

Abstract
Aer, the Escherichia coli aerotaxis (oxygen-sensing) receptor is representative of a small class of receptors that face the cytoplasm in bacteria. Instead of sensing oxygen directly, Aer detects redox changes in the electron transport system or cytoplasm when the bacteria enter or leave a hypoxic micro-niche. As a result, Aer sensing also enables bacteria to avoid environments where carbon deficiency, unfavourable reduction potential or other insults would limit energy production. An FAD-binding PAS domain is the sensor for Aer and a HAMP domain interacts with the PAS domain to form an input-output module for signal transduction. By analogy to the first solution structure of an isolated HAMP domain from Archaeoglobus, Aer HAMP is proposed to fold into a four-helix bundle that rotates between a signal-on and signal-off conformation. Aer is the first protein in which a PAS-HAMP input-output module has been investigated. The structure and signal transduction mechanism of Aer is providing important insights into signalling by PAS and HAMP domains.
Keywords: Aerotaxis, chemotaxis, PAS domains, HAMP domains, signal transduction, receptors
a. Introduction
The Aer (aerotaxis) receptor in Escherichia coli senses redox and oxygen inside the cell, enabling the bacteria to respond to changes in internal energy (Taylor et al., 1999). The sensing module is a PAS (Per-Arnt-Sim) domain, and the signal output is used by E. coli to navigate to microenvironments where the bacteria can support optimal energy production. The Aer aerotaxis (oxygen-sensing) response is a simple behavioural system that is being elucidated at the molecular level, and is also a model for how cells can sense their own internal energy. Aer has a HAMP (histidine kinases, adenylyl cyclases, methyl-accepting chemotaxis proteins, phosphatases) domain, and the PAS and HAMP domains function together as an input-output module for signal transduction (Buron-Barral et al., 2006; Ma et al., 2005)). This is important. More than 14,900 PAS domains and 8,300 HAMP domains have been identified in sensory proteins, which ranks them among the more frequently reported protein domains. However, there is limited knowledge of the role of these modules in sensory transduction systems, and almost nothing was known previously about how PAS and HAMP domains function together.
Typical receptors, such as the E. coli chemoreceptor, Tsr, have external (periplasmic) sensing domains, transmembrane segments that transmit a signal from the outside sensor to the inside of the cell, and cytoplasmic output domains (Fig. 1). In contrast, Aer has an FAD-binding PAS domain in the cytoplasm, joined to a membrane anchor by the F1 linker (Bibikov et al., 1997; Bibikov et al., 2000; Repik et al., 2000) (Fig. 1). The anchor consists of two transmembrane segments separated by four residues in the periplasm (Amin et al., 2006). A HAMP domain links the second transmembrane sequence to the C-terminal signalling domain (Ma, 2001). The signalling domains in Aer and the chemoreceptors are highly conserved, and regulate the same chemotaxis histidine kinase system [see (Wadhams and Armitage, 2004) for a review] (Fig. 1). The structural unit of Aer and chemoreceptors is a homodimer. The signalling domain of the Tsr chemoreceptor has an extended helix-turn-helix configuration that forms a four-helix bundle in the dimer (Fig. 1) (Kim et al., 1999). The HAMP domain was first projected to be a linear helix that extended from the proximal helix of the signalling domain to the transmembrane segment (Kim et al., 1999). Subsequent high resolution electron microscopy indicated that the intact Tsr receptor is shorter than predicted by the extended helix model (Weis et al., 2003). Figure 1 illustrates two possible structures for HAMP domains: an extended helix (Kim et al., 1999) or a four-helix bundle (Hulko et al., 2006).
Fig. 1.

Comparison of the E. coli aerotaxis receptor, Aer, and the Tsr chemoreceptor, an alternative receptor for aerotaxis. The HAMP domain is shown in Tsr as an extended helix and in Aer as a four-helix bundle to illustrate different models for HAMP structure. Receptors shown are in silico constructions modeled from resolved structures and disulfide crosslinking studies: Aer-PAS [blue, (Key et al., 2007)]; F-1 (cyan); Aer-transmembrane [yellow, (Amin et al., 2006)]; Aer-HAMP [green, (Hulko et al., 2006)]; Aer-signalling [red, (Park et al., 2006)]. Tsr coordinates are from S-H. Kim as described in (Chi et al., 1997; Kim et al., 1999). The chemotaxis signalling pathway that controls flagellar rotation is also shown. Abbreviations: A, CheA protein; W, CheW; Y, CheY, Z, CheZ; P, phosphate; IM, inner membrane; OM, outer membrane.
b. Navigating with an energy/redox sensor
Some of the earliest studies of aerotaxis in bacteria demonstrated that, in a gradient of oxygen, microaerophilic bacteria always migrate to form a band where oxygen concentration is low, and aerobic bacteria accumulate where oxygen concentration is high (Beijerinck, 1893; Taylor et al., 1999). In some aquatic environments, bacteria accumulate in horizontal veils; each species navigates to the precise oxygen concentration, or reduction potential, that is optimal for growth (Taylor et al., 1999). Similarly, oxygen concentration is one of the ecological parameters that determine where commensal and virulent bacteria can establish colonies in mammals. As the mechanism of aerotaxis is investigated in different species, it is becoming clear that multiple strategies are utilized by bacteria to sense oxygen, but E. coli aerotaxis is the system that is understood best at the molecular level (Hou et al., 2000; Taylor et al., 1999).
E. coli does not sense oxygen directly; aerotaxis requires a functional electron transport system (Taylor et al., 1999). In response to an increase or decrease in oxygen, the respiratory components undergo oxidation or reduction, and the proton motive force changes accordingly. Aer and the serine chemoreceptor, Tsr, could sense the change in electron transport by sensing a change in redox potential, proton motive force, or possibly electron flux. These respiratory components are closely coupled, making it difficult to determine which parameter is the stimulus for E. coli aerotaxis. Recently, the parameters sensed by Aer and Tsr were successfully identified using electron transport system mutants that have different H+/e- ratios (Edwards et al., 2006). That is, each mutant differed in the number of H+ atoms extruded by the electron transport system per electron that traverses the pathway. The flux of electrons through the pathway was similar for all mutants, indicating it was not the stimulus for aerotaxis. Tsr-mediated aerotaxis correlated directly with the jump in proton motive force when anaerobic E. coli were exposed to air (Fig. 2) (Edwards et al., 2006). On the other hand, Aer-mediated responses did not correlate with the jump in proton motive force. These and other findings indicate that Tsr senses changes in proton motive force and Aer directly, or indirectly, senses redox changes in the electron transport system. Further studies are needed to establish the precise sensing mechanism of Aer.
