

Abstract
Neocortex, a new and rapidly evolving brain structure in mammals, has a similar layered architecture in species over a wide range of brain sizes. Larger brains require longer fibers to communicate between distant cortical areas; the volume of the white matter that contains long axons increases disproportionally faster than the volume of the gray matter that contains cell bodies, dendrites, and axons for local information processing, according to a power law. The theoretical analysis presented here shows how this remarkable anatomical regularity might arise naturally as a consequence of the local uniformity of the cortex and the requirement for compact arrangement of long axonal fibers. The predicted power law with an exponent of 4/3 minus a small correction for the thickness of the cortex accurately accounts for empirical data spanning several orders of magnitude in brain sizes for various mammalian species, including human and nonhuman primates.
1. Introduction
The neocortex has expanded greatly during evolution of mammalian brains (1, 2). In insectivores, such as hedgehogs, for instance, the combined volume of neocortical gray matter and the adjacent white matter occupies no more than 10% to 20% of the total brain volume, whereas in humans it reaches 80% (3).
As illustrated schematically in Fig. 1, as brain size increases, the volume of the white matter beneath the cortex tends to increase faster than the volume of the cortical gray matter itself. Whereas the gray matter consists of neuronal cell bodies, their dendrites, and local ramifications of axons, plus glial cells and blood vessels, the white matter consists mostly of bundles of axons running a long distance. It is estimated that the great majority of white matter fibers connect different cortical regions rather than connect the cortex and subcortical structures (p. 106 in ref. 4 and p. 35 in ref. 5), although quantitative and comparative data are not available.
Figure 1.

Schematic section diagrams showing that the cerebral cortex of a larger brain tends to have disproportionally more long-distance connection fibers or white matter (dark regions) than the gray matter (folded outer surface). Adapted from a drawing by E. de Vries in ref. 7, reoriented and rescaled approximately by using the cat and puma brains at the University of Wisconsin website www.neurophys.wisc.edu/brain.
Allman (1) recently described an interesting power law relation between cortical gray matter volume and white matter volume in primates, based on an analysis of the anatomical data collected by Stephan and colleagues. The observed exponent was close to 4/3, suggesting that some simple geometric factors may be responsible for this regularity (1). This observation initiated our inquiry.
Brain scaling issues were studied by Dubois and his contemporaries (6). Danilewsky first demonstrated that larger brains contain relatively more white matter, and de Vries argued that the white matter should increase as the cube, and the gray matter as the square, which would imply a power law with the exponent 3/2 (7). Schlenska (8) presented quantitative data from different species and demonstrated power law relation between the volumes of gray and white matters with an exponent around 1.22 (inferred by using Eqs. 16 below). Frahm et al. (9) found similar results in a new data set, with an inferred exponent around 1.24. Hofman (10, 11) proposed different empirical scaling laws for different brain sizes, distinguishing the smooth (lissencephalic) brains from the convoluted (gyrencephalic) ones. He also included cortical thickness in the scaling and arrived at a 4/3 power law. Prothero and Sundsten (12) proposed explicit cubic geometric models for the cortical folding and generated various scaling laws numerically.
The purpose of this paper is to present an analytical theory for the gross organization of cortical gray matter and the associated white matter. The predicted relationships among anatomical variables, confirmed partially by the existing anatomical data, can account for the empirical scaling law and may serve as the basis for new avenues of investigation.
2. Empirical Scaling Law
As illustrated in Fig. 2 with 59 mammalian species, a single power law can adequately describe the relationship between the volume G of neocortical gray matter and the volume W of the adjacent white matter (including the corpus callosum but excluding the internal capsule), even though the neocortex itself may scale differently with the rest of brains for different species (9). Here G and W span more than five and six orders of magnitude, respectively, whereas their ratio W/G ranges over only one order of magnitude from about 0.06–0.07 to about 0.7–0.8.
Figure 2.

