

Abstract
Background/Aims
Circadian control of behaviour and physiology is a central characteristic of all living organisms. The master clock in mammals resides in the hypothalamus, where the suprachiasmatic nucleus (SCN) synchronises daily rhythms. A variety of recent evidence indicates that the neuropeptide vasoactive intestinal peptide (VIP) is critical for normal functioning of the SCN. The aim of our study was to examine the possible role of VIP in driving circadian rhythms in the hypothalamic-pituitary-adrenal axis.
Methods
Circulating ACTH and corticosterone concentrations were determined by round-the-clock sampling under diurnal and circadian conditions. The responsive aspects of the HPA axis were tested by application of acute stress by footshock and light.
Results
We demonstrate that the circadian rhythms in ACTH and corticosterone are lost in VIP-deficient mice. The ability of light to induce a corticosterone response was also compromised in the mutant mice, as was photic induction of Per1 in the adrenal glands. In contrast, the acute stress response was apparently unaltered by the loss of VIP.
Conclusion
Thus, our data demonstrates that VIP is essential for the circadian regulation of an otherwise intact hypothalamic-pituitary-adrenal axis.
Keywords: vasoactive intestinal peptide, circadian rhythms, glucocorticoids, HPA axis, stress, photic induction
Introduction
In mammals, the timing and occurrence of many physiological processes critically depends on the proper functioning of the suprachiasmatic nucleus (SCN). Current thinking places the SCN at the head of a hierarchy of oscillators distributed throughout a mammalian body [1,2]. The SCN plays interwoven dual roles, taking in light information from the environment to organise peripheral oscillators, as well as synchronising these oscillators even in the absence of external cues. Output signals from the SCN include neuronal signals and secreted factors that govern downstream rhythms in behaviour and endocrinology.
One of the downstream rhythmic signals is the daily rise in circulating corticosterone that occurs just prior to waking in mammalian species. It is postulated that this rise prepares an animal for the various stressors that are associated with activity onset, acting as a global signal for various processes like metabolism and the immune system. Glucocorticoid and mineralocorticoid receptors have a widespread distribution in the peripheral organs as well as throughout the central nervous system but are not found in the SCN [3], suggesting that the daily increase in corticosterone concentration could act as a unidirectional output signal from the SCN.
The circuitry by which the SCN regulates the HPA axis has been previously studied. Anatomical studies have provided evidence that the paraventricular nucleus (PVN) receives innervations from the SCN. Release of corticotrophin releasing factor (CRF) by neurons within the PVN is the critical step in stimulating ACTH release from the pituitary and thus the activation of the HPA axis [4]. In this manner, the SCN could provide circadian information to the HPA axis by direct innervation. Furthermore, SCN-lesioned rats show a loss of daily rhythm in ACTH and corticosterone [5-7]. Transplantation of foetal SCN tissue into SCN-lesioned hamsters does not restore the loss of glucocorticoid rhythmicity even though rhythms in voluntary wheel-running behaviour are restored [8]. These lines of evidence point towards the necessity of synaptic connections between the SCN and the PVN for circadian regulation of at least some endocrine rhythms. Of the SCN neurons that project to the PVN, a large number use the neuropeptide vasoactive intestinal peptide (VIP) as a signalling molecule [9,10]. Application of VIP into the hypothalamus, close to the PVN increases circulating ACTH and corticosterone concentration [11]. It is thus a potential regulator of the circadian drive of the HPA axis.
We have previously shown that mice lacking VIP have deficiencies in circadian behaviour, including poor locomotor rhythmicity under conditions of constant darkness and inappropriate responses to phase-shifting light pulses [12]. The receptor for VIP in the SCN is the VPAC2 receptor, mutants of which also have pronounced circadian defects [13,14]. Additionally, the SCN of VIP-deficient and VPAC2R mutant mice have fewer intrinsically rhythmic cells, and those that remain rhythmic show a lack of synchrony of electrical firing with each other [15,16]. This suggests the underlying cause of the circadian deficiencies to be within the SCN, due to both loss of intrinsic rhythmicity within each SCN neuron as well as defects in coupling of rhythms between SCN neurons. It also makes VIP-deficient mice a useful model in which to study the effects of an arrhythmic SCN on neuroendocrine output. Previous lesion studies have demonstrated the need for an intact SCN for neuroendocrine rhythms. However, electrolytic or physical cutting in the hypothalamus might damage connections with neighbouring nuclei. In the case of VIP-deficient mice, the SCN are still anatomically intact, albeit without the developmental and functional expression of VIP. We can thus determine the necessity of SCN rhythmicity on the circadian responses in the periphery. To this end, we designed our study to test the hypothesis that VIP plays a critical role in conveying time of day information within the HPA axis.
Materials and Methods
Experimental animals
Our studies to measure diurnal and circadian circulating glucocorticoid concentrations and the effect of adrenalectomy on glucocorticoid concentrations utilised adult male offspring of F1 C57BL/6J × 129/Sv mice lacking the gene encoding for VIP and PHI: Vip−/−. Wild-type controls from the same litter were analysed when available, but controls from other age-matched litters were used when necessary. Response to acute stress and light treatment was measured in male Vip−/− mice on a C57BL/6J background (backcrossed for 12 generations) with age-matched controls from inbred C57BL/6J lines. In all studies, the recommendations for animal use and welfare, as dictated by the UCLA Division of Laboratory Animals and the guidelines from the National Institutes of Health, were followed.
