OP9-DL1 cultures shown in (B). Samples were also analyzed for actin expression to control for cDNA quality. The data are representative of 3 (A, B) or 2 (C) independent experiments.
Therefore, we transduced Sap−/− FL cells with bicistronic retroviruses encoding GFP alone, GFP plus WT SAP or GFP plus SAP R78A. Infected cells were placed into culture on OP9-DL1 cells and monitored for the emergence of GFP+ NKT{ cells. As expected, the expression of GFP alone did not support NKT cell development (Fig. 3B, MIGR). In contrast, when cells expressed WT SAP, NKT cell development resembled that of unmanipulated WT hematopoietic progenitors. To our surprise, the SAP R78A mutant also supported NKT cell development (Fig. 3B); however, the percentage of NKT cells that emerged was consistently lower but not statistically different from that induced by WT SAP (Fig. 3B and not shown). Since it was not possible to reliably distinguish visually between hematopoietic and non-adherent stromal cells, we did not calculate the absolute number of NKT cells generated in these experiments. We readily identified transcripts encoding Sap and SapR78A by RT-PCR (Fig. 3C). Thus, the restoration of NKT development is likely due to re-expression of WT or mutant SAP protein. To ascertain whether cells transduced with the SapR78A retrovirus exhibited alterations in the kinetics of NKT cell development, wells were examined daily through day 28 of culture. These longitudinal studies did not reveal any difference between WT and SapR78A-transduced cells in terms of the timing of appearance or disappearance of NKT cells in the wells (data not shown). Therefore, it is not likely that an altered time course of development accounts for the modest but reproducible reduction in NKT cells that was observed in SapR78A-transduced wells at day 17 of culture.
Because these results were unanticipated, we next examined whether SAP R78A could promote sustained NKT cell ontogeny in vivo, a process that requires more complex interactions between NKT cell progenitors and supporting hematopoietic and non-hematopoietic cells. We adoptively transferred Sap−/− BM cells similarly transduced to express GFP, GFP and WT SAP or GFP and SAP R78A into lethally irradiated WT mice. After 6–8 weeks, we examined recipient mice for the presence of GFP+ NKT cells in various organs. All mice exhibited between 55–70% GFP+ cells within the examined organs (Fig. 4A, top histograms and not shown), demonstrating that all retroviruses exhibited similar efficiency in transducing hematopoietic cells. Again, we observed that the SAP R78A mutant could support the development of a reduced percentage of NKT cells (Fig. 4A, lower plots, right column) and that the number of NKT cells in SAP R78A reconstituted mice was statistically lower than the number in mice reconstituted with WT SAP (Fig. 4B). Collectively, these studies suggest that the SAP Fyn-binding mutant retains an ability to promote NKT cell ontogeny, although with reduced efficiency compared to WT SAP.
FIGURE 4. Transduction of Sap−/− BM cells with retroviruses encoding WT SAP or SAP R78A allows NKT cell development in vivo.

SAP-deficient BM cells were transduced in vitro with retroviruses encoding GFP (MIGR), GFP and WT SAP (SAP) or GFP and SAP R78A (SAP R78A) and transferred into lethally irradiated recipients. After 6–8 weeks, reconstituted mice were examined for presence of GFP+ lymphocytes in the thymus (A, top histograms) and percentage (A) or absolute number (B) of GFP+ Tet+ TCRβ+ NKT cells in various organs. The data in (B) are averaged from 1 experiment in which 12 mice were reconstituted with Sap−/− BM cells infected with the indicated retroviruses (MIGR: n = 4; SAP: n = 4; SAP R78A: n = 4). This experiment was repeated 3 times. Ten mice reconstituted with MIGR, 12 mice reconstituted with WT SAP and 9 mice reconstituted with SAP R78A were analyzed.
SapR78A mice have reduced NKT cell numbers
To ensure that the rescue of NKT cell ontogeny in these experiments was due to the expression of WT or mutant SAP protein and not secondary to effects resulting from the use of retroviruses, we chose to examine the NKT cell compartment in SapR78A knock-in mice, which express SAP R78A under control of the endogenous Sap gene promoter and at levels comparable to WT SAP (6). As previously reported (6), SapR78A mice had a normal number of thymocytes and splenocytes (not shown). Furthermore, there was no obvious perturbation in the percentages of mature T cell subsets or in thymic T cell development (data not shown). We next quantified the NKT cells in WT, Sap−/− and SapR78A mice. Consistent with our prior retroviral reconstitution studies, SapR78A animals retained NKT cells, but at a lower percentage than control animals (Fig. 5A and not shown). The absolute number of thymic and splenic NKT cells was also significantly lower in SapR78A mice and measured on average 5- and 4-fold less than the number in WT mice (Fig. 5B). While the average number of liver NKT cells was reduced by 50% in SapR78A mice, this number did not differ statistically from that observed in WT animals. Thymocytes and splenocytes from WT and SapR78A mice expressed CD1d comparably, and following incubation with α-GC, splenocytes of both genotypes induced IL-2 production by invariant TCR+ NKT cell hybridomas (data not shown). Thus, the reduced NKT cell number in SapR78A mice is not likely due to impairments in their expression or function of CD1d.