Cortical white and gray matter volumes of various species (n = 59) are related by a power law that spans five to six orders of magnitude. Most data points are based on measurement of a single adult animal. The line is the least squares fit, with a slope around 1.23 ± 0.01 (mean ± SD). The average and median deviations of the white matter volumes from the regression line are, respectively, 18% and 13% on a linear scale. Sources of data: If the same species appeared in more than one source below, the one mentioned earlier was used. All 38 species in table 2 in ref. 3 were taken, including 23 primates, 2 tree shrews, and 13 insectivores. Another 11 species were taken from table 2 in ref. 8, including 3 primates, 2 carnivores, 4 ungulates, and 2 rodents. Five additional species came from table 1 in ref. 11, including 1 elephant and 4 cetaceans. The data point for the mouse (G = 112 mm3 and W = 13 mm3) was based on ref. 30, and that for the rat (G = 425 mm3 and W = 59 mm3) was measured from the serial sections in a stereotaxic atlas (42). The estimates for the fisherman bat (Noctilio leporinus, G = 329 mm3 and W = 43 mm3) and the flying fox (Pteropus lylei, G = 2,083 mm3 and W = 341 mm3) were based on refs. 43 and 44, with the ratios of white and gray matters estimated roughly from the section photographs in the papers. The sea lion data (Zalophus californianus, G = 113,200 mm3 and W = 56,100 mm3) were measured from the serial sections at the website given in the legend to Fig. 1, with shrinkage correction.
The slope of the best fit line in Fig. 2 is 1.23 ± 0.01 by three different methods, including the standard least squares, least squares of perpendicular distance, and least absolute deviations, the third of which is more robust against outliers (13). The standard deviations for the slope and intercept were estimated directly for the first method and by bootstrap for the last two methods (14). Bootstrap may help detect outliers in the data because, when they are left out from a same-size resample, the correlation coefficient often increases, which could be exploited to improve estimation. Systematic bias caused by outliers was not detected in Fig. 2.
3. Theory of Scaling
Our analysis rests on two assumptions. First, we assume that each small piece of cortex of unit area, regardless of its thickness and the overall brain size, sends and receives about the same total cross-sectional area of long-distance connection fibers to and from other cortical regions. Second, we assume that the global geometry of the cortex minimizes the average length of the long-distance fibers.
The second assumption follows from Ramon y Cajal's principle for conservation of space, conduction time, and cellular materials (Chap. V in ref. 15). This principle has been explored more recently as the principle of minimal axon length (16–18). Consistent with previous observations on the basic uniformity of the cortex (19–21), the first assumption is supported loosely by the evidence that the total number of neurons beneath a unit cortical surface area is about 105/mm2 across different cortical regions for several species, from mouse to human (22) (after shrinkage correction). But there are exceptions, including the higher density in striate cortex of primates (22, 23), the lower density in dolphin cortex (24), and the variability observed in cat cortex (25). The number of axons leaving or entering the gray–white boundary per unit cortical area should be comparable to the number of cells, because the majority of cortical neurons are pyramidal cells and because most of them send axons into white matter, whereas most nonpyramidal cells do not (26). The numbers of efferent and afferent corticocortical fibers should roughly match up. Another piece of evidence consistent with the first assumption is that the cross-sectional area of corpus callosum, the collection of fibers connecting the two hemispheres, is proportional to the total cortical surface area for different species (21, 27). We emphasize that the uniformity assumption involves only the total cross-sectional area of white matter fibers, without directly specifying other factors, such as the total numbers of neurons and axons per unit area, the percentage of neurons that project into white matter, the thickness of axons, their branching, and the relative proportion of efferent and afferent fibers.
Our goal is to show that these two assumptions and constraints from geometry yield the empirical power law. The total volume of gray matter (G) is given by
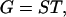 |
1 |
where S is the total area of the cortical surface, which can be convoluted, and T is the average thickness of the cortex. Because the thickness of a local cortical region often varies along the convolution (28), this relation is only a global average and is sometimes used to define the average thickness.