Fig. 2.

Comparison of the aerotaxis response of Aer- and Tsr- expressing strains as a function of the step increase in Δψ (ΔψO2 - ΔψN2, the jump in proton motive force) when anaerobic bacteria are exposed to air [modified from (Edwards et al., 2006)]. Abbreviation: r2, coefficient of determination
The ability to sense proton motive force or redox potential provides E. coli with a measure of internal energy. In aerobic bacteria, proton motive force is the major source of energy for ATP synthesis, active transport, and motility (Harold and Maloney, 1996). Homeostasis in cells ceases in the absence of a membrane potential. By sensing proton motive force or redox, rather than oxygen per se, E. coli is able to navigate away from any environmental insults that threaten energy production (Taylor et al., 1999). The threats include carbon starvation, respiratory poisons, hypoxia, and extremes in pH and redox. A universal receptor that can measure internal redox potential or proton motive force in all cells has long been hypothesized (Glagolev, 1980; Taylor et al., 1979), but Aer and Tsr were the first such receptors shown to sense internal energy (Bibikov et al., 1997; Rebbapragada et al., 1997). If there is a decline in internal energy, aerotaxis guides the bacteria to a more favourable environment where higher energy levels can be supported.
c. Signal excitation in the PAS domain
The PAS domain is the sensory input site for Aer (Bibikov et al., 2000; Rebbapragada et al., 1997; Repik et al., 2000). In the PAS superfamily, PAS domains are cytoplasmic sensing modules of signal transduction proteins in archaea, bacteria, and eukarya (Taylor and Zhulin, 1999). Stimuli recognized by PAS domains include light, oxygen, redox and small molecules. Biological processes that utilize PAS domains include global regulation of metabolism, behaviour and development in prokaryotes, and circadian rhythms, response to hypoxia, ion channel function, and development in eukaryotes. The specificity in sensing is determined, in part, by cofactors that are associated with PAS domains: 4-hydroxycinnamic acid in photoactive yellow protein (PYP)(Pellequer et al., 1998); a heme in the oxygen sensor, FixL (Gong et al., 1998); FAD in Aer and NifL (Bibikov et al., 1997; Crosson and Moffat, 2001; Repik et al., 2000); and flavin mononucleotide (FMN) in the plant NPH1 photoreceptor that also senses redox (Christie et al., 1999). Some PAS domains may not have a cofactor: the crystal structure of the PAS domain from the HERG cardiac K+ channel lacks a cofactor (Morais Cabral et al., 1998).
The PAS structure, resolved for more than 15 proteins, including PYP, FixL, HERG, NifL and NPH1, has a distinctive αβ fold with a five-stranded antiparallel β-sheet core (Pellequer et al., 1998). The structure is also described as a left-handed glove in which the fingers enclose a pocket that binds the cofactor. The fingers of the glove are formed primarily from the β-sheet, the palm from α-helical loops and the thumb from β-strands (Gong et al., 1998). The common structure of PAS domains could be associated with similar mechanisms for signalling, but different cofactors in the PAS fold are likely to introduce significant variation in the signalling pathways. The in silico modelling of the Aer structure using the coordinates of resolved PAS structures has been very successful in advancing investigations of the Aer PAS domain. The Azotobacter vinelandii NifL and Neurospora crassa Vivid PAS domains were resolved recently with FAD cofactors (Key et al., 2007; Zoltowski et al., 2007) and the NifL coordinates were used to further refine the Aer model to include a putative position of the FAD cofactor (Fig. 3).
Fig. 3.

In silico model of the Aer PAS domain, highlighting residues involved in aerotaxis. The cleft in which FAD binds is shown. Replacement of the residues shown produced a null aerotaxis phenotype (red), a loss of FAD binding and a null phenotype (green), an inverted response (yellow), and a CW-signalling bias (magenta).
Critical residues for signalling by the Aer PAS domain have been identified (Bibikov et al., 2000; Buron-Barral et al., 2006; Repik et al., 2000; Watts et al., 2006b). Null Aer mutants have a signal-off conformation that produces a counterclockwise (CCW) rotational bias of the flagellar motors. The signal from Aer PAS enhances the signal-on conformation of the signalling domain (Fig. 1), imposing a clockwise (CW) bias on the motors. Thirteen cysteine PAS mutants had defective input-output control and were not rescued by simultaneous production of the Tar, Trg, and Tap chemoreceptors (Repik et al., 2000; Watts et al., 2006b). Cysteine replacements at Arg57, His58, and Asp60 abolished FAD binding to Aer. These residues surround the pocket in which FAD is predicted to bind (Fig 3). Residues Arg57, His58 and Asp60 are unique to the Aer _ PAS (FAD-binding) subfamily and are conserved in members of the subfamily, but not in other PAS domains (L. Ulrich, W. Black and I. Zhulin, personal communication). This suggests that they bind to FAD or are structural residues that determine the shape of the binding site. Although these particular mutants are stable, some null Aer mutants are unstable and give a false-negative result for FAD-binding. The lesions in those mutants are usually not associated with the FAD-binding site (Buron-Barral et al., 2006) Tyr111 also projects into the putative FAD pocket (Fig. 3) and the Aer-Y111C mutant has an inverse response in a temporal oxygen gradient (Repik et al., 2000). E. coli cells producing wild-type Aer give a positive (CCW) response to an increase in oxygen, whereas cells producing Aer-Y111C display a negative (CW) response. A putative signalling pathway in Aer is proposed to include residues in contact with the isoallaxazine ring of FAD that transduce redox changes in FAD into conformational changes in the PAS domain. The latter are then transmitted to downstream components of the pathway. The inverted aerotaxis response caused by the Y111C substitution can be explained by a 3-state model, where fully oxidized and reduced forms of FAD generate a CW signal, and the semiquinone generates a CCW signal. Normally, aerobic/anaerobic changes produce the semiquinone/quinol forms, but the mutation might shift the redox potential of the FAD cofactor so that it is fully oxidized during maximal rates of electron transport (Repik et al., 2000; Watts et al., 2006b).