FIGURE 5. SapR78A mice have reduced NKT cell numbers but normal NKT cell phenotype.

(A) Flow cytometric plots showing the percentage of mature HSAlow Tet+ TCRβ+ NKT cells in a representative cohort of WT, Sap−/− and SapR78A mice. (B) Number of Tet+ TCRβ+ NKT cells from WT (n = 9), Sap−/− (n = 9) and SapR78A (n = 5) mice. Statistical differences in the number of NKT cells in the organs of WT versus SapR78A mice are shown. (C) Thymic Tet+ TCRβ+ NKT cells were examined for expression of HSA (CD24) and NK1.1. The percentage of cells positive for each marker is shown. After gating on TCR+ Tet+ cells within the live lymphocyte gate, cells were also analyzed for expression of NK1.1, CD44, CD24, CD4, CD8, CD122 and CD69. The data in (C) are representative of 4 age-matched mice (6–8 weeks) of each genotype.
In the mouse, NKT cells diverge from conventional αβ T cells at the DP stage of thymocyte development (22, 23), when progenitors expressing the Vα14-Jα281 TCR are selected by interaction with other CD1d-expressing DP cells. Positive selection is followed by cell division and lineage commitment, marked by the loss of HSA (CD24) expression and acquisition of markers typical of NKT cells, including CD44, DX5 and NK1.1 (24). To elucidate why NKT cells were diminished in SapR78A mice, we analyzed the phenotype of PBS57-loaded CD1d tetramer-reactive (Tet+) NKT cells in the thymi of adult 8–12 week old WT, Sap−/− and SapR78A animals. Remarkably, TCR+ tetramer reactive (Tet+) thymocytes from WT and SapR78A mice exhibited a similar profile of surface marker expression, as exemplified by a predominance of mature HSAlow/neg NK1.1+ cells (Fig. 5C). This phenotype was distinct from that seen on the few TCR+ Tet+ cells that remained in Sap−/− thymi, which were arrested at the immature HSA+ NK1.1− stage. In addition to HSA and NK1.1, we could detect no differences in the expression of other developmental markers between unmanipulated WT and SapR78A TCR+ Tet+ thymic NKT cells, including CD44, CD4, CD8, DX5, CD69 and CD122 (Fig. 5C and data not shown).
SapR78A hematopoietic cells develop into NKT cells in competitive BM chimeras
Although SapR78A NKT cell progenitors might exhibit defects in marker expression that went undetected in our analyses, their normal surface phenotype suggests that the diminished number of mature NKT cells in these animals is not the result of gross perturbations in the ability to differentiate into mature NKT cells. Instead, this reduction in cell number could reflect defects in progenitor cell positive or negative selection, expansion or survival. To address the effectiveness of NKT cell selection by SapR78A cells, we performed a series of competitive BM chimera experiments in which CD45-allelically marked WT, Sap−/− or SapR78A BM cells were mixed at a ratio of 1:1 in varying combinations and injected intravenously into sublethally irradiated Rag2−/−hosts. Since WT, Sap−/− and SapR78A DP cells expressed comparable levels of CD1d and SLAM receptors (not shown), we anticipated that within individual chimeras, the relative efficiency of NKT cell selection and/or development could be determined based on the contribution of cells of different genotypes to the thymic and peripheral NKT cell compartments.
WT, Sap−/− and SapR78A BM cells reconstituted chimeric animals equally well, with each genotype comprising 30–70% of the lymphocytes within the thymus, spleen and liver (Fig. 6A and not shown). Each genotype also generated comparable numbers of conventional T, B and NK cells (not shown). As anticipated, Sap−/− BM cells failed to give rise to NKT cells in irradiated recipients, representing on average 0.005% (range 0.0018 – 0.014), 0.018% (0.0024 –0.039) and 0.032% (0.013 – 0.05) of thymic, splenic and intrahepatic lymphocytes (Fig. 6A). These data translate into a 93–97% reduction in the absolute number of NKT cells compared to competing WT counterparts (Fig. 6B). SapR78A BM cells demonstrated improved efficiency over Sap−/− cells in generating NKT cells, and averaged 0.03% (range 0.012–0.046), 0.12% (0.02–0.25) and 1.3% (0.3–2.13) of thymocytes, splenocytes and liver lymphocytes. Although the number of NKT cells generated by SapR78A BM cells was reduced compared to WT competitors, SapR78A BM cells were 9-, 6- and 24-fold better than Sap−/− cells in repopulating the thymic, splenic and hepatic NKT compartments. These data are consistent with our retroviral reconstitution experiments (Fig. 4), and analyses of SapR78A mice (Fig. 5) and suggest that an intact SAP-Fyn signaling axis is important, but not exclusively required, for NKT cell selection and development in the setting of WT hematopoietic competitors.