Behavioural analysis
Male mice of at least 10 weeks of age were housed individually and their wheel-running activity was recorded as revolutions per 3 minute interval. The running wheels and data acquisition system were obtained from Mini Mitter Co. (Bend, OR). The mice were exposed to a 12:12 light-dark cycle (LD) for 2−3 weeks (light intensity 36μW/cm2 ≅ 120 lux). The mice were then placed into constant darkness (DD) to assess their free-running activity pattern. Circadian time (CT) was determined by activity records with activity onset denoted as CT12. Sample collections and treatments in DD were typically performed after 2−4 days in constant darkness. In the dark portion of LD conditions, as well as under DD conditions, handling of mice was carried out with the aid of an infrared viewer (FJW Industries, Ohio).
Adrenalectomy
Bilateral adrenalectomies were performed through dorsal incisions under anaesthesia by ketamine/xylazine. Sham operations were performed in a similar fashion with the exception that the adrenal glands were left intact. Following surgery, adrenalectomised (ADX) mice received a 0.9% NaCl solution to drink. Blood was collected by terminal cardiac puncture under isoflurane anaesthesia in late day (ZT10) two weeks after adrenalectomy.
Induction of stress in mice
Acute stress was applied in early night at Zeitgeber Time (ZT) 15, 3 hours after lights-off, in the form of an underfoot stimulus (0.5 mA, footshock) as previously described [17]. Following the footshock, mice were returned to their home cages and anaesthetised with isoflurane 20 minutes after initial introduction to the stress protocol. Trunk blood was collected after decapitation for glucocorticoid assays. In a separate experiment to determine induction of gene expression in the adrenal glands, following footshock, mice were returned to their home cages and anaesthetised with isoflurane 60 minutes after initiation of the stress protocol. Adrenal glands were dissected following decapitation and immediately homogenised in Trizol.
Acute light treatment
A sub-maximal (15μW/cm2, approx. 50 lux) light treatment was applied in early night (ZT16) for 30 minutes, after which the mice were returned to dark conditions for an additional 30 minutes. Trunk blood was sampled after anaesthesia with isoflurane. The same light treatment was applied to free-running mice in early night (CT16) in a separate experiment to determine induction of gene expression in the adrenal glands. 60 min after the initiation of the light treatment, mice were anaesthetised, decapitated and the adrenal glands dissected and homogenised in Trizol.
Glucocorticoid assays
To obtain serum, blood samples were allowed to clot at room temperature for 30 minutes prior to centrifugation at 1000 × g. The supernatant was stored at −20°C until assayed. Serum ACTH concentration was determined using a commercially available competitive enzyme immunoassay (ACTH 1−39 EIA, Peninsula Laboratories, California). The intra-assay coefficient of variation (CV) was <5%, the inter-assay CV was <14% and the sensitivity was 0.04−0.06 ng/ml. Serum samples were typically diluted between 2-fold and 10-fold prior to assay. Serum corticosterone concentration for the round-the-clock and adrenalectomy experiments were measured by competitive enzyme immunoassay (R&D Systems, Minneapolis). Intra-assay precision was <8.4%, inter-assay CV was <13.1% and sensitivity was 27 pg/ml. Samples were typically diluted between 8-fold and 150-fold prior to assay. Due to discontinuation of the R&D corticosterone EIA kit, serum corticosterone concentration for the acute stress and light treatment experiments was measured using a different competitive enzyme immunoassay (Correlate-EIA Corticosterone kit, Assay Designs, Michigan). The intra-assay CV was <8%, the inter-assay CV was <13.1% and the sensitivity was 27 pg/ml. Serum samples were diluted 100-fold prior to assay.
Quantitative real-time PCR
RNA was extracted from adrenal glands using the Trizol (Invitrogen) procedure and treated with DNase (Turbo DNA-free, Ambion) for 30 min to remove possible DNA contaminants. cDNA was obtained by reverse-transcription of 1μg total RNA (High Capacity cDNA Reverse Transcription Kit, Applied Biosystems) and no-RT controls were run routinely. Primers for Per1 and Hprt were designed using Oligo6 (Molecular Biology Insights) and Mfold [18] programs to cross intron-exon boundaries and hence further prevent contaminants in the subsequent RT-PCR. Oligonucleotide primer sequences used were Per1-sense: 5’-TCC TCC TCC TAC ACT GCC TCT-3’ and Per1-antisense: 5’-TTG CTG ACG ACG GAT CTT T-3’; and Hprt-sense: 5’-TGG TGA AAA GGA CCT CTC GAA-3’ and Hprt-anti-sense: 5’-TCA AGG GCA TAT CCA ACA ACA-3’. PCR products from each primer pair were cloned and sequenced to confirm specificity as well as run on an agarose gel to ensure no primer dimers were formed. Quantitative PCR using SyBr Green (SYBR Green PCR Master Mix, Applied Biosystems) was performed using the equivalent of 50ng of starting total RNA (1:20 of the initial cDNA reaction from 1μg of total RNA) in a 25μl reaction comprising of the 2x SYBR Green reaction mix and 1.2μM each of the forward and reverse primers. The efficiency of each primer pair was confirmed prior to experimental use by serial dilutions of a cDNA sample, and by plotting Ct (cycle number at which threshold within the linear range was reached) values against log(total RNA). Melting curves were determined at the end of each reaction to check product specificity. The relative levels of Per1 transcripts were determined using the 2−ΔΔCt method, using Hprt as the normalising reference gene.
Statistical measurements
Between-group differences were generally determined by t-tests or Mann-Whitney rank sum tests when appropriate. Values were considered significantly different if P<0.05. All tests were performed using GraphPad Prism or SigmaStat. In the text, values are shown as mean ± SEM.