The Aer PAS domain has a hot spot for mutations at the junction of the N-terminal cap (N-cap) and the PAS core, suggesting residues Thr19, Leu20, Met21, and Ser22 are critical for Aer structure and/or signalling (Watts et al., 2006b). The resulting null mutant Aer proteins did not bind FAD, suggesting that the N-cap stabilizes FAD binding to the PAS domain. Substitutions at Leu20 and Met21 are dominant, indicating that the defective receptors disrupt both signalling within the Aer dimer and interactions with neighbouring dimers. The PAS N-cap from HERG and PYP also have an important role in signalling. Deleting part of the N-cap from the PAS domain of the PYP photoreceptor prolonged the lifetime of the active form of PYP (Imamoto et al., 2002; van der Horst et al., 2001). Deleting the PAS N-cap from the HERG voltage-dependent K+ channel had a similar effect on the kinetics of channel deactivation as removing the entire PAS domain (Morais Cabral et al., 1998).
Two classes of mutations in the PAS domain might interrupt the aerotaxis pathway from FAD to the downstream components (Buron-Barral et al., 2006; Repik et al., 2000). The first class results in proteins that bind FAD but have an extreme CCW bias (e.g., M21D, L20K, G42C, W79C, G90C in Fig 3). The second class includes those with a gain-of-function mutation that results in a strong signal-on (CW) bias (e.g., L20C, M34C, F66C, N85C). These variant proteins bind FAD, are not rescued by chemoreceptors, and are present at normal levels (Fig. 3). The CW-biased mutants probably augment an interaction between the PAS and HAMP domains that promotes CW signal output [(Buron-Barral et al., 2006); see section d below].
PAS domains are present as dimers in other sensory proteins, but it is not known whether the Aer PAS domain forms stable dimers. The A. vinelandii NifL PAS-FAD domain crystallized as a dimer, in part due to a N-cap dimerization motif (A’α), which is also conserved in Aer (Key et al., 2007). In NifL, the A’α helix packs against the hydrophobic surface of the β-sheet on the opposing monomer. The Aer-PAS domain is predicted to have a similar PAS/PAS’ dimerization interface. The putative contact surfaces would be asymmetric and would not have been detectable by our earlier study of symmetric crosslinking surfaces in the Aer N-cap (Watts et al., 2006b).
The most significant gap in our understanding of the Aer signalling pathway is the mechanism by which PAS-FAD detects changes in the electron transport system when the concentration of oxygen increases or decreases. Two hybrid studies designed to identify direct interaction of PAS-FAD with a respiratory component, such as NADH dehydrogenase 1 (NDH-1), were unsuccessful. NDH-1 has an important role in Aer signalling but is not essential (Edwards et al., 2006). Another possibility is that PAS-FAD interacts with a quinone, NADH or free FAD/FADH from the electron transport system (Fig. 4). The purified Aer protein has a low affinity for FAD and it is possible that the FAD cofactor in the PAS domain readily exchanges with the cytoplasmic FAD/FADH pool.
Fig. 4.

Components that might influence Aer signalling activity. Abbreviations: Cyo, cytochrome bo; Cyd, cytochrome bd; Ndh1, NADH dehydrogenase 1; Ndh2, NADH dehydrogenase 2; Sdh, succinate dehydrogenase.
In E. coli, the NADH-dependent flavin reductase (Fre) reduces approximately 80% of all free cytosolic FAD (Woodmansee and Imlay, 2002). Purified FAD-bound Aer is reduced by Fre and an E. coli fre mutant is only weakly aerotactic in succinate soft agar (Webster and Xun, 2003), suggesting that Fre might be involved in the aerotaxis signalling pathway (Fig. 4). Expression of Fre from a plasmid or slight overproduction of Aer, however, restores normal aerotaxis (L. Xun and M.S. Johnson, unpublished observation). Higher Aer levels inhibited swarming in fre and wild-type cells. These results suggest that Fre contributes to the parameters sensed by Aer on succinate soft agar, but Aer does not require Fre expression to function. The effects of Fre are similar in a true oxygen gradient. In an open-ended capillary tube, E. coli respiration generates an oxygen gradient and the cells form a band near the meniscus at a preferred oxygen concentration (Taylor et al., 2001). Cells with Aer-mediated aerotaxis consistently banded further from the meniscus in the absence of Fre than when Fre was present. This indicated that the fre strain preferred a lower oxygen concentration. These results can be accommodated by the following hypothesis. Fre maintains the intracellular reducing potential within a range favourable to growth and aerotaxis. In the absence of Fre, the reduction potential of FAD/FADH is lower and the critical number of Aer receptors in a reduced, signal-on state is reached at a lower concentration of environmental oxygen. The number of reduced Aer receptors in the fre strain is less than the critical number for signalling at normal oxygen levels but can be increased by overproduction of Aer. This hypothesis can also explain the observation that, in a capillary, the band of E. coli cells moves closer to the meniscus when Aer is overproduced (Ma et al., 2005). In Tsr-mediated aerotaxis, which is not dependent on reduced flavin for signalling, the band position did not reproducibly shift in fre strains.
d. HAMP domain structure and function
The HAMP domain in E. coli chemoreceptors was initially considered a linker that relayed a mechanical displacement in the periplasmic sensor domain to the signalling domain (compare Fig. 1). The linker was shown to consist of two amphipathic helices (AS-1 and AS-2) connected by a sequence of unknown structure (Butler and Falke, 1998). The HAMP domain was recognized as a sensory transduction module after in silico analyses showed that the domain is widely conserved in sensory proteins that include histidine kinases, adenyl cyclases, methyl-accepting chemotaxis proteins and phosphatases (Aravind and Ponting, 1999; Williams and Stewart, 1999). The AS-2 helix, as defined, ends at residue C253 in Aer but the sequence is conserved in HAMP proteins through Aer residue 271, forming a proximal signalling domain (Ma, 2001; Ma et al., 2005) (Fig. 5A).