FIGURE 6. SapR78A hematopoietic cells retain the ability to repopulate the NKT cell compartment in competitive BM chimeras.

(A) WT, Sap−/−and SapR78A BM cells were mixed at a ratio of 1:1 and injected into sublethally-irradiated Rag2−/− mice. Recipients were analyzed at 6–8 weeks of age. (A) The percent chimerism in the thymus and incidence of NKT cells in various organs was analyzed by flow cytometry. To examine the percentage of NKT cells, lymphocytes were gated first based on CD45.1 positivity, followed by the identification of Tet+TCRβ+ cells. (B) Absolute NKT cell numbers were calculated for each genotype as described (21). Data in (B) were averaged from 4 chimeric mice representing each combination of genotypes (WT:WT, WT: Sap−/−, WT:SapR78A), with a total of 12 mice analyzed.
SapR78A NKT cells produce cytokines in response to α-GC
Because SapR78A mice retained a greater number of NKT cells than could be generated using the OP9-DL1 system or the in vivo BM reconstitution approach, we took advantage of these animals to examine whether the Sap R78A mutation could support TCR-induced NKT cell functions. Before beginning these studies, we examined the surface phenotype of splenic and hepatic TCR+ Tet+ cells from WT and SapR78A mice and found that they expressed similar levels of the TCR (data not shown) as well as CD44, CD4, CD8, NK1.1, CD69 and CD122 (Fig. 7A), suggesting that they could be directly compared in functional assays.
FIGURE 7. SapR78A peripheral NKT cells have a normal surface phenotype and produce cytokines in vivo in response to α-GC.

(A) The surface phenotype of Tet+ TCRβ+ NKT cells in the livers and spleens of WT (filled histograms) and SapR78A (open histograms) mice was examined by surface staining and flow cytometry. (B) WT, Sap−/− or SapR78A mice were injected intravenously with 4 μg of vehicle or α-GC (black bars). After 2 hours, serum was collected and cytokines measured by ELISA. Statistical differences between the levels of serum cytokines in α-GC injected WT and SapR78A mice are as indicated. N.d. – no cytokines detected. (C) Mice were similarly injected with vehicle (filled histograms) or α-GC (open histograms) and 2 hours later, the percentages and MFI (in parentheses) of liver Tet+ TCRβ+ NKT cells producing IFN-γ or IL-4 directly ex vivo were analyzed by intracellular cytokine staining and flow cytometric analysis. (D) NKT cell depleted CD4+ T cells were purified from the spleens of WT, Sap−/− or SapR78A mice by negative selection. Cells were placed in culture with IL-2 (100 Units/ml) and stimulated in the presence of medium, plate-bound anti-CD3 antibody (3 μg/ml), anti-CD3 plus soluble anti-CD28 (1 μg/ml), or PMA (100 ng/ml) and ionomycin (1μM). After 48 hours, supernatants were harvested and cytokine production measured by ELISA.
Injection of α-GC into WT mice leads to NKT cell activation, resulting in the rapid production of TH1- and TH2-type cytokines. To investigate whether in vivo cytokine responses were intact in SapR78A mice, we injected WT, Sap−/− and SapR78A animals with vehicle or α-GC. After 2 hours, we harvested sera from injected animals and assayed for the presence of cytokines by ELISA. There were no cytokines detected in the serum of vehicle treated mice (n.d. – no cytokines detected; Fig. 7B). As anticipated, treatment with α-GC resulted in a marked increase in serum IFN-γ and IL-4 in WT mice, while no cytokines were detected in Sap−/− animals. SapR78A mice exhibited an intermediate phenotype, with a ~1.5–3-fold reduction in both cytokines compared to controls. To establish whether the diminished serum cytokine response in SapR78A mice might result from their reduced number of NKT cells, altered NKT cell function or both possibilities, we injected mice with α-GC and 2 hours later examined NKT cell cytokine production by intracellular staining. In these studies, SapR78A NKT cells produced IFN-γ as well as WT cells (Fig. 7C), suggesting that the reduced serum IFN-γ levels in SapR78A mice resulted from their lower NKT cell number. While ~25–30% fewer SapR78A NKT cells produced IL-4 in response to α-GC, those cells that stained for this cytokine did so with a mean fluorescence intensity (MFI) comparable to WT cells (Fig. 7C). These data stand in contrast to TCR-induced IL-4 secretion by purified NKT cell depleted SapR78A CD4+ T cells, which exhibit a 60–70% reduction in the production of this cytokine (references 5, 6 and Fig. 7D). They also differ from prior studies of NKT cell depleted naïve (CD44lowCD62Lhi) and memory (CD44hiCD62Llow) T cells, which demonstrate that the TH2 defect is common to SapR78A CD4+ cells, regardless of their state of activation (6). Although the assays we used to induce and measure cytokine production varied between NKT cells and CD4+ T cells, our findings suggest that NKT cells and conventional T cells differ in their requirement for the SAP-Fyn interaction during TH2-type responses. Since the majority of SapR78A NKT cells robustly produced IL-4 in response to α-GC, we also believe that the diminished serum IL-4 levels observed in these animals primarily reflect their reduced number of NKT cells. However, we cannot exclude a minor functional defect.