Results
Deficits in diurnal rhythms of glucocorticoids in Vip−/− mice
We first determined concentration of corticosterone in serum across a 24-hour period in normal lighting conditions of 12:12 hours LD (Figure 1A). Blood was sampled from wild-type (WT) and Vip−/− (VIP KO) mice every 3 hours (±1h to accommodate multiple sampling), starting at 2 hours after lights-on (ZT2). We next determined circulating corticosterone concentration for both WT and VIP KO mice every 4 hours in DD at CT points determined by locomotor activity (Figure 1B). As CT is calculated based on voluntary wheel running activity and VIP KO mice have varied locomotor activity patterns [12], only behaviourally rhythmic mice were used. We found that WT mice exhibit a strong rhythm in circulating corticosterone in both LD and DD (P<0.001 and P<0.05 by ANOVA), with a daily rise in either late day or late subjective day respectively. In contrast, VIP KO mice did not exhibit a daily rhythm under either condition.
Figure 1.

Concentration of corticosterone in serum over a 24 hour period in wild-type (WT) and Vip−/− (VIP KO) mice. Serum was sampled every 3 hours (n=3 per time point) under light-dark conditions (LD) and every 4 hours under constant darkness (DD). Circulating corticosterone concentration (ng/ml) was determined by ELISA.
A. Serum corticosterone concentration under 12:12 LD conditions plotted against zeitgeber time (ZT). The WT peak in corticosterone at ZT8 was 30-fold higher than the trough at ZT2 and cycling conditions for WT concentrations were determined to be significant (P<0.001) by ANOVA. In contrast, VIP KO mice displayed no such rhythmicity in basal concentration of corticosterone.
B. Corticosterone measurements plotted against circadian time (CT) in DD. Again, WT concentrations were determined to be significantly different by ANOVA (P<0.05) with a fourfold difference between peak and trough values at CT10 and CT2 respectively.
Further characterisation of the effect of loss of VIP on endocrine function was carried out by determining concentration of circulating ACTH over the course of a 24-hour day in LD (Figure 2A). Serum ACTH was measured for WT and VIP KO mice in the same manner as for corticosterone. The serum concentration of ACTH in DD was also measured (Figure 2B). We found that WT mice display a clear rhythm in ACTH under LD as well as DD (P<0.01). VIP KO mice, on the other hand, do not have rhythmic ACTH concentration. In addition, basal concentration of ACTH in VIP KO mice is considerably higher than in WT mice. This observation is consistent with a loss of the negative feedback regulation of ACTH by corticosterone.
Figure 2.

ACTH measurements from WT and VIP KO mice in LD plotted against ZT (A) and DD plotted against CT (B). Concentration of ACTH in the serum was determined by ELISA in 3 hour intervals in LD and 4 hour intervals in DD (n=2 or 3 for every time point). Circulating ACTH concentrations were found to be significantly rhythmic by ANOVA (P<0.01) for WT mice in LD and DD. The peak concentration of diurnal (LD) ACTH was determined to be at ZT11, and was 3-fold higher than at ZT2. Under DD, ACTH concentration peaked at CT12, where it was found to be 2-fold higher than CT4. In contrast, VIP KO mice did not display rhythmicity under either LD or DD, with circulating corticosterone concentrations remaining high throughout the 24-hour period.
Feedback regulation in the HPA axis of Vip−/− mice
One measure of corticosterone feedback in the HPA axis is that ACTH secretion increases in response to the removal of the adrenal glands. In this set of experiments, we sought to determine whether this negative feedback response is altered in VIP-deficient mice. As a control, we first confirmed that removing the adrenals (ADX) results in the sharp reduction of serum corticosterone concentration in both WT and VIP-deficient mice (WT sham 167.5±27.66 ng/ml, n=4; WT ADX 14.02±4.58 ng/ml, n=8; VIP KO sham 72.26±28.12 ng/ml, n=4; VIP KO ADX 10.28±2.94 ng/ml, n=7). WT mice exhibited a robust increase (P<0.01) in serum concentration of ACTH in response to the removal of the adrenal glands (WT sham 0.52±0.11 ng/ml, n=4; WT ADX 1.68±0.22 ng/ml, n=8). VIP-deficient mice started with high concentrations of ACTH (VIP KO sham 1.18±0.48 ng/ml, n=4), which remained high after adrenalectomy (VIP KO ADX 1.08±0.21 ng/ml, n=7). The similar serum concentration of ACTH pre- and post-adrenalectomy in the VIP KO mice suggests aberrations in the negative feedback of corticosterone on CRF or ACTH directly.
Acute stress corticosterone response
While the concentration of circulating corticosterone fluctuates with the 24-hr cycle, in a shorter time scale, the concentration of this hormone is actively regulated as part of the stress response exhibited by mice. We sought to determine whether the endocrine response to an acute stressor is altered in VIP-deficient mice. For these experiments, we examined corticosterone response to a mild footshock in WT and VIP KO mice in early night (ZT15). WT mice subjected to footshock exhibited significantly higher concentrations (P<0.01) of serum corticosterone than unstressed mice (Figure 3), and VIP KO mice showed a similar increase in corticosterone concentration upon exposure to acute stress (P<0.05). Serum corticosterone concentration upon acute stress was not significantly different between WT and VIP KO mice, suggesting the induction of corticosterone in response to stress is intact in VIP-deficient mice.
Figure 3.

Corticosterone response to acute stress by footshock in early night (ZT15). WT mice displayed a significant induction of corticosterone in response to acute stress (baseline: 33.63±16.07 ng/ml, n=5; vs. stress: 201.30±17.16 ng/ml, n=5; P<0.01). A similar corticosterone response to acute stress was also observed in VIP KO mice (baseline: 13.14±6.49 ng/ml, n=3; vs. stress 243.00±35.39 ng/ml, n=6; P<0.05). Statistical significances were determined by non-parametric Mann-Whitney t-tests.