The volume of the white matter (W) that connects different cortical areas can be expressed as
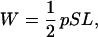 |
2 |
where p is the fraction of the surface area occupied by the cross section of the axonal fibers entering or leaving the white matter and L is the average length of the white matter fibers. To derive this expression, imagine that the white matter is divided conceptually into bundles of axonal fibers of the same cross-sectional area a. Area a is fixed but can be arbitrarily chosen, as long as the fibers within the same bundle are of the same length. Axons are allowed to run in different directions within the same imaginary bundle; occasional branching is allowed as long as the cross section of the bundle is conserved. Suppose there are a total of n such bundles and let Li be the length of the ith bundle, then the total volume of the white matter should be the sum of the volumes of all these bundles:
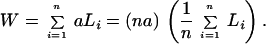 |
3 |
The second step is an identity, with 1/n ∑i=1n Li = L the average length of the fibers in white matter, and na the total cross-sectional area of these fibers. Because each fiber bundle connects two distinct cortical areas, imagine that every bundle is painted red at one end and green at the other end. Consider all of the red ends only, which should contact half of the total cortical surface area S/2. The total cross-sectional area at the red ends is still na. This area, by assumption, is related to the contacted half of cortex by the factor p, namely, na = pS/2, which leads to Eq. 2.
The uniformity assumption implies that the areal wiring fraction p should be constant across species. In addition, p < 1 is expected because axons leaving or entering the gray matter often run at an angle other than perpendicular to the cortical surface, so that their total cross-sectional area must be less than the cortical surface area.
The key question is how the gray matter volume is related to the white matter volume. Because they share the same surface area S, up to a constant fraction p, the real unknown quantity is the average fiber length L in white matter, which should be somehow related to the volume of the gray matter. We postulate that
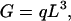 |
4 |
where q is a dimensionless constant, i.e., a pure number independent of the unit of length. In Section 5, we justify this postulate by a theoretical argument and support it with anatomical evidence.
The scaling law between gray and white matters now follows. Eliminating S from Eqs. 1 and 2 gives W = ½ pGL/T, and then substituting L in terms of G by using Eq. 4 yields
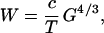 |
5 |
where
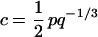 |
6 |
is a dimensionless constant. If the cortical thickness T is fixed, the scaling law predicts that the white matter volume increases exactly as the 4/3 power of the gray matter volume. In reality, the thickness increases slowly with the gray matter volume, so that the effective exponent should be slightly smaller than 4/3.
4. Cortical Thickness
The slow thickening of cortex for larger brains can be fitted by power laws against various variables (8, 12, 19, 28, 29). Consistent with previous results, direct power law regression of cortical thickness against gray matter volume yielded T ∼ G0.10±0.02, with a moderate correlation coefficient r = 0.81 in log–log space, based on 22 species from the data in table 1 in ref. 11. The five smallest brains with the inferred cortical thickness all smaller than 0.7 mm were left aside because they were too thin to fit comfortably into the same power law. For comparison, the average thickness of mouse cortex is 0.8–0.9 mm (30). Combining this result with Eq. 5, one expects that the white matter volume should scale as
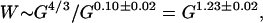 |
7 |
which naturally accounts for the empirical power law with the exponent 1.23 ± 0.01 in Fig. 2.
Power law scaling for cortical thickness may be justified theoretically (19). Hofman (11) proposed that a logarithm function would fit the thickness data for the smallest brains better than a power law, but his analysis included inferred thickness estimates, not all from direct measurement. For our current purposes, the thickening of cortex with brain size is treated simply as a small but noisy empirical correction to the dominant 4/3 exponent. From the perspective of dimensional consistency (31), neither a power law with a small exponent nor a logarithm function by itself has consistent dimensional units for variables from the same brain, although they are useful as numerical approximations.