We next examined whether SapR78A NKT cells could appropriately activate other immune cells in vivo by injecting animals with α-GC and 4 hours later examining splenocytes for upregulated expression of the CD69 activation marker and production of IFN-γ. In WT mice, α-GC induced robust upregulation of CD69 on > 80% of NK, T and B cells (Fig. 8A) and stimulated the production of IFN-γ by ~30% of splenic NK cells (Fig. 8B). As previously reported, these responses were completely lacking in Sap−/− mice. SapR78A animals exhibited a mixed phenotype, with fewer splenocytes expressing CD69, yet a normal percentage of NK cells secreting IFN-γ. Since the molecular basis underlying α-GC-induced CD69 upregulation is not well understood, we do not know whether this diminished response reflects the reduced NKT cell number in SapR78A mice and/or additional defects in cell function. Nonetheless, the partial preservation of this response, along with the normal NK cell and vigorous NKT cell cytokine responses, suggest that NKT cell functions are for the most part retained in SapR78A mice.
Figure 8. SapR78A NKT cells activate bystander lymphocytes in α-GC-treated mice.
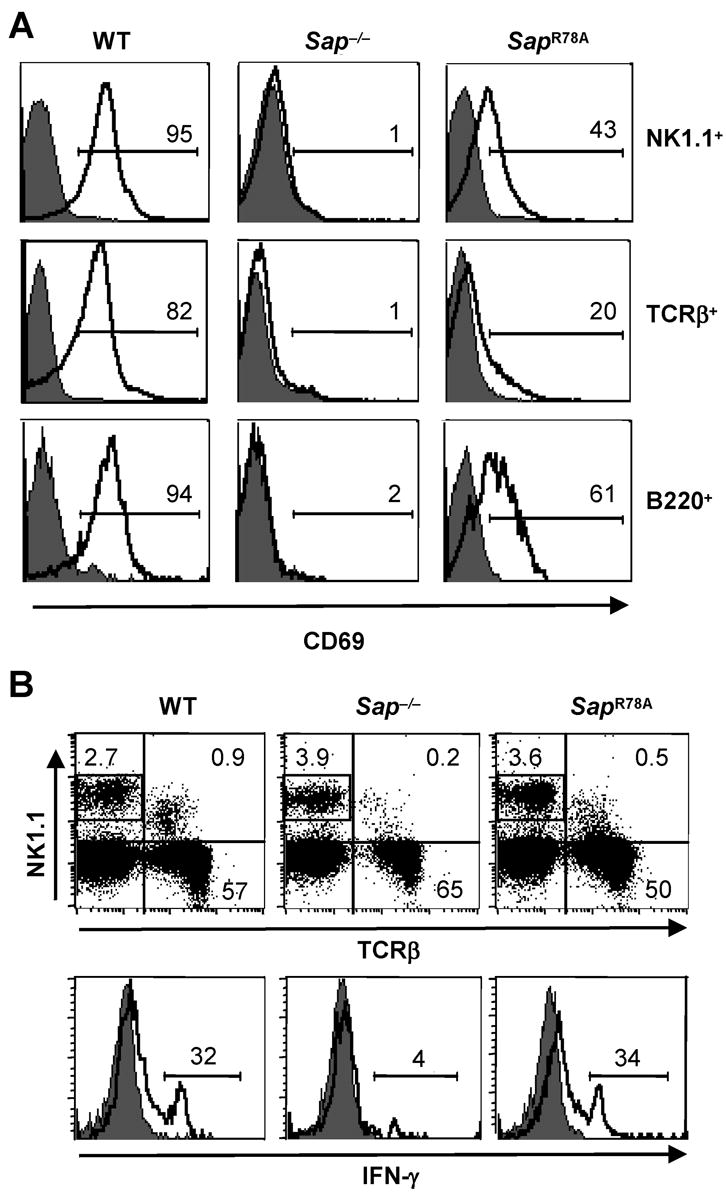
(A) WT, Sap−/− and SapR78A mice were injected with vehicle or α-GC and 4 hours later, splenocytes from vehicle (filled histograms) or α-GC-treated mice (open histograms) were analyzed for the percentage of NK1.1+, TCRβ+ or B220+ cells expressing CD69. Alternatively, splenocytes were stained with NK1.1 and TCRβ antibodies and the percentage of NK1.1+ TCRβ− cells (as gated in the upper dot plots) producing IFN-γ directly ex vivo was determined using intracellular staining and flow cytometry (D). Data are representative of 3 experiments in which a minimum of 4 mice of each genotype were analyzed per condition.