Light-induced corticosterone production
Light treatment during early night has been shown to increase circulating corticosterone concentration in an ACTH-independent manner [19]. To test the role of VIP in the photic induction of corticosterone, we exposed WT and VIP KO mice to light (30 min duration) in early night (ZT16) and sampled serum 60 min after initial exposure to light. We observed a significant increase in corticosterone concentration in WT mice following the light pulse (P<0.05; Figure 4), confirming the previous study. In contrast, VIP KO mice failed to exhibit a significant induction of corticosterone in response to light, thus demonstrating a deficit in the light-responsive aspect of the HPA axis in VIP-deficient mice.
Figure 4.

Corticosterone induction by exposure to light in early night (ZT16). Serum corticosterone from untreated mice (n=4) and mice subjected to a light treatment (15μW/cm2, 30 minute duration, n=4) were determined by ELISA. WT mice respond to the light with a significance increase in circulating corticosterone concentration (283.60±28.84 ng/ml; P<0.05), but VIP KO mice failed to show a significant corticosterone response (33.42±17.79 ng/ml).
Measurement of Per1 expression in the adrenal glands following light treatment and acute stress
To further study the effect of the loss of VIP on photic induction in the periphery, we exposed WT and VIP KO to a 30 min light treatment in early night (CT16) and determined Per1 expression in the adrenal glands 60 min after initiation of the light treatment. A significant increase in Per1 expression levels was observed in the adrenal glands of WT mice 60 min after the initiation of the light pulse (P<0.001; Figure 5). In contrast, measurement of Per1 expression following light treatment was not significantly higher in VIP KO mice. Acute stress evoked by mild footshock in early night (ZT15) also increased Per1 expression in adrenal glands. Per1 expression was increased 2.7-fold in WT mice (baseline: 6.16±0.44, n=3; vs. stress 16.65±1.23, n=3; P<0.005). VIP KO also displayed a significant, albeit blunted, 1.7-fold increase (baseline: 4.67±0.60, n=3; vs. stress 7.94±0.96, n=3; P<0.05) in Per1 expression. These gene expression results corroborate our finding that the loss of VIP does not affect some aspects of the HPA axis, but is critical for response to light.
Figure 5.
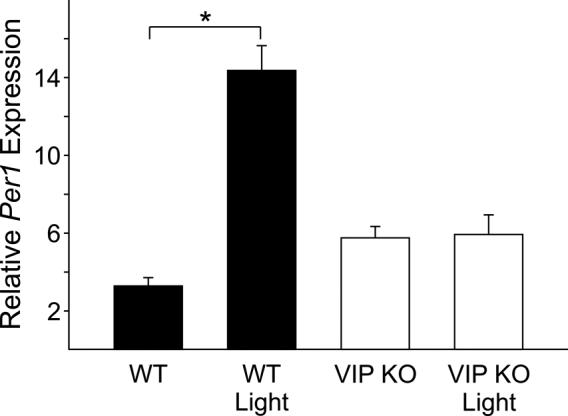
Photic induction of Per1 in the adrenal glands. 60 min after the start of the light treatment in early night (CT16; 15μW/cm2, 30 minute duration), adrenal glands were removed for RNA analysis by quantitative RT-PCR. Expression levels of Per1 were normalised to Hprt using the 2−ΔΔCt method. WT mice showed a significant increase in relative Per1 expression (baseline: 3.29±0.43, n=4; vs. light pulse 14.37±1.27, n=4; P<0.001). VIP KO mice did not show a significant increase in Per1 expression (baseline: 5.76±0.59, n=4; vs. light pulse 5.93±1.01, n=4).
Discussion
The secretion of glucocorticoid hormones is a highly regulated process. One of the important regulators of the HPA axis is circadian information originating in the SCN. This temporal information is critical for coordination of whole body physiology, preparing the body for daily changes between activity and rest. The SCN can be divided into two anatomically distinct portions based on the expression patterns of two neuropeptides: VIP and vasopressin (AVP). VIP-ir fibers make up the “core”, which receives input from the retinohypothalamic tract (RHT) and in all likelihood signals to the “shell”, made up of AVP-ir neurons. It is postulated that the AVP-neurons, situated next to the third ventricle, play a neurosecretory role. Both VIP-ir and AVP-ir fibres have been found to innervate various hypothalamic nuclei [20-23]. In particular, VIP-ir fibres in the PVN show a similar distribution to transcripts of the VPAC2 receptor [24]. This anatomical distribution of VIP and VPAC2R strongly suggests the importance of VIP in SCN to PVN communication, and may be critical for transmission of circadian signals to the HPA axis.
The necessity of VIP and its receptor in the SCN, VPAC2R, for rhythms in electrical activity in SCN neurons and behavioural rhythms has been previously demonstrated [12,14-16,25]. We now demonstrate that diurnal and circadian rhythms of serum concentration of ACTH and corticosterone are lost in VIP-deficient mice. As such, we propose that VIP, and likely its receptor VPAC2R, are critical for the daily rhythms of glucocorticoids by their action to maintain rhythmicity and synchrony within the SCN.