5. The Cubic Postulate: Theory and Evidence
In the theoretical treatment above, the cubic relation between the gray matter volume G and the average length L of white matter fibers was taken as a postulate. This possibility is dimensionally consistent, but alternative relations also have consistent units, for instance, G = qL2T and G = qL1/2T5/2, where T is the cortical thickness. Here, we prove that the cubic relation allows the shortest average fiber length among other possible relations of the general product form
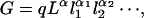 |
8 |
where q is a dimensionless constant and where l1, l2, … are additional length variables that might describe, for example, the thickness, the folding or gyri of the cortex, and so on, and are assumed to be smaller than the average fiber length L, namely, li < L; all of the exponents are assumed to be nonnegative: α ≥ 0, αi ≥ 0, and should sum up to the correct dimension:
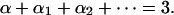 |
9 |
Under these assumptions, we have
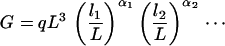 |
10 |
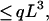 |
11 |
where the first equality follows directly from Eqs. 8 and 9 and where the inequality holds because each αi ≥ 0 and li/L < 1. Thus, we always have G ≤ qL3, regardless of the exact values of the auxiliary length variables and the choice of all of the exponents. Given a fixed G, the minimum length of L is achieved when the equality holds, which requires α = 3 and α1 = α2 = ⋯ = 0, so that all other variables disappear except for a cubic term of L, which is the original postulate.
Therefore, when the average fiber length has the longest length scale, the cubic relation should be optimal in reducing the length of white matter fibers. Indeed, when the total exponent of 3 is shared by shorter lengths instead of being occupied exclusively by the average fiber length L, then L must be longer to reach the same gray matter volume G. Although the constant q was fixed when comparing different possible products, this condition can be relaxed by allowing q to vary within fixed bounds. An even more general formulation is as follows. The most general relation between the volume G and the length variables L, l1, l2, … can be written as
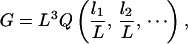 |
12 |
where Q is an arbitrary function whose value is independent of the unit used for length measurement. This result may be regarded as a consequence of Buckingham's π-theorem in dimensional analysis (32).
A necessary and sufficient condition for the optimality of the cubic relation is that the function Q is bounded, namely, Q ≤ q for some fixed q, and the equality can be achieved within the allowed ranges of variables and other hidden parameters (such as the exponents αi in the product case). Once we obtain G = L3Q ≤ qL3, the previous optimality argument following Eq. 11 applies without change.
How can the cubic scaling be tested, given that the average axonal length L in white matter is typically unknown? We may infer its value indirectly from Eq. 2, namely,
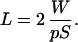 |
13 |
This formula provides new information because the value of the surface area S has not been explicitly used above (33, 34). The value of the constant p for areal wiring fraction has no effect on the exponent of the power law and will be estimated in Section 8.
As shown in Fig. 3, the data are noisy but support the cubic relation within the margin of error. Part of the noise may have arisen because the volume and surface data were compiled from different sources and different individual animals (11). Several smaller brains were not used here because the inferred values of L seemed too small to fit into the same power law. However, for the mouse brain, which is also quite small, an estimate of fiber length in white matter gives the average L ≈ 3 mm (figure 62 in ref. 30). Together with G = 112 mm3, this data point would be close to the regression line in Fig. 3. Therefore, more experimental data would be required to test whether the cubic relation may fail for the smallest brains.
Figure 3.

Testing the cubic postulate between the gray matter volume and the average length of fibers in white matter, which was inferred from Eq. 13 assuming p = 0.08. The theoretical slope of 3 (dashed line) is reasonably close to the slope of 3.3 estimated by least squares as shown, or the slope of 3.2 estimated by robust estimation with absolute deviations (not shown). Data were taken from table 1 in ref. 11, where cortical surface area and thickness are related rigorously by Eq. 1.
6. Arbitrary Dimensionless Product
According to the theory (Eq. 5), the product WG−4/3T = c should be a dimensionless constant across species, independent of the choice of length unit. An arbitrary product WαGβTγ is dimensionless whenever 3α + 3β + γ = 0. Can such products that are constant across species be discovered directly from the data?