Discussion
NKT cells comprise a unique subset of thymus-derived lymphocytes with developmentally and functionally distinct properties compared to conventional αβ T cells. These cells display a variety of activities, including augmentation of antimicrobial immunity and tumor rejection, suppression of autoimmunity and promotion of tolerance. Consistent with these properties, alterations in NKT cell number or activity are deleterious, as is evident in patients with certain diseases such as cancer, asthma, graft rejection and autoimmune conditions. Based on their role in normal and pathologic immune responses, there has been an increased interest in elucidating the molecular pathways controlling the ontogeny and function of this lineage.
We and other groups showed that patients with XLP, who harbor mutations in the gene encoding SAP, lack peripheral blood NKT cells. These findings are recapitulated in Sap−/− mice, which lack NKT cells in the thymus and other organs. Together, these data indicate that SAP is central to the development of NKT cells. To understand the molecular mechanisms by which SAP orchestrates NKT cell ontogeny and to provide further insights into its signaling capacity in mature NKT cells, we completed the current series of in vitro and in vivo structure function studies. We focused our attention on the importance of the SAP-Fyn interaction, as prior studies have shown that this physical interaction is critical for conventional CD4+ T cell function.
Our data show for the first time that it is possible to generate NKT cells using the OP9-DL1 co-culture system. Like NKT cell ontogeny in the thymus, NKT cell development on OP9 cells requires stromal cell expression of a Notch ligand as well as hematopoietic cell expression of CD1d and specific SLAM receptors. Since the requirements for NKT cell development are similar in vitro and in vivo, it is likely that the OP9-DL1 system will provide an attractive and flexible assay for dissecting the molecules required for NKT cell development in the thymus.
Interestingly, NKT cells appear only transiently in this system. Although we do not understand the mechanisms underlying this finding, there are several possible explanations. First, the cytokine milieu may not be adequate to support the longer-term generation of NKT cells in these cultures. To examine this possibility, we increased the levels of IL-7 and Flt3 ligand in the culture wells and added other factors known to support NKT survival and proliferation, such as IL-15 (25), or differentiation, such as granulocyte macrophage colony stimulating factor (26). However, none of these maneuvers altered the percentage or persistence of NKT cells in the cultures (data not shown). Second, thymic stromal cells express other Notch ligands, such as DL4 (27), which may also be important for NKT cell ontogeny. Since the OP9 cells used in these experiments lacked DL4 expression, it is possible that the lack of this signal did not allow for the extended growth of NKT cells in the tissue culture dish. Third, in addition to T and NKT cells, the thymus contains a limited number of dendritic cells and macrophages, which are not present in OP9-DL1 cultures. Because these cells are involved in cross-presentation of IL-15, they may contribute to the proliferation or survival of NKT cells. Despite these potential limitations, our data suggest that the OP9-DL1 system will provide a powerful tool to identify the molecules that regulate NKT cell development. We are now using this system to identify other signaling molecules that cooperate with SAP during NKT cell ontogeny.
Through use of the OP9-DL1 co-culture system, we find that mutation of the Fyn-binding region of SAP negatively influences the development of NKT cells. More striking impairments in NKT cell development are observed in vivo following reconstitution of lethally irradiated mice with Sap−/− BM cells transduced to express SAP R78A, in knock-in mice that harbor the SapR78A mutation and in mixed BM chimera experiments, where the absolute number of thymic and splenic SAP R78A expressing NKT cells is significantly reduced compared to WT NKT cells. Together, these data are in accord with our original hypothesis and suggest that optimal NKT cell ontogeny requires signals that rely upon a strong SAP-Fyn interaction.
It is intriguing, however, that the defect in NKT cell development induced by the SapR78A mutation is not as severe as that observed in mice lacking expression of SAP itself. There are several potential reasons for this finding. First, it is possible that the Sap R78A mutation allows for a low level of Fyn binding that is sufficient to support the development of a reduced number of NKT cells. This possibility seems unlikely since prior studies of this mutation demonstrate that it is unable to recruit and activate Fyn and to promote signaling in response to SLAM engagement in a T cell line (4). Alternatively, SAP may signal in NKT cell progenitors, albeit less efficiently, in a manner that does not require Fyn binding. In support of this notion, sequences distinct from R78 mediate SAP’s interaction with Lck in vitro and in yeast (28).