The VPAC2R knockout mice show a lack of rhythms in expression levels of clock genes [14] and glucocorticoid rhythmicity [26]. We would predict a similar loss in clock gene rhythmicity in VIP-deficient mice. Mutants in clock genes, e.g. the Period and Cryptochrome genes, show a variety of circadian defects. In particular, the Per2/Cry1 double knockout model shows a lack of glucocorticoid rhythms [27]. In this instance, the authors determined that a functional oscillator in the SCN is necessary for the rhythms in ACTH and that a circadian rhythm in corticosterone is dependent on the oscillator in adrenals. They suggest the adrenals can modulate production of corticosterone in a manner dependent on time of day. Their data and ours highlight the importance of a functional/rhythmic SCN in determining rhythmicity in the periphery. It remains to be seen if the VIP-deficient mice have functional peripheral oscillators and if such oscillators require a daily rhythm in glucocorticoids for synchrony with the SCN.
Other dimensions of VIP action in the HPA axis could be within the adrenals, where VIP has been shown to be synthesised in adrenal glands [28,29]. VIP-ir fibres innervate both the cortex and medulla of adrenal glands and originate from the splanchnic nerve or from ganglion cells in the medulla [30,31]. Both receptors for VIP, VPAC1R and VPAC2R, are also found in the adrenal medulla and capsule [32,33]. The role of VIP in the adrenal glands is not entirely clear; functional studies have demonstrated a response by VIP to ACTH administration and splanchnic nerve stimulation as well as effects of VIP on glucocorticoid secretion by the adrenal glands [28,34,35]. Splanchnic denervation in rats has been shown to influence the diurnal rhythm of corticosterone production, either by sympathetic signals to directly induce corticosterone production or in adapting the adrenal responsiveness to ACTH [36]. This suggests some form of interaction between the HPA axis and the autonomic nervous system controls corticosterone rhythmicity.
In our study, VIP-deficient mice maintain their capacity to produce corticosterone; basal concentrations, while not rhythmic, are similar to non-peak concentrations in WT mice, suggesting that VIP signalling is not critical for normal adrenal production of corticosterone. Alternatively, if VIP does play a role within the adrenals in interpretation of the ACTH or splanchnic nerve signal, other neuropeptides, like PACAP or AVP, may play a compensatory role in the mutant mice. While PACAP has been implicated in a number of stress responses (e.g. 37, 38), recent data makes it less likely that an up-regulation PACAP may compensate for the loss of VIP (39). The basal concentration of ACTH is significantly higher in the VIP-deficient mice at all time points tested. This raises the question of whether the loss of the ACTH rhythm is due to a breakdown in its circadian control or if VIP is necessary for the repression of ACTH from the pituitary. Adrenalectomy normally results in the loss of the major source of circulating corticosterone and a corresponding increase in ACTH. Prior to adrenalectomy, ACTH concentration in VIP mutants is already significantly higher than in WT mice, and remains high following adrenalectomy. The high basal concentration of ACTH may indicate defects in the ability of corticosterone to feedback on CRF-releasing neurons in the PVN.
One of the key components driven by the HPA axis is the rapid release of ACTH and adrenal corticosterone into the circulation in response to a variety of stressors (like noise, restraint, footshock, immune challenge, hypoglycaemia, haemorrhage, novel environment, predator exposure). This responsiveness of the HPA axis has a diurnal rhythm, and the magnitude of the corticosteroid response to stress is stressor specific. Stress by ether and novel environment has been reported to produce a greater corticosterone response at the start of the active phase [40], with restraint stress inducing a larger response in the early day [6, 41]. Lesioning the SCN causes a loss in the diurnal difference in response, but not the ability to respond [6]. Furthermore, application of anti-sense VIP transcripts by intracranial infusion into the hypothalamic region transiently blunts the corticosterone rhythm without affecting corticosterone production in response to acute stress [42]. In support of this, our study shows that mice lacking VIP not only in the hypothalamus but also in the adrenals are capable of mounting a normal corticosterone response to acute stress. Taken together with the ability of VIP-deficient mice to maintain normal basal concentration of corticosterone, we can conclude that VIP is not essential for non-circadian functioning of the HPA axis.
Light acts to entrain the circadian clock and synchronises it to the environment. The term circadian is literally translated to “about a day”, and the free-running period of nocturnal animals tends to be just under 24 hours. As such, under conditions of LD of 12:12 or normal changing LD patterns in the wild, light synchronises the circadian clock daily by shifting the phase of the oscillator [43]. The ability of light to shift the endogenous clock is well documented, and even brief pulses of light (from seconds to minutes) can effect a change in the rhythm of locomotor behaviour and shift peripheral molecular oscillators [e.g. 44]. The phase shifting effect of light has been shown to pass through the RHT to VIP neurons in the SCN [45], modifying the concentration of one of the clock genes, Per1 [46], and changing the phase of the molecular oscillator. The SCN is necessary for light-induced changes in the adrenals: in SCN-lesioned rodents, there was no corticosterone induction nor Per1 up regulation [19]. This adrenal response is also dependent on the integrity of the splanchnic nerve [19,47,48]. In response to exposure to light in the early night, we observed an increase in corticosterone concentration in WT mice, in agreement with evidence from Ishida and colleagues [19]. In contrast, VIP mutant mice are deficient in this light-induced response, suggesting that VIP is critical for interpretation of light input.
We also showed that we can measure a significant up regulation of Per1 expression following light treatment in WT mice. This response is lost in VIP KO mice, providing further evidence of a defect in response to light. The Per1 response to a different acute stressor, footshock, remains present, if blunted, in VIP KO mice, again confirming that some aspects of the HPA axis are unaffected by the loss of VIP. The selective deficits in behavioural, hormonal and gene expression response to light continue to be the most intriguing aspects of VIP loss. It remains to be determined if this deficiency is due to loss of VIP in the SCN or a breakdown of signalling in the rest of the HPA axis that may be dependent on VIP.