Fig. 5.

The Af1503 HAMP domain as a model for the Aer HAMP domain. A. Domain sub-structure of the Aer HAMP and proximal signalling domains showing gain-of-function residue substitutions (vertical lines below the HAMP diagram) that resulted in Aer proteins with a CW-biased signal output and bound FAD. Vertical lines above the HAMP diagram represent substitutions that resulted in a null aerotaxis phenotype, loss of FAD binding and defective maturation of Aer [Data from (Buron-Barral et al., 2006; Ma et al., 2005)]. B. Solution structure of the isolated Af1503 HAMP domain from A. fulgidus. C. The knobs-to-knobs packing geometry of the Af1503 HAMP domain is converted to a knobs-to-holes geometry by a 26° rotation of the HAMP helices. Compare the orientation of Ile residues (red). D. Proposed direction of rotation of interlocking helices in the Af1503 HAMP domain. (B, C, and D are modified from (Hulko et al., 2006). Used with permission).
Recently the tertiary fold of an isolated HAMP domain from the Archaeoglobus fulgidus Af1503 protein was resolved (Hulko et al., 2006). The AS-1 and AS-2 helices from the dimeric protein are folded into a parallel four-helix bundle with the connectors tightly packed into a groove between AS-1 and AS-2 (Fig. 5B). The packing of HAMP core residues deviates from the knobs-into-holes packing that defines a coiled-coil structure. The latter geometry specifies the packing of a core residue (knob) from one helix into a space surrounded by four residues from the facing helix (hole). In the Af1503 HAMP domain, the residues of the hydrophobic core have knobs-to-knobs packing with alternating layers in which hydrophobic side chains point at the central supercoil axis in one layer, and point sideways to form an interacting ring of residues enclosing the central cavity in the alternating layer (Fig. 5C) (Hulko et al., 2006).
The solution structure of the Af1503 HAMP domain is of great interest, but there is some concern that the structure of an Af1503 HAMP domain may be atypical and not resemble the conformation of other HAMP domains.The Af1503 HAMP domain is normally tethered to a transmembrane domain in vivo but not to a signalling domain, as are most HAMP domains, and it has no known signalling function. However, there are now some indications that the structure of other HAMP domains may be similar to the Af1503 HAMP domain. Models based on the coordinates of the Af1503 HAMP structure have been developed for the HAMP domains from Aer (K. Watts, unpublished data) and the Salmonella Tar chemoreceptor (K. Swain and J. Falke, personal communication). Investigations are currently underway to test these in silico models. In crosslinking studies, the cysteine replacements with the most extensive crosslinking are those that collide most often and are therefore expected to be at the dimer interface. The residues on the AS-1 and AS-2 dimer interface of the Aer model were similar to the residues predicted by in vivo crosslinking to be on the interface (K. Watts, unpublished observation), and mutations that disrupt Aer stability and maturation would be predicted to disrupt the four-helix structure (K. Watts, unpublished finding).
One of the properties that makes HAMP domains so interesting is their versatility. The ligand-binding domains of E. coli chemoreceptors generate a mechanical signal, an inward displacement of the second transmembrane segment that is perpendicular to the plane of the membrane (Falke and Hazelbauer, 2001; Milburn et al., 1991). On the other hand, in Aer, the PAS domain interacts directly with the HAMP domain so that the signal is transmitted through the cytoplasm and parallel to the plane of the membrane. The strongest evidence for a direct PAS-HAMP interaction is from allele-specific suppression in intragenic pseudoreversion analysis (Watts et al., 2004). Specifically, a missense mutation in the HAMP domain (C253R) is exclusively suppressed by a missense mutation in the PAS domain (N34D), indicating that these residues are in close proximity in the Aer protein (Watts et al., 2004). It remains to be determined how a HAMP domain can transform different signals from chemoreceptors and Aer into a common output signal for the similar signalling domains in these proteins. Considering that there are more than 8,300 HAMP domains, it is likely that HAMP domains have adapted to process a variety of different transduction signals.
e. PAS-HAMP as input-output module
There is increasing evidence that the PAS and HAMP domains function together as the input-output module for Aer. Chimeric receptors were constructed by fusing the PAS domain of Aer to the signalling domain of the Tar or Tsr chemoreceptors (Bibikov et al., 2000; Repik et al., 2000). The chimeric receptors were competent for signalling only if the PAS and HAMP domains were both derived from Aer. This suggests that specific contact surfaces are required for signalling between the domains. Mutations in the HAMP domain can affect FAD binding to the PAS domain and maturation of the Aer protein (Bibikov et al., 2000; Buron-Barral et al., 2006; Ma et al., 2005). Folding of the nascent Aer peptide into the native protein is a complex process. The nascent peptide is sequestered in the GroEL chaperone protein until critical structural determinants are correctly assembled (Herrmann et al., 2004). The determinants include the transmembrane sequences and AS-1. To complete folding of the PAS domain, AS-2 is also required and interaction of the HAMP and PAS domains is postulated. Aer folding has been investigated by comparing proteolytic degradation of variant Aer proteins that have residue replacements in the HAMP domain. Some variants exist as a mixture of native and partially unfolded forms (Buron-Barral et al., 2006). There are varying degrees of stability of the mature protein, which presumably depend on how far the folding process has progressed. A critical step in maturation is FAD binding. FAD deficiency could result from failure of the Aer protein to complete the folding process, or alternatively, FAD binding might be required before folding can be completed.
Most HAMP mutations that prevent FAD binding are suppressed by three non-specific suppressor mutations that map in the PAS domain (Watts et al., 2004). These suppressors restore FAD binding, signalling and stability to null HAMP mutants (Buron-Barral et al., 2006; Watts et al., 2004). By analogy to the NifL PAS domain, the suppressors (S28G, A65V, A99V) are predicted to surround the FAD binding pocket and might increase the affinity of the PAS domain for FAD (Fig. 6). The clustering of critical residues and suppressor mutations around the putative FAD binding site (Fig. 6) supports a role for FAD binding in Aer maturation. We propose that interactions between HAMP AS-2 and the PAS domain stabilize folding of the PAS domain into the FAD-binding conformation and that FAD binding is necessary for completion of the maturation process. Once the native fold is formed, it is relatively stable and FAD can be removed by dialysis and replaced without significant degradation of the protein (M.S. Johnson and S. L. Fry, unpublished observation).