Given a dimensionless number, its square or cube or any power function is also dimensionless, although larger powers tend to amplify the noise in the data. To compare with the theoretical formula, we set α = 1, leaving only one degree of freedom in choosing a dimensionless product. So we need only consider
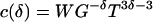 |
14 |
with an arbitrary δ. Eq. 5 corresponds to the case δ = 4/3.
To characterize the constancy of c(δ) across species for each given δ, we use the ratio of the standard deviation over the mean, because both quantities may vary over several orders of magnitude as δ changes. In Fig. 4, the best constancy achieved for δ = 1.36 ± 0.06 agrees with the theoretical value 4/3 ≈ 1.33. Therefore, the dimensionless product predicted by the theory, although still quite variable across species, is about the most constant among arbitrary dimensionless products.
Figure 4.

The coefficient of variation (standard deviation/mean) of the dimensionless number c(δ) plotted as a function of the test exponent δ in Eq. 14. The theoretical optimal value 4/3, as indicated by the dashed line, is close to the empirical minimum (•) reached at δ = 1.36 (±0.06, SD by bootstrap). Data were from table 1 in ref. 11 (27 species).
7. Limitation of Power Law Scaling
A problem arises when comparing Fig. 2 with Fig. 5 (8, 9), where the total volume
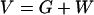 |
15 |
is defined as the sum of gray and white matter volumes. Because we already have the power law W ∼ Gα with α > 1 (Fig. 2), it is mathematically impossible to accommodate two additional power laws G ∼ Vβ and W ∼ Vγ without violating Eq. 15. An inescapable conclusion is that at least two of the power laws, and possibly all three of them, must be approximate, even in theory. Suppose one of them is genuine and known, then how can the other two approximate power laws be justified? (We would choose W ∼ Gα as genuine, given our theory. Consistently, the power law G ∼ Vβ in Fig. 5 is unlikely to be exact, despite its high correlation coefficient, because of the small but systematic deviations.) Given any one exponent, the other two exponents can be determined as a linear approximation by solving the following equations:
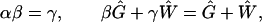 |
16 |
where Ĝ = exp(〈ln G〉) and Ŵ = exp(〈ln W〉) are averages performed in log space. For the data in Figs. 1 and 5, we have Ĝ = 5200 mm3 and Ŵ = 1248 mm3, and the three exponents determined directly by least squares are α = 1.2284, β = 0.9549, and γ = 1.1739. Given α, for instance, both β and γ can be predicted within 0.3% of error by Eqs. 16.
Figure 5.

The same data as in Fig. 1 plotted against the total volume of gray matter and white matter, showing the approximate nature of power laws for additive components. Of the three power laws shown here and in Fig. 1, at most one can be assumed to be rigorously true in theory without logical inconsistency.
The effect of adding power laws has been studied in allometry (35) and brain scaling (C. F. Stevens, personal communication). Brain volume has many components, such as cell bodies, dendrites, axons, blood vessels, and glia cells. Unless all components scale with the same exponent, it is logically inconsistent to assume rigorous power law relations among different components and their sums. Although power laws are often excellent approximations, a theory for power laws should be expected only to reveal the leading contributing factors. Additional information may be obtained by examining the relationships between the residuals from the fits (36).
8. Determining the Constants
The theory presented in this paper contains a total of three free parameters, namely, p, q, and c, all of which are dimensionless constants. Their values are not constrained by the theory and have to be estimated separately. Because they are related by Eq. 6, only two are independent.
The constant p is the fraction of the cortical surface area that is occupied by the total cross section of the fibers in white matter
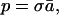 |
17 |
where ā is the average cross-sectional area of a single axonal fiber either entering or leaving the white matter, and σ is the total number of these axons per a unit surface area of the cortex. The diameters of axons, which may vary over an order of magnitude, have an average value of roughly 1 μm in corpus callosum (37, 38). A rough estimate ā ≈ 0.8 μm2 for the average cross-sectional area of a single fiber, corresponding to a diameter about 1 μm, is loosely compatible with the distributions shown in ref. 20. Sholl (26) estimated that the density of the axons crossing the gray–white boundary in cat cortex is about σ = 105/mm2, the same order of magnitude as neuron density per unit area (22, 28, 30). Taken together, we obtain a crude estimate p = σā ≈ 0.08.