Despite their reduced number of NKT cells, the surface phenotype of TCR+ Tet+ thymocytes is grossly normal in adult SapR78A mice. Currently, it is postulated that there are 2 major checkpoints controlling NKT cell ontogeny (24). The first checkpoint occurs at the DP stage of thymocyte development, where immature NKT cell progenitors undergo CD1d-dependent positive selection and diverge from the developmental pathway of conventional αβ T cells. Subsequently, a second checkpoint occurs, where selected progenitors acquire phenotypic and functional characteristics of mature NKT cells. The reduced NKT cell number but normal surface phenotype of SapR78A NKT cells, suggests that abrogation of the SAP-Fyn interaction may preferentially interfere with progression of progenitors beyond checkpoint 1, perhaps by influencing the efficiency of selection, or by diminishing proliferation or survival. For those cells that advance beyond checkpoint 1, differentiation may occur normally, at least as determined based on expression of surface markers typical of mature NKT cells and acquisition of NKT cell functions, such as TCR-induced cytokine production. Through the analysis of tetramer-enriched cells, it was recently shown that NKT cell development is arrested in 2-week-old Sap−/− and Fyn−/− mice at the CD24hiTethiCD69hi stage, which correlates with a period of cellular expansion immediately following positive selection (21). In this study, the arrested cells did not exhibit increased apoptosis, and NKT cell number could not be rescued by expression of the pro-survival molecule Bcl-xL. Along with our data, these findings suggest that manipulations that interfere with the SAP-Fyn interaction, either by reducing the levels of expression of these molecules or by diminishing their ability to bind to one another, lead to defects in NKT cell development that extend beyond the exclusive provision of a survival signal.
Through a series of in vivo functional studies, we demonstrate the surprising finding that in response to α-GC, NKT cells expressing SAP R78A robustly produce IL-4 and IFN-γ and activate NK cells as well as WT cells. Given its inability to fully support TH2-type cytokine production by TCR-stimulated CD4+ T cells (5, 6), it is interesting that the SAP R78A variant enables TCR-induced NKT cell functions. These observations put forth the possibility that the SAP-dependent signals supporting NKT cell IL-4 production are distinct from those required by conventional αβT cells, which are Fyn dependent. However, one must take into consideration that the NKT cells that emerge in SapR78A mice might not reflect the functions of normal NKT cells due to their altered selection in the thymus based on autonomy from Fyn.
Throughout our studies, the SAP R78A mutation appears to have a greater influence on NKT cell number than NKT cell function. This finding is unusual since the requirements for specific signaling molecules are often shared during ontogeny and activation. However, our data closely resemble prior observations of Fyn−/− mice, which exhibit a 5–10-fold reduction in NKT cell number (16, 17) but only modest defects in cell function (29). Collectively, these findings support the idea that the SAP-Fyn interaction is differentially required during NKT cell development versus activation, with development being in large part Fyn-dependent and functions such as TCR-induced cytokine production remaining Fyn-independent. Indeed, Fyn-dependent and -independent functions of SAP have been previously demonstrated (30, 31), with proper CD4 T cell TH2-differentiation requiring Fyn, yet regulated expression of co-stimulatory molecules and T cell help for immunoglobulin expression remaining Fyn independent. Future efforts to identify the signaling molecules that cooperate with SAP in NKT cells will increase our understanding of the ontogeny and function of this unique immune cell lineage and may provide further insights into the pathogenesis and treatment of XLP.
Acknowledgments
We thank Drs. Mitchell Kronenberg, Pamela Schwartzberg, Jennifer Cannons, Luc van Kaer and Juan Carlos Zuniga-Pflucker for providing reagents and expert advise, Taku Kambayashi, Martha Jordan, Jonathan Maltzman and Avinash Bhandoola for reviewing this manuscript and Justina Stadanlick for editorial assistance.
Footnotes
This work was funded by a grant from the NIH (K.E.N).
Abbreviations: NKT, natural killer T cell; α–GC, α-galactosyl ceramide; SAP, signaling lymphocytic activation molecule (SLAM)-associated protein; XLP, X-linked lymphoproliferative disease; FL, fetal liver; DL1, Delta-like 1; WT, wild type; DP, double positive; HSA, heat stable antigen; Tet, tetramer; BM, bone marrow; MFI, mean fluorescence intensity
Conflict of interest statement
The authors declare no competing financial interests.