Fig. 6.
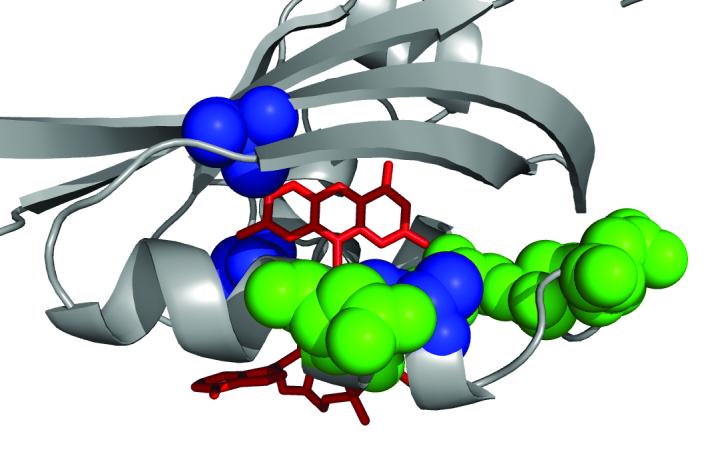
A view of the FAD-binding cavity in Aer PAS showing residues critical for FAD binding (green), and location of general suppressors (blue) of HAMP[AS-2] null mutants. FAD coordinates are modeled on the crystal structure of the NifL protein.
The lesions that affect Aer maturation are distributed throughout the HAMP domain (Fig. 5A) (Buron-Barral et al., 2006). However, it is likely that interactions required for Aer maturation are between PAS and HAMP-AS-2. In addition to allele-specific suppression between AS-2 and PAS (Watts et al., 2004), there are conserved residues in all AS-2 sequences of Aer-like proteins, more so than in the HAMP AS-1 and connector. Our current hypothesis is that maturation requires a four-helix HAMP structure to interact with the nascent PAS domain. Lesions in AS-1 and the connector that interrupt maturation are proposed to disrupt the four-helix HAMP structure.
In addition to PAS-HAMP contacts that are required for maturation, there is a second class of interaction in which the PAS domain promotes formation of the kinase-on (CW) conformation of the HAMP domain (Fig. 5A) (Buron-Barral et al., 2006; Ma et al., 2005). Gain-of-function mutants with a CW-bias in signalling have lesions in the PAS or HAMP domain that augment the PAS-HAMP interaction that is responsible for promoting CW signalling output. Such lesions in the HAMP domain are focused at the AS-1/connector junction and AS-2/proximal signalling junction. In the folded four-helix HAMP structure, the CW lesions from AS-1 and AS-2 are grouped around the distal end of the HAMP domain and likely define the contact domain where the PAS domain promotes CW signalling by the HAMP domain. The PAS domain appears to interact with the HAMP domain from the cognate subunit in the Aer homodimer (Watts et al., 2006a).
Null mutants with defects in the proximal signalling domain differ from those with defects in the HAMP domain. The proximal signalling mutants have higher levels of Aer, bind FAD and are dominant over wild-type Aer (Buron-Barral et al., 2006; Ma et al., 2005). Some defects are rescued in the presence of the Tar or Tsr chemoreceptors, and these defects might have less severe structural effects on maturation. Other lesions are not rescued and are more likely to affect determinants or structures for PAS/HAMP signalling directly.
There are further clues to the signal transduction mechanism in the PAS-HAMP module. The knobs-into-holes and knobs-to-knobs packing geometries predicted for the Af1503 HAMP domain are nearly isoenergetic and could be interconverted by a concerted rotation of 26° in all four helices (Fig. 5C, D) (Hulko et al., 2006). The interdigitation of side chains gives this HAMP rotation the characteristics of a gear box with four cogwheels, where neighbouring helices must rotate in opposite directions. The four helices of the Aer HAMP domain might also rotate between the kinase-on and kinase-off state. If so, changes in the redox state of the isoalloxazine ring of the FAD co-factor in the PAS domain must trigger an appropriate HAMP rotation, presumably through contact of the PAS domain with the distal end of the four-helix HAMP domain (Fig. 5A) (Ma et al., 2005; Watts et al., 2004). However, it is unlikely that the final output from the signalling domain is a rotation. In a fluorescence polarization study of signalling by E. coli chemoreceptors, the motion of a yellow-fluorescent-protein probe was interpreted to be consistent with displacement but not rotation of the signalling domain (Vaknin and Berg, 2007). Signalling changed the spacing between chemoreceptor dimers within a trimer-of-dimers: attractants increased the spacing by 10% and repellents decreased the spacing. Flexing of the signalling domain is made possible by a glycine-hinge (Gly330 and Gly331 in Aer) within the flexible-bundle subdomain of the signalling domain (Coleman et al., 2005). Aer presumably uses a similar signalling mechanism because the signalling domains of chemoreceptors are interchangeable with the Aer signalling domain (Bibikov et al., 2000; Repik et al., 2000), and Aer can signal in mixed trimers-of-dimer teams with the other chemoreceptors (Gosink et al., 2006).