There are many peripheral organs in which rhythms in gene expression have been demonstrated, including the liver and adrenal glands [49-51]. For the SCN to synchronise the molecular oscillators in the periphery, there must be an entraining or resetting signal originating from the SCN. This could take a number of forms, including direct secretion from secretory AVP-ir neurons next to the third ventricle, neurotransmitters through the autonomic nervous system, or activation of the HPA axis [52]. There is a growing body of evidence for the HPA axis as an output of the SCN. A glucocorticoid receptor agonist, dexamethasone, can induce Per1 expression in the liver with subsequent phase shifting of the liver oscillator [3]. Further evidence comes from food-restriction studies, where loss of endogenous corticosterone by adrenalectomy has been found to speed up synchronisation of the liver oscillator to food-restriction regimes [53]. Indications of the possible roles corticosterone could play as an output signal were teased out in a study where the rhythmicity of genes involved in glycogenesis, gluconeogenesis and cholesterol synthesis, amongst other pathways, in the liver were affected by adrenalectomy [54]. Another striking effect of adrenalectomy on circadian rhythms in non-SCN tissues is the loss of PER2 rhythms in the oval nucleus of stria terminalis and the central nucleus of the amygdala in rats, which can be rescued by rhythmic application of corticosterone [55].
The VIP knockout mouse provides a unique model in which the circadian information from the SCN is lost or not transmitted. So far, we have determined that rhythms in glucocorticoids are lost even though the ability of the HPA axis to respond to acute stress is intact. It is reasonable to assume that other endocrine rhythms are lost as well. This unique dissociation of circadian and responsive aspects of the HPA axis thus makes VIP-deficient mice a good model for dissecting out downstream physiological processes that require signals from the SCN. Conversely, it may also prove a useful model in which to study the HPA axis without the confounding effect of circadian regulation. Furthermore, the deficient corticosterone response to light provides an interesting avenue to explore the mechanisms by which light signals from the environment change physiological state.
Acknowledgements
This research was supported by NIH grants NS-043169 to CSC and MH-65497 to JAW.
References
- 1.Hastings MH, Reddy AB, Maywood ES. A clockwork web: circadian timing in brain and periphery, in health and disease. Nat Rev Neurosci. 2003;4:649–661. doi: 10.1038/nrn1177. [DOI] [PubMed] [Google Scholar]
- 2.Schibler U, Ripperger J, Brown SA. Peripheral circadian oscillators in mammals: time and food. J Biol Rhythms. 2003;18:250–260. doi: 10.1177/0748730403018003007. [DOI] [PubMed] [Google Scholar]
- 3.Balsalobre A, Brown SA, Marcacci L, Tronche F, Kellendonk C, Reichardt HM, Schutz G, Schibler U. Resetting of circadian time peripheral tissues by glucocorticoid signaling. Science. 2000;289:2344–2347. doi: 10.1126/science.289.5488.2344. [DOI] [PubMed] [Google Scholar]
- 4.Swanson LW, Sawchenko PE, Rivier J, Vale WW. Organization of ovine corticotropin-releasing factor immunoreactive cells and fibers in the rat brain: an immunohistochemical study. Neuroendocrinology. 1983;36:165–186. doi: 10.1159/000123454. [DOI] [PubMed] [Google Scholar]
- 5.Moore RY, Eichler VB. Loss of a circadian adrenal corticosterone rhythm following suprachiasmatic lesions in rat. Brain Research. 1972;42:201–206. doi: 10.1016/0006-8993(72)90054-6. [DOI] [PubMed] [Google Scholar]
- 6.Sage D, Maurel D, Bosler O. Involvement of the suprachiasmatic nucleus in diurnal ACTH and corticosterone responsiveness to stress. Am J Physiol Endocrinol Metab. 2001;280:E260–E269. doi: 10.1152/ajpendo.2001.280.2.E260. [DOI] [PubMed] [Google Scholar]
- 7.Buijs RM, Kalsbeek A, van der Woude TP, van Heerikhuize JJ, Shinn S. Suprachiasmatic nucleus lesion increases corticosterone secretion. Am J Physiol. 1993;264:R1186–1192. doi: 10.1152/ajpregu.1993.264.6.R1186. [DOI] [PubMed] [Google Scholar]
- 8.Meyer-Bernstein EL, Jetton AE, Matsumoto SI, Markuns JF, Lehman MN, Bittman EL. Effects of suprachiasmatic transplants on circadian rhythms of neuroendocrine function in golden hamsters. Endocrinology. 1999;140:207–218. doi: 10.1210/endo.140.1.6428. [DOI] [PubMed] [Google Scholar]
- 9.Teclemariam-Mesbah R, Kalsbeek A, Pevet P, Buijs RM. Direct vasoactive intestinal polypeptide-containing projection from the suprachiasmatic nucleus to spinal projecting hypothalamic paraventricular neurons. Brain Res. 1997;748:71–76. doi: 10.1016/s0006-8993(96)01246-2. [DOI] [PubMed] [Google Scholar]