References
- 1.Taniguchi M, Harada M, Kojo S, Nakayama T, Wakao H. The regulatory role of Valpha14 NKT cells in innate and acquired immune response. Annu Rev Immunol. 2003;21:483–513. doi: 10.1146/annurev.immunol.21.120601.141057. [DOI] [PubMed] [Google Scholar]
- 2.Ma CS, Nichols KE, Tangye SG. Regulation of Cellular and Humoral Immune Responses by the SLAM and SAP Families of Molecules. Annu Rev Immunol. 2007;25:337–379. doi: 10.1146/annurev.immunol.25.022106.141651. [DOI] [PubMed] [Google Scholar]
- 3.Chan B, Lanyi A, Song HK, Griesbach J, Simarro-Grande M, Poy F, Howie D, Sumegi J, Terhorst C, Eck MJ. SAP couples Fyn to SLAM immune receptors. Nat Cell Biol. 2003;5:155–160. doi: 10.1038/ncb920. [DOI] [PubMed] [Google Scholar]
- 4.Latour S, Roncagalli R, Chen R, Bakinowski M, Shi X, Schwartzberg PL, Davidson D, Veillette A. Binding of SAP SH2 domain to FynT SH3 domain reveals a novel mechanism of receptor signalling in immune regulation. Nat Cell Biol. 2003;5:149–154. doi: 10.1038/ncb919. [DOI] [PubMed] [Google Scholar]
- 5.Cannons JL, Yu LJ, Hill B, Mijares LA, Dombroski D, Nichols KE, Antonellis A, Koretzky GA, Gardner K, Schwartzberg PL. SAP regulates T(H)2 differentiation and PKC-theta-mediated activation of NF-kappaB1. Immunity. 2004;21:693–706. doi: 10.1016/j.immuni.2004.09.012. [DOI] [PubMed] [Google Scholar]
- 6.Davidson D, Shi X, Zhang S, Wang H, Nemer M, Ono N, Ohno S, Yanagi Y, Veillette A. Genetic evidence linking SAP, the X-linked lymphoproliferative gene product, to Src-related kinase FynT in T(H)2 cytokine regulation. Immunity. 2004;21:707–717. doi: 10.1016/j.immuni.2004.10.005. [DOI] [PubMed] [Google Scholar]
- 7.Benoit L, Wang X, Pabst HF, Dutz J, Tan R. Defective NK cell activation in X-linked lymphoproliferative disease. J Immunol. 2000;165:3549–3553. doi: 10.4049/jimmunol.165.7.3549. [DOI] [PubMed] [Google Scholar]
- 8.Bloch-Queyrat C, Fondaneche MC, Chen R, Yin L, Relouzat F, Veillette A, Fischer A, Latour S. Regulation of natural cytotoxicity by the adaptor SAP and the Src-related kinase Fyn. J Exp Med. 2005;202:181–192. doi: 10.1084/jem.20050449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Parolini S, Bottino C, Falco M, Augugliaro R, Giliani S, Franceschini R, Ochs HD, Wolf H, Bonnefoy JY, Biassoni R, Moretta L, Notarangelo LD, Moretta A. X-linked lymphoproliferative disease. 2B4 molecules displaying inhibitory rather than activating function are responsible for the inability of natural killer cells to kill Epstein-Barr virus-infected cells. J Exp Med. 2000;192:337–346. doi: 10.1084/jem.192.3.337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Tangye SG, Phillips JH, Lanier LL, Nichols KE. Functional requirement for SAP in 2B4-mediated activation of human natural killer cells as revealed by the X-linked lymphoproliferative syndrome. J Immunol. 2000;165:2932–2936. doi: 10.4049/jimmunol.165.6.2932. [DOI] [PubMed] [Google Scholar]
- 11.Dupre L, Andolfi G, Tangye SG, Clementi R, Locatelli F, Arico M, Aiuti A, Roncarolo MG. SAP controls the cytolytic activity of CD8+ T cells against EBV-infected cells. Blood. 2005;105:4383–4389. doi: 10.1182/blood-2004-08-3269. [DOI] [PubMed] [Google Scholar]
- 12.Sharifi R, Sinclair JC, Gilmour KC, Arkwright PD, Kinnon C, Thrasher AJ, Gaspar HB. SAP mediates specific cytotoxic T cell functions in X-linked lymphoproliferative disease. Blood. 2004;103:3821–3827. doi: 10.1182/blood-2003-09-3359. [DOI] [PubMed] [Google Scholar]
- 13.Nichols KE, Hom J, Gong SY, Ganguly A, Ma CS, Cannons JL, Tangye SG, Schwartzberg PL, Koretzky GA, Stein PL. Regulation of NKT cell development by SAP, the protein defective in XLP. Nat Med. 2005;11:340–345. doi: 10.1038/nm1189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Chung B, Aoukaty A, Dutz J, Terhorst C, Tan R. Signaling lymphocytic activation molecule-associated protein controls NKT cell functions. J Immunol. 2005;174:3153–3157. doi: 10.4049/jimmunol.174.6.3153. [DOI] [PubMed] [Google Scholar]
- 15.Pasquier B, Yin L, Fondaneche MC, Relouzat F, Bloch-Queyrat C, Lambert N, Fischer A, de Saint-Basile G, Latour S. Defective NKT cell development in mice and humans lacking the adapter SAP, the X-linked lymphoproliferative syndrome gene product. J Exp Med. 2005;201:695–701. doi: 10.1084/jem.20042432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Eberl G, Lowin-Kropf B, MacDonald HR. Cutting edge: NKT cell development is selectively impaired in Fyn- deficient mice. J Immunol. 1999;163:4091–4094. [PubMed] [Google Scholar]