Another independent estimate of p is possible by using Greilich's estimate of the distance of corticocortical connections in mouse (30). From the histogram in figure 62 of ref. 30, we estimate that in mouse the average axon length in white matter is around L = 3 mm. From Eqs. 1 and 2:
 |
18 |
where D ≡ W/S = WT/G can be interpreted as the average thickness of the layer of white matter, with S being the cortical surface area. Inserting the estimate of L and other known quantities (gray matter volume G = 112 mm3, white matter volume W = 13 mm3, cortical thickness T = 0.85 mm) into Eq. 18 yields p ≈ 0.07, close to the first estimate.
The constant q is the coefficient for the cubic relation between gray matter volume and average axonal length. Assuming p = 0.08 in Eq. 13 and by using Eq. 4, q can be estimated by averaging G/L3 over the same data as in Fig. 3. The result is q = 10 ± 5.
The constant c is the only free parameter in the 4/3 scaling law in Eq. 5. Averaging the product c = WG−4/3T across the same data in Fig. 4 yields c = 0.02 ± 0.01. Alternatively, Eq. 6 with the existing estimates p = 0.08 and q = 10 gives a consistent value c = ½pq−1/3 ≈ 0.02. Thus, all three constants can be estimated consistently with moderate accuracy from the existing data.
9. Discussion
The volumes of cortical gray matter and the adjacent white matter are closely related by a power law across nearly the full range of mammalian brain sizes (Fig. 2). The theoretical arguments in this paper show quantitatively how this relationship might follow as a necessary consequence of the basic uniformity of the neocortex and the compact wiring of its interconnection. The theory can account for the exponent of the empirical scaling law (Fig. 2 and Eq. 7) and allows all free parameters to be determined from the existing data (Section 8).
Intuitively, if each piece of cortex of a unit area is wired up by the same total cross-sectional area of long-distance fibers regardless of brain size, then these fibers would have to go longer distances to connect different parts of a larger brain, leading to a disproportionate expansion of the white matter. Although the exact relationship must depend on the exact pattern of connectivity and the exact manner of cortical folding, we have avoided explicitly modeling these details by invoking a scaling argument, according to which the gray matter volume should be proportional to the cube of the average fiber length in the white matter. The cubic relation is theoretically optimal in some general situations and appears to be valid empirically, at least for larger brains (Fig. 3). Taken together, as shown here the amount of white matter needed to interconnect the whole neocortex must follow a power law if cortical uniformity and compact wiring are assumed.
The theory here does not presume the uniformity of the cortex beyond the assumption that the total cross section of the axons passing the gray–white matter boundary should occupy a constant fraction of the cortical surface area across species. Although we have reviewed evidence that makes the assumption look plausible (Section 3), it remains to be determined to what extent the areal wiring fraction is actually constant, and how it compares with the neuron and axon densities per unit area, for different cortical regions of the same animal and across different species.
Additional anatomical measurements are needed. For example, it is uncertain exactly how many fibers in white matter interconnect the cortex and how many connect the cortex with various subcortical structures. We assumed here that the former is much more numerous. One crucial variable used in this paper is the average length of corticocortical fibers in the white matter, which has been inferred theoretically (Fig. 3). Measuring the fiber length in a wide range of species would provide valuable information, as for mouse cortex (30). Such information would be a key link between a global principle of minimal wiring, such as the cubic postulate above, and the geometry of the brain that actually embeds the hierarchical organization of the multiple cortical areas, as illustrated by the primate visual systems (39–41). The minimal wiring principle itself also needs to be assessed quantitatively for its range of validity. Noninvasive measurement with magnetic resonance imaging, by avoiding the shrinkage and distortion caused by fixation, staining, and sectioning, may yield useful geometric information about the cortex and its associated fibers, sampling across more living animals, and tracking the same individuals throughout development. These measurements combined with theoretical analysis should provide more accurate information on the basic constraints underlying brain development and evolution.