- 10.Buijs RM, Wortel J, van Heerikhuize JJ, Feenstra MGP, Ter Horst GJ, Romijn HJ, Kalsbeek A. Anatomical and functional demonstration of a multisynaptic suprachiasmatic nucleus adrenal (cortex) pathway. European Journal of Neuroscience. 1999;11:1535–1544. doi: 10.1046/j.1460-9568.1999.00575.x. [DOI] [PubMed] [Google Scholar]
- 11.Alexander LD, Sander LD. Vasoactive intestinal peptide stimulates ACTH and corticosterone release after injection into the PVN. Regulatory Peptides. 1994;51:221–227. doi: 10.1016/0167-0115(94)90068-x. [DOI] [PubMed] [Google Scholar]
- 12.Colwell CS, Michel S, Itri J, Rodriguez W, Tam J, Lelievre V, Hu Z, Liu X, Waschek JA. Disrupted circadian rhythms in VIP- and PHI-deficient mice. American Journal of Physiology-Regulatory Integrative and Comparative Physiology. 2003;285:R939–R949. doi: 10.1152/ajpregu.00200.2003. [DOI] [PubMed] [Google Scholar]
- 13.Reed HE, Meyer-Spasche A, Cutler DJ, Coen CW, Piggins HD. Vasoactive intestinal polypeptide (VIP) phase-shifts the rat suprachiasmatic nucleus clock in vitro. Eur J Neurosci. 2001;13:839–843. doi: 10.1046/j.0953-816x.2000.01437.x. [DOI] [PubMed] [Google Scholar]
- 14.Harmar AJ, Marston HM, Shen S, Spratt C, West KM, Sheward WJ, Morrison CF, Dorin JR, Piggins HD, Reubi JC, Kelly JS, Maywood ES, Hastings MH. The VPAC(2) receptor is essential for circadian function in the mouse suprachiasmatic nuclei. Cell. 2002;109:497–508. doi: 10.1016/s0092-8674(02)00736-5. [DOI] [PubMed] [Google Scholar]
- 15.Aton SJ, Colwell CS, Harmar AJ, Waschek J, Herzog ED. Vasoactive intestinal polypeptide mediates circadian rhythmicity and synchrony in mammalian clock neurons. Nat Neurosci. 2005;8:476–483. doi: 10.1038/nn1419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Brown TM, Colwell CS, Waschek JA, Piggins HD. Disrupted neuronal activity rhythms in the suprachiasmatic nuclei of vasoactive intestinal polypeptide-deficient mice. J Neurophysiol. 2007;97:2553–2558. doi: 10.1152/jn.01206.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Chaudhury D, Colwell CS. Circadian modulation of learning and memory in fear-conditioned mice. Behavioural Brain Research. 2002;133:95–108. doi: 10.1016/s0166-4328(01)00471-5. [DOI] [PubMed] [Google Scholar]
- 18.Zuker M. Mfold web server for nucleic acid folding and hybridization prediction. In Nucleic Acids Res. 2003:3406–3415. doi: 10.1093/nar/gkg595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ishida A, Mutoh T, Ueyama T, Bando H, Masubuchi S, Nakahara D, Tsujimoto G, Okamura H. Light activates the adrenal gland: Timing of gene expression and glucocorticoid release. Cell Metabolism. 2005;2:297–307. doi: 10.1016/j.cmet.2005.09.009. [DOI] [PubMed] [Google Scholar]
- 20.Besson J, Rotsztejn W, Laburthe M, Epelbaum J, Beaudet A, Kordon C, Rosselin G. Vasoactive intestinal peptide (VIP): brain distribution, subcellular localization and effect of deafferentation of the hypothalamus in male rats. Brain Res. 1979;165:79–85. doi: 10.1016/0006-8993(79)90046-5. [DOI] [PubMed] [Google Scholar]
- 21.Hoorneman EM, Buijs RM. Vasopressin fiber pathways in the rat brain following suprachiasmatic nucleus lesioning. Brain Res. 1982;243:235–241. doi: 10.1016/0006-8993(82)90246-3. [DOI] [PubMed] [Google Scholar]
- 22.Mezey E, Kiss JZ. Vasoactive intestinal peptide-containing neurons in the paraventricular nucleus may participate in regulating prolactin secretion. Proc Natl Acad Sci U S A. 1985;82:245–247. doi: 10.1073/pnas.82.1.245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Watts AG, Swanson LW, Sanchez-Watts G. Efferent projections of the suprachiasmatic nucleus: I. Studies using anterograde transport of Phaseolus vulgaris leucoagglutinin in the rat. J Comp Neurol. 1987;258:204–229. doi: 10.1002/cne.902580204. [DOI] [PubMed] [Google Scholar]
- 24.Kallo I, Kalamatianos T, Wiltshire N, Shen S, Sheward WJ, Harmar AJ, Coen CW. Transgenic approach reveals expression of the VPAC2 receptor in phenotypically defined neurons in the mouse suprachiasmatic nucleus and in its efferent target sites. Eur J Neurosci. 2004;19:2201–2211. doi: 10.1111/j.0953-816X.2004.03335.x. [DOI] [PubMed] [Google Scholar]
- 25.Maywood ES, Reddy AB, Wong GK, O'Neill JS, O'Brien JA, McMahon DG, Harmar AJ, Okamura H, Hastings MH. Synchronization and maintenance of timekeeping in suprachiasmatic circadian clock cells by neuropeptidergic signaling. Curr Biol. 2006;16:599–605. doi: 10.1016/j.cub.2006.02.023. [DOI] [PubMed] [Google Scholar]
- 26.Sheward WJ, Maywood ES, French KL, Horn JM, Hastings MH, Seckl JR, Holmes MC, Harmar AJ. Entrainment to feeding but not to light: circadian phenotype of VPAC2 receptor-null mice. J Neurosci. 2007;27:4351–4358. doi: 10.1523/JNEUROSCI.4843-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Oster H, Damerow S, Kiessling S, Jakubcakova V, Abraham D, Tian J, Hoffmann MW, Eichele G. The circadian rhythm of glucocorticoids is regulated by a gating mechanism residing in the adrenal cortical clock. Cell Metab. 2006;4:163–173. doi: 10.1016/j.cmet.2006.07.002. [DOI] [PubMed] [Google Scholar]