- 17.Gadue P, Morton N, Stein PL. The Src family tyrosine kinase Fyn regulates natural killer T cell development. J Exp Med. 1999;190:1189–1196. doi: 10.1084/jem.190.8.1189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Schmitt TM, Zuniga-Pflucker JC. Induction of T cell development from hematopoietic progenitor cells by delta-like-1 in vitro. Immunity. 2002;17:749–756. doi: 10.1016/s1074-7613(02)00474-0. [DOI] [PubMed] [Google Scholar]
- 19.Pear WS, Nolan GP, Scott ML, Baltimore D. Production of high-titer helper-free retroviruses by transient transfection. Proc Natl Acad Sci U S A. 1993;90:8392–8396. doi: 10.1073/pnas.90.18.8392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Nichols KE, Ma CS, Cannons JL, Schwartzberg PL, Tangye SG. Molecular and cellular pathogenesis of X-linked lymphoproliferative disease. Immunol Rev. 2005;203:180–199. doi: 10.1111/j.0105-2896.2005.00230.x. [DOI] [PubMed] [Google Scholar]
- 21.Griewank K, Borowski C, Rietdijk S, Wang N, Julien A, Wei DG, Mamchak AA, Terhorst C, Bendelac A. Homotypic interactions mediated by Slamf1 and Slamf6 receptors control NKT cell lineage development. Immunity. 2007;27:751–762. doi: 10.1016/j.immuni.2007.08.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Bezbradica JS, Hill T, Stanic AK, Van Kaer L, Joyce S. Commitment toward the natural T (iNKT) cell lineage occurs at the CD4+8+ stage of thymic ontogeny. Proc Natl Acad Sci U S A. 2005;102:5114–5119. doi: 10.1073/pnas.0408449102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Egawa T, Eberl G, Taniuchi I, Benlagha K, Geissmann F, Hennighausen L, Bendelac A, Littman DR. Genetic evidence supporting selection of the Valpha14i NKT cell lineage from double-positive thymocyte precursors. Immunity. 2005;22:705–716. doi: 10.1016/j.immuni.2005.03.011. [DOI] [PubMed] [Google Scholar]
- 24.Godfrey DI, Berzins SP. Control points in NKT-cell development. Nat Rev Immunol. 2007;7:505–518. doi: 10.1038/nri2116. [DOI] [PubMed] [Google Scholar]
- 25.Matsuda JL, Gapin L, Sidobre S, Kieper WC, Tan JT, Ceredig R, Surh CD, Kronenberg M. Homeostasis of V alpha 14i NKT cells. Nat Immunol. 2002;3:966–974. doi: 10.1038/ni837. [DOI] [PubMed] [Google Scholar]
- 26.Bezbradica JS, Gordy LE, Stanic AK, Dragovic S, Hill T, Hawiger J, Unutmaz D, Van Kaer L, Joyce S. Granulocyte-Macrophage Colony-Stimulating Factor Regulates Effector Differentiation of Invariant Natural Killer T Cells during Thymic Ontogeny. Immunity. 2006;25:487–497. doi: 10.1016/j.immuni.2006.06.017. [DOI] [PubMed] [Google Scholar]
- 27.Besseyrias V, Fiorini E, Strobl LJ, Zimber-Strobl U, Dumortier A, Koch U, Arcangeli ML, Ezine S, Macdonald HR, Radtke F. Hierarchy of Notch-Delta interactions promoting T cell lineage commitment and maturation. J Exp Med. 2007;204:331–343. doi: 10.1084/jem.20061442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Simarro M, Lanyi A, Howie D, Poy F, Bruggeman J, Choi M, Sumegi J, Eck MJ, Terhorst C. SAP increases FynT kinase activity and is required for phosphorylation of SLAM and Ly9. Int Immunol. 2004;16:727–736. doi: 10.1093/intimm/dxh074. [DOI] [PubMed] [Google Scholar]
- 29.Gadue P, Yin L, Jain S, Stein PL. Restoration of NK T cell development in fyn-mutant mice by a TCR reveals a requirement for Fyn during early NK T cell ontogeny. J Immunol. 2004;172:6093–6100. doi: 10.4049/jimmunol.172.10.6093. [DOI] [PubMed] [Google Scholar]
- 30.Cannons JL, Yu LJ, Jankovic D, Crotty S, Horai R, Kirby M, Anderson S, Cheever AW, Sher A, Schwartzberg PL. SAP regulates T cell-mediated help for humoral immunity by a mechanism distinct from cytokine regulation. J Exp Med. 2006;203:1551–1565. doi: 10.1084/jem.20052097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.McCausland MM, Yusuf I, Tran H, Ono N, Yanagi Y, Crotty S. SAP regulation of follicular helper CD4 T cell development and humoral immunity is independent of SLAM and Fyn kinase. J Immunol. 2007;178:817–828. doi: 10.4049/jimmunol.178.2.817. [DOI] [PubMed] [Google Scholar]