A mechanical (Lego©) model that simulates a four-helix domain is able to convert a gear-box rotation of the ‘helices’ into bending of an attached helix, if a flexible hinge is inserted at the distal end (R. Alexander and I. Zhulin, personal communication). It remains to be seen whether the mechanical model has relevance for HAMP proteins. The proximal signalling domain that adjoins the HAMP domain is a candidate for a hinge in Aer (Fig. 5A). Crosslinking data indicate that the helical structure of the proximal signalling domain is interrupted by a four-residue loop. The concept of bending is conjecture at this time but torque conversion of a HAMP signal into displacement of the signalling domain is a fruitful area for further investigation in Aer, and in chemoreceptors, to determine whether torque conversion is a general principle in these sensory systems..
f. Organization of the Aer receptor
The basic building block of the Aer protein is a homodimer (Ma et al., 2004). Domain interactions that transduce the aerotaxis signal could occur in cis (within one subunit), or in trans (between cognate subunits). To address this question, two Aer proteins with amino acid substitutions in different domains were expressed from compatible plasmids and intragenic complementation was examined (Watts et al., 2006a). Aer heterodimers can function with one wild-type PAS and HAMP sequence per dimer, provided the functional PAS and HAMP sequences are on different subunits. However, Aer signals from the PAS to the signalling domain of the same subunit. To confirm an intrasubunit signalling pathway, truncated Aer proteins were produced together with a full-length Aer-HAMP missense variant. Truncations that deleted most of the signalling domain (Aer1-285), or the PAS domain (Aer120-506) from the non-signalling subunit, did not disrupt aerotaxis of heterodimers (Fig. 7). However, two complete HAMP sequences were required for aerotaxis, even where one sequence had an amino acid substitution. This suggests that a four-helix HAMP structure is essential for signalling. Aer can therefore sense and signal with one PAS and one signalling domain per dimer, provided they are on the signalling subunit. However, as stated above, the AS-2 helix interacts with the cognate PAS domain to stabilize the native fold of Aer. These findings do not exclude the likelihood that signalling domains, and possibly PAS domains from heterodimers, dimerize by interacting with a second heterodimer.
Fig. 7.

The minimal sequence required for aerotaxis in an Aer heterodimer. A. The organisation and boundaries of domains relevant to the experiments described in B and C. B. For heterodimer experiments, mutant Aer constructs were expressed from compatible plasmids in BT3400 (aer, tsr, recA), an aerotaxis- and recombination-deficient E. coli strain. In succinate soft agar, aerotactic colonies, such as BT3400 expressing wild-type Aer, expand rapidly and form a defined ring at their edge. In contrast, non-aerotactic colonies (e.g., BT3400 expressing vector alone) stay trapped near the inoculation site. C. In succinate soft agar, colonies formed by cells expressing the homodimers of each mutant Aer protein were non-aerotactic (as shown on both sides of each cartoon), whereas colonies formed by cells expressing the heterodimers shown were aerotactic (as shown beneath each cartoon). Arrows indicate the proposed signalling pathway within each heterodimer. Heterodimer experiments, such as those shown, indicate that Aer requires one PAS and one signalling domain, but two HAMP domains, in order to support aerotaxis. Modified from (Watts et al., 2006a). Used with permission. Abbreviation: Sig., signalling.
Aer, like chemoreceptors (Kim et al., 2002), forms trimers-of-dimers, the functional signalling team for signal transduction (Gosink et al., 2006). Aer is a low abundance receptor, comprising only about 2.5% of the total receptor pool (D. Salcedo and M. S. Johnson, unpublished observation), but Aer apparently enhances signalling by forming mixed trimers-of-dimers with the high abundance receptors Tsr and Tar (Gosink et al., 2006). An important unknown is whether the abundant receptors have a role in adaptation of Aer to aerotaxis stimuli. Aer lacks the glutamyl-methylation system that enables Tar and Tsr to adapt to stimuli (Bibikov et al., 2004; Niwano and Taylor, 1982). However, Aer can function in E. coli without chemoreceptor assistance, provided that it is produced at high enough levels to establish a suitable prestimulus swimming pattern.
g. Summary
Aer and the aerotaxis system enable E. coli to navigate to environments that support optimal energy production. Aer is a cytoplasmic receptor that senses changes in the redox state of the electron transport system via an FAD-containing PAS domain. The PAS and HAMP domains constitute an input-output module that transduces FAD reduction into a signal that promotes a signal-on conformational change in the HAMP domain. The emerging paradigm for Aer includes a four-helix HAMP domain which is a versatile signal-transduction module that might respond to diverse stimuli by rotating interlocking helices. The HAMP output is probably not a rotation but might flex the signalling domains of Aer and chemoreceptor proteins.
Acknowledgements
I thank I. B. Zhulin and J. Falke for communicating findings prior to publication, and S-H Kim for providing the coordinates for the Tsr model.. M.S. Johnson and K.J. Watts provided data prior to publication, prepared figures, and critically reviewed the manuscript. The work in my laboratory is supported by an award (GM0297481) from the Institute of General Medical Sciences.
Abbreviations
- CW
clockwise
- CCW
counterclockwise
References
- Amin DN, Taylor BL, Johnson MS. Topology and boundaries of the aerotaxis receptor Aer in the membrane of Escherichia coli. J Bacteriol. 2006;186:894–901. doi: 10.1128/JB.188.3.894-901.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aravind L, Ponting CP. The cytoplasmic helical linker domain of receptor histidine kinase and methyl-accepting proteins is common to many prokaryotic signalling proteins. FEMS Microbiol Lett. 1999;176:111–116. doi: 10.1111/j.1574-6968.1999.tb13650.x. [DOI] [PubMed] [Google Scholar]