Acknowledgments
We thank J. M. Allman and C. F. Stevens for helpful discussions and H. J. Jerison, J. H. Kaas, S. LeVay, K. S. Rockland, and a reviewer for useful suggestions.
Footnotes
This paper was submitted directly (Track II) to the PNAS office.
Article published online before print: Proc. Natl. Acad. Sci. USA, 10.1073/pnas.090504197.
Article and publication date are at www.pnas.org/cgi/doi/10.1073/pnas.090504197
References
- 1.Allman J M. Evolving Brains. New York: Freeman; 1999. [Google Scholar]
- 2.Northcutt R G, Kaas J H. Trends Neurosci. 1995;18:373–379. doi: 10.1016/0166-2236(95)93932-n. [DOI] [PubMed] [Google Scholar]
- 3.Stephan H, Frahm H, Baron G. Folia Primatol (Basel) 1981;35:1–29. doi: 10.1159/000155963. [DOI] [PubMed] [Google Scholar]
- 4.Braitenberg V. On the Texture of Brains. New York: Springer; 1977. [Google Scholar]
- 5.Abeles M. Corticonics. Cambridge, U.K.: Cambridge Univ. Press; 1991. [Google Scholar]
- 6.Theunissen B. Eugène Dubois and the Ape-Man from Java. Dordrecht, the Netherlands: Kluwer; 1989. [Google Scholar]
- 7.Ariëns Kappers C U, Huber G C, Crosby E C. The Comparative Anatomy of the Nervous System of Vertebrates, Including Man. New York: Macmillan; 1936. [Google Scholar]
- 8.Schlenska G. J Hirnforsch. 1974;15:401–408. [Google Scholar]
- 9.Frahm H D, Stephan H, Stephan M. J Hirnforsch. 1982;23:375–389. [PubMed] [Google Scholar]
- 10.Hofman M A. Brain Behav Evol. 1985;27:28–40. doi: 10.1159/000118718. [DOI] [PubMed] [Google Scholar]
- 11.Hofman M A. Brain Behav Evol. 1988;32:17–26. doi: 10.1159/000116529. [DOI] [PubMed] [Google Scholar]
- 12.Prothero J W, Sundsten J W. Brain Behav Evol. 1984;24:152–167. doi: 10.1159/000121313. [DOI] [PubMed] [Google Scholar]
- 13.Press W H, Teukolsky S A, Vetterling W T, Flannery B P. Numerical Recipes in C. 2nd Ed. Cambridge, U.K.: Cambridge Univ. Press; 1992. [Google Scholar]
- 14.Efron B, Tibshirani R J. An Introduction to the Bootstrap. New York: Chapman & Hall; 1993. [Google Scholar]
- 15.Ramon y Cajal S. In: Histology of the Nervous System of Man and Vertebrates, trans. Swanson N, Swanson L W, editors. I. New York: Oxford Univ. Press; 1995. [Google Scholar]
- 16.Mitchison G. Trends Neurosci. 1992;15:122–126. doi: 10.1016/0166-2236(92)90352-9. [DOI] [PubMed] [Google Scholar]
- 17.Van Essen D C. Nature (London) 1997;385:313–318. doi: 10.1038/385313a0. [DOI] [PubMed] [Google Scholar]
- 18.Chklovskii D, Stevens C F. Advances in Neural Information Processing Systems 12. Cambridge, MA: MIT Press; 2000. , in press. [Google Scholar]
- 19.Stevens C F. Neural Comput. 1989;1:473–479. [Google Scholar]
- 20.Jerison H J. Brain Size and the Evolution of Mind. New York: American Museum of Natural History; 1991. [Google Scholar]
- 21.Ringo J L. Brain Behav Evol. 1991;38:1–6. doi: 10.1159/000114375. [DOI] [PubMed] [Google Scholar]