- Beijerinck MW. Ueber Atmungsfiguren beweglicher Bakterien. Zentrabl Bakteriol Parasitenkd. 1893;14:827–845. [Google Scholar]
- Bibikov SI, Biran R, Rudd KE, Parkinson JS. A signal transducer for aerotaxis in Escherichia coli. J Bacteriol. 1997;179:4075–4079. doi: 10.1128/jb.179.12.4075-4079.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bibikov SI, Barnes LA, Gitin Y, Parkinson JS. Domain organization and flavin adenine dinucleotide-binding determinants in the aerotaxis signal transducer Aer of Escherichia coli. Proc Natl Acad Sci U S A. 2000;97:5830–5835. doi: 10.1073/pnas.100118697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bibikov SI, Miller AC, Gosink KK, Parkinson JS. Methylation-independent aerotaxis mediated by the Escherichia coli Aer protein. J Bacteriol. 2004;186:3730–3737. doi: 10.1128/JB.186.12.3730-3737.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buron-Barral M, Gosink KK, Parkinson JS. Loss- and gain-of-function mutations in the F1-HAMP region of the Escherichia coli aerotaxis transducer Aer. J Bacteriol. 2006;188:3477–3486. doi: 10.1128/JB.188.10.3477-3486.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Butler SL, Falke JJ. Cysteine and disulfide scanning reveals two amphiphilic helices in the linker region of the aspartate chemoreceptor. Biochemistry. 1998;37:10746–10756. doi: 10.1021/bi980607g. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chi YI, Yokota H, Kim SH. Apo structure of the ligand-binding domain of aspartate receptor from Escherichia coli and its comparison with ligand-bound or pseudoligand-bound structures. FEBS Lett. 1997;414:327–332. doi: 10.1016/s0014-5793(97)01027-2. [DOI] [PubMed] [Google Scholar]
- Christie JM, Salomon M, Nozue K, Wada M, Briggs WR. LOV (light, oxygen, or voltage) domains of the blue-light photoreceptor phototropin (nph1): binding sites for the chromophore flavin mononucleotide. Proc Natl Acad Sci U S A. 1999;96:8779–8783. doi: 10.1073/pnas.96.15.8779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coleman MD, Bass RB, Mehan RS, Falke JJ. Conserved glycine residues in the cytoplasmic domain of the aspartate receptor play essential roles in kinase coupling and on-off switching. Biochemistry. 2005;44:7687–7695. doi: 10.1021/bi0501479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crosson S, Moffat K. Structure of a flavin-binding plant photoreceptor domain: insights into light-mediated signal transduction. Proc Natl Acad Sci U S A. 2001;98:2995–3000. doi: 10.1073/pnas.051520298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Edwards JC, Johnson MS, Taylor BL. Differentiation between electron transport sensing and proton motive force sensing by the Aer and Tsr receptors for aerotaxis. Mol Microbiol. 2006;62:823–837. doi: 10.1111/j.1365-2958.2006.05411.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Falke JJ, Hazelbauer GL. Transmembrane signaling in bacterial chemoreceptors. Trends Biochem Sci. 2001;26:257–265. doi: 10.1016/s0968-0004(00)01770-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glagolev AN. Reception of the energy level in bacterial taxis. J Theor Biol. 1980;82:171–185. doi: 10.1016/0022-5193(80)90097-1. [DOI] [PubMed] [Google Scholar]
- Gong W, Hao B, Mansy SS, Gonzalez G, Gilles-Gonzalez MA, Chan MK. Structure of a biological oxygen sensor: a new mechanism for heme-driven signal transduction. Proc Natl Acad Sci U S A. 1998;95:15177–15182. doi: 10.1073/pnas.95.26.15177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gosink KK, del Carmen Buron-Barral M, Parkinson JS. Signaling interactions between the aerotaxis transducer Aer and heterologous chemoreceptors in Escherichia coli. J Bacteriol. 2006;188:3487–3493. doi: 10.1128/JB.188.10.3487-3493.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harold FM, Maloney PC. Energy transduction by ion currents. In: Neidhardt FC, editor. Escherichia coli and Salmonella: Cellular and Molecular Biology. Vol. 1. ASM Press; Washington, D.C.: 1996. pp. 283–306. [Google Scholar]
- Herrmann S, Ma Q, Johnson MS, Repik AV, Taylor BL. PAS domain of the Aer redox sensor requires C-terminal residues for native-fold formation and flavin adenine dinucleotide binding. J Bacteriol. 2004;186:6782–6791. doi: 10.1128/JB.186.20.6782-6791.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hou S, Larsen RW, Boudko D, Riley CW, Karatan E, Zimmer M, Ordal GW, Alam M. Myoglobin-like aerotaxis transducers in Archaea and Bacteria. Nature. 2000;403:540–544. doi: 10.1038/35000570. [DOI] [PubMed] [Google Scholar]
- Hulko M, Berndt F, Gruber M, Linder JU, Truffault V, Schultz A, Martin J, Schultz JE, Lupas AN, Coles M. The HAMP Domain Structure Implies Helix Rotation in Transmembrane Signaling. Cell. 2006;126:929–940. doi: 10.1016/j.cell.2006.06.058. [DOI] [PubMed] [Google Scholar]
- Imamoto Y, Kamikubo H, Harigai M, Shimizu N, Kataoka M. Light-induced global conformational change of photoactive yellow protein in solution. Biochemistry. 2002;41:13595–13601. doi: 10.1021/bi0264768. [DOI] [PubMed] [Google Scholar]
- Key J, Hefti M, Purcell EB, Moffat K. Structure of the Redox Sensor Domain of Azotobacter vinelandii NifL at Atomic Resolution: Signaling, Dimerization, and Mechanism(,) Biochemistry. 2007;46:3614–3623. doi: 10.1021/bi0620407. [DOI] [PubMed] [Google Scholar]
- Kim KK, Yokota H, Kim SH. Four-helical-bundle structure of the cytoplasmic domain of a serine chemotaxis receptor. Nature. 1999;400:787–792. doi: 10.1038/23512. [DOI] [PubMed] [Google Scholar]
- Kim SH, Wang W, Kim KK. Dynamic and clustering model of bacterial chemotaxis receptors: structural basis for signaling and high sensitivity. Proc Natl Acad Sci U S A. 2002;99:11611–11615. doi: 10.1073/pnas.132376499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma Q. Department of Microbiology and Molecular Genetics. Loma Linda University; Loma Linda, CA: 2001. HAMP domain and signaling mechanism of the Aer protein. [Google Scholar]
- Ma Q, Roy F, Herrmann S, Taylor BL, Johnson MS. The Aer protein of Escherichia coli forms a homodimer independent of the signaling domain and flavin adenine dinucleotide binding. J Bacteriol. 2004;186:7456–7459. doi: 10.1128/JB.186.21.7456-7459.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma Q, Johnson MS, Taylor BL. Genetic analysis of the HAMP domain of the Aer aerotaxis sensor localizes flavin adenine dinucleotide-binding determinants to the AS-2 helix. J Bacteriol. 2005;187:193–201. doi: 10.1128/JB.187.1.193-201.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Milburn MV, Prive GG, Milligan DL, Scott WG, Yeh J, Jancarik J, Koshland DE, Jr., Kim SH. Three-dimensional structures of the ligand-binding domain of the bacterial aspartate receptor with and without a ligand. Science. 1991;254:1342–1347. doi: 10.1126/science.1660187. [DOI] [PubMed] [Google Scholar]
- Cabral J.H. Morais, Lee A, Cohen SL, Chait BT, Li M, Mackinnon R. Crystal structure and functional analysis of the HERG potassium channel N terminus: a eukaryotic PAS domain. Cell. 1998;95:649–655. doi: 10.1016/s0092-8674(00)81635-9. [DOI] [PubMed] [Google Scholar]
- Niwano M, Taylor BL. Novel sensory adaptation mechanism in bacterial chemotaxis to oxygen and phosphotransferase substrates. Proc Natl Acad Sci U S A. 1982;79:11–15. doi: 10.1073/pnas.79.1.11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park SY, Borbat PP, Gonzalez-Bonet G, Bhatnagar J, Pollard AM, Freed JH, Bilwes AM, Crane BR. Reconstruction of the chemotaxis receptor-kinase assembly. Nat Struct Mol Biol. 2006;13:400–407. doi: 10.1038/nsmb1085. [DOI] [PubMed] [Google Scholar]
- Pellequer JL, Wager-Smith KA, Kay SA, Getzoff ED. Photoactive yellow protein: a structural prototype for the three-dimensional fold of the PAS domain superfamily. Proc Natl Acad Sci U S A. 1998;95:5884–5890. doi: 10.1073/pnas.95.11.5884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rebbapragada A, Johnson MS, Harding GP, Zuccarelli AJ, Fletcher HM, Zhulin IB, Taylor BL. The Aer protein and the serine chemoreceptor Tsr independently sense intracellular energy levels and transduce oxygen, redox, and energy signals for Escherichia coli behavior. Proc Natl Acad Sci U S A. 1997;94:10541–10546. doi: 10.1073/pnas.94.20.10541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Repik A, Rebbapragada A, Johnson MS, Haznedar JO, Zhulin IB, Taylor BL. PAS domain residues involved in signal transduction by the Aer redox sensor of Escherichia coli. Mol Microbiol. 2000;36:806–816. doi: 10.1046/j.1365-2958.2000.01910.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taylor BL, Miller JB, Warrick HM, Koshland DE., Jr. Electron acceptor taxis and blue light effect on bacterial chemotaxis. J Bacteriol. 1979;140:567–573. doi: 10.1128/jb.140.2.567-573.1979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taylor BL, Zhulin IB. PAS domains: internal sensors of oxygen, redox potential, and light. Microbiol Mol Biol Rev. 1999;63:479–506. doi: 10.1128/mmbr.63.2.479-506.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taylor BL, Zhulin IB, Johnson MS. Aerotaxis and other energy-sensing behavior in bacteria. Annu Rev Microbiol. 1999;53:103–128. doi: 10.1146/annurev.micro.53.1.103. [DOI] [PubMed] [Google Scholar]
- Taylor BL, Rebbapragada A, Johnson MS. The FAD-PAS domain as a sensor for behavioral responses in Escherichia coli. Antioxid Redox Signal. 2001;3:867–879. doi: 10.1089/15230860152665037. [DOI] [PubMed] [Google Scholar]
- Vaknin A, Berg HC. Physical responses of bacterial chemoreceptors. J Mol Biol. 2007;366:1416–1423. doi: 10.1016/j.jmb.2006.12.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van der Horst MA, van Stokkum IH, Crielaard W, Hellingwerf KJ. The role of the N-terminal domain of photoactive yellow protein in the transient partial unfolding during signalling state formation. FEBS Lett. 2001;497:26–30. doi: 10.1016/s0014-5793(01)02427-9. [DOI] [PubMed] [Google Scholar]
- Wadhams GH, Armitage JP. Making sense of it all: bacterial chemotaxis. Nat Rev Mol Cell Biol. 2004;5:1024–1037. doi: 10.1038/nrm1524. [DOI] [PubMed] [Google Scholar]
- Watts KJ, Ma Q, Johnson MS, Taylor BL. Interactions between the PAS and HAMP domains of the Escherichia coli aerotaxis receptor Aer. J Bacteriol. 2004;186:7440–7449. doi: 10.1128/JB.186.21.7440-7449.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watts KJ, Johnson MS, Taylor BL. Minimal requirements for oxygen sensing by the aerotaxis receptor Aer. Mol Microbiol. 2006a;59:1317–1326. doi: 10.1111/j.1365-2958.2005.05012.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watts KJ, Sommer K, Fry SL, Johnson MS, Taylor BL. Function of the N-terminal cap of the PAS domain in signaling by the aerotaxis receptor Aer. J Bacteriol. 2006b;188:2154–2162. doi: 10.1128/JB.188.6.2154-2162.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Webster CM, Xun L.Escherichia coli general NAD(P)H:flavin oxidoreductase is involved in aerotaxis. Abstr. I-036. 2003103rd meeting of The American Society for MicrobiologyWashington, D.C. [Google Scholar]
- Weis RM, Hirai T, Chalah A, Kessel M, Peters PJ, Subramaniam S. Electron microscopic analysis of membrane assemblies formed by the bacterial chemotaxis receptor Tsr. J Bacteriol. 2003;185:3636–3643. doi: 10.1128/JB.185.12.3636-3643.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams SB, Stewart V. Functional similarities among two-component sensors and methyl-accepting chemotaxis proteins suggest a role for linker region amphipathic helices in transmembrane signal transduction. Mol Microbiol. 1999;33:1093–1102. doi: 10.1046/j.1365-2958.1999.01562.x. [DOI] [PubMed] [Google Scholar]
- Woodmansee AN, Imlay JA. Reduced flavins promote oxidative DNA damage in non-respiring Escherichia coli by delivering electrons to intracellular free iron. J Biol Chem. 2002;277:34055–34066. doi: 10.1074/jbc.M203977200. [DOI] [PubMed] [Google Scholar]
- Zoltowski BD, Schwerdtfeger C, Widom J, Loros JJ, Bilwes AM, Dunlap JC, Crane BR. Conformational switching in the fungal light sensor Vivid. Science. 2007;316:1054–1057. doi: 10.1126/science.1137128. [DOI] [PMC free article] [PubMed] [Google Scholar]