- 22.Rockel A J, Hiorns R W, Powell T P S. Brain. 1980;103:221–244. doi: 10.1093/brain/103.2.221. [DOI] [PubMed] [Google Scholar]
- 23.Peters A. In: Cerebral Cortex. Jones E G, Peters A, editors. Vol. 6. New York: Plenum; 1987. pp. 267–294. [Google Scholar]
- 24.Garey L J, Leuba G. J Comp Neurol. 1986;247:491–496. doi: 10.1002/cne.902470408. [DOI] [PubMed] [Google Scholar]
- 25.Beaulieu C, Colonnier M. J Comp Neurol. 1989;279:228–234. doi: 10.1002/cne.902790206. [DOI] [PubMed] [Google Scholar]
- 26.Sholl D A. The Organization of the Cerebral Cortex. New York: Wiley; 1956. [Google Scholar]
- 27.Bauchot R, Stephan H. Mammalia. 1961;25:314–341. [Google Scholar]
- 28.Bok S T. Histonomy of the Cerebral Cortex. Amsterdam: Elsevier; 1959. [Google Scholar]
- 29.Jerison H J. In: Primate Brain Evolution: Methods and Concepts. Armstrong E, Falk D, editors. New York: Plenum; 1982. pp. 77–84. [Google Scholar]
- 30.Braitenberg V, Schüz A. Cortex: Statistics and Geometry of Neuronal Connectivity. 2nd Ed. Berlin: Springer; 1998. [Google Scholar]
- 31.Jerison H J. Ann NY Acad Sci. 1977;299:146–160. doi: 10.1111/j.1749-6632.1977.tb41903.x. [DOI] [PubMed] [Google Scholar]
- 32.Langhaar H L. Dimensional Analysis and Theory of Models. New York: Wiley; 1951. [Google Scholar]
- 33.Elias H, Schwartz D. Science. 1969;166:111–113. doi: 10.1126/science.166.3901.111. [DOI] [PubMed] [Google Scholar]
- 34.Haug H. Am J Anat. 1987;180:126–142. doi: 10.1002/aja.1001800203. [DOI] [PubMed] [Google Scholar]
- 35.Reeve E C R, Huxley J S. In: Essays on Growth and Form. Le Gros Clark W E, Medawar P B, editors. Oxford: Clarendon; 1945. pp. 121–156. [Google Scholar]
- 36.Allman J M, McLaughlin T, Hakeem A. Proc Natl Acad Sci USA. 1993;90:3559–3563. doi: 10.1073/pnas.90.8.3559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Innocenti G M. In: Cerebral Cortex. Jones E G, Peters A, editors. Vol. 5. New York: Plenum; 1986. pp. 291–353. [Google Scholar]
- 38.LaMantia A-S, Rakic P. J Comp Neurol. 1990;291:520–537. doi: 10.1002/cne.902910404. [DOI] [PubMed] [Google Scholar]
- 39.Maunsell J H R, Van Essen D C. J Neurosci. 1983;3:2563–2586. doi: 10.1523/JNEUROSCI.03-12-02563.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Felleman D J, Van Essen D C. Cereb Cortex. 1991;1:1–47. doi: 10.1093/cercor/1.1.1-a. [DOI] [PubMed] [Google Scholar]
- 41.Rockland K S. In: Cerebral Cortex. Rockland K S, Kaas J H, Peters A, editors. Vol. 12. New York: Plenum; 1997. pp. 243–293. [Google Scholar]
- 42.Paxinos G, Watson C. The Rat Brain in Stereotaxic Coordinates. 2nd Ed. Sydney: Academic; 1986. [DOI] [PubMed] [Google Scholar]
- 43.Stephan H, Pirlot P. Zeitschrift für zoologische Systematik und Evolutionsforschung. 1970;19:200–236. [Google Scholar]
- 44.Stephan H, Pirlot P, Schneider R. Acta Anat. 1974;87:161–192. doi: 10.1159/000144169. [DOI] [PubMed] [Google Scholar]