- 28.Bodnar M, Sarrieau A, Deschepper CF, Walker CD. Adrenal vasoactive intestinal peptide participates in neonatal corticosteroid production in the rat. American Journal of Physiology-Regulatory Integrative and Comparative Physiology. 1997;42:R1163–R1172. doi: 10.1152/ajpregu.1997.273.3.R1163. [DOI] [PubMed] [Google Scholar]
- 29.Hinson JP, Renshaw D, Carroll M, Kapas S. Regulation of rat adrenal vasoactive intestinal peptide content: effects of adrenocorticotropic hormone treatment and changes in dietary sodium intake. J Neuroendocrinol. 2001;13:769–773. doi: 10.1046/j.1365-2826.2001.00692.x. [DOI] [PubMed] [Google Scholar]
- 30.Cunningham LA, Holzwarth MA. Autoradiographic distribution of 125I-VIP binding in the rat adrenal cortex. Peptides. 1989;10:1105–1108. doi: 10.1016/0196-9781(89)90196-4. [DOI] [PubMed] [Google Scholar]
- 31.Holzwarth MA. The distribution of vasoactive intestinal peptide in the rat adrenal cortex and medulla. J Auton Nerv Syst. 1984;11:269–283. doi: 10.1016/0165-1838(84)90041-9. [DOI] [PubMed] [Google Scholar]
- 32.Karacay B, O'Dorisio MS, Summers M, Robinson M, Bonthius DJ. VIP receptor 1 (VPAC1) promoter targets the expression of a reporter gene to cerebellum and adrenal medulla in transgenic mice. Regul Pept. 2003;116:1–12. doi: 10.1016/s0167-0115(03)00170-8. [DOI] [PubMed] [Google Scholar]
- 33.Harmar AJ, Sheward WJ, Morrison CF, Waser B, Gugger M, Reubi JC. Distribution of the VPAC2 receptor in peripheral tissues of the mouse. Endocrinology. 2004;145:1203–1210. doi: 10.1210/en.2003-1058. [DOI] [PubMed] [Google Scholar]
- 34.Ehrhart-Bornstein M, Bornstein SR, Scherbaum WA, Pfeiffer EF, Holst JJ. Role of the vasoactive intestinal peptide in a neuroendocrine regulation of the adrenal cortex. Neuroendocrinology. 1991;54:623–628. doi: 10.1159/000125969. [DOI] [PubMed] [Google Scholar]
- 35.Cunningham LA, Holzwarth MA. Vasoactive intestinal peptide stimulates adrenal aldosterone and corticosterone secretion. Endocrinology. 1988;122:2090–2097. doi: 10.1210/endo-122-5-2090. [DOI] [PubMed] [Google Scholar]
- 36.Dijkstra I, Binnekade R, Tilders FJ. Diurnal variation in resting levels of corticosterone is not mediated by variation in adrenal responsiveness to adrenocorticotropin but involves splanchnic nerve integrity. Endocrinology. 1996;137:540–547. doi: 10.1210/endo.137.2.8593800. [DOI] [PubMed] [Google Scholar]
- 37.Morita Y, Yanagida D, Shintani N, Ogita K, Nishiyama N, Tsuchida R, Hashimoto H, Baba A. Lack of trimethyltin (TMT)-induced elevation of plasma corticosterone in PACAP-deficient mice. Ann N Y Acad Sci. 2006;1070:450–6. doi: 10.1196/annals.1317.060. [DOI] [PubMed] [Google Scholar]
- 38.Agarwal A, Halvorson LM, Legradi G. Pituitary adenylate cyclase-activating polypeptide (PACAP) mimics neuroendocrine and behavioral manifestations of stress: Evidence for PKA-mediated expression of the corticotropin-releasing hormone (CRH) gene. Brain Res Mol Brain Res. 2005;138(1):45–57. doi: 10.1016/j.molbrainres.2005.03.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Girard BA, Lelievre V, Braas KM, Razinia T, Vizzard MA, Ioffe Y, El Meskini R, Ronnett GV, Waschek JA, May V. Noncompensation in peptide/receptor gene expression and distinct behavioral phenotypes in VIP- and PACAP-deficient mice. J Neurochem. 2006;99(2):499–513. doi: 10.1111/j.1471-4159.2006.04112.x. [DOI] [PubMed] [Google Scholar]
- 40.Dunn J, Scheving L, Millet P. Circadian variation in stress-evoked increases in plasma corticosterone. Am J Physiol. 1972;223:402–406. doi: 10.1152/ajplegacy.1972.223.2.402. [DOI] [PubMed] [Google Scholar]
- 41.Kant GJ, Mougey EH, Meyerhoff JL. Diurnal variation in neuroendocrine response to stress in rats: plasma ACTH, beta-endorphin, beta-LPH, corticosterone, prolactin and pituitary cyclic AMP responses. Neuroendocrinology. 1986;43:383–390. doi: 10.1159/000124553. [DOI] [PubMed] [Google Scholar]
- 42.Scarbrough K, Harney JP, Rosewell KL, Wise PM. Acute effects of antisense antagonism of a single peptide neurotransmitter in the circadian clock. American Journal of Physiology-Regulatory Integrative and Comparative Physiology. 1996;39:R283–R288. doi: 10.1152/ajpregu.1996.270.1.R283. [DOI] [PubMed] [Google Scholar]
- 43.Roenneberg T, Daan S, Merrow M. The art of entrainment. Journal of Biological Rhythms. 2003;18:183–194. doi: 10.1177/0748730403018003001. [DOI] [PubMed] [Google Scholar]
- 44.Sakamoto K, Ishida N. Light-induced phase-shifts in the circadian expression rhythm of mammalian period genes in the mouse heart. Eur J Neurosci. 2000;12:4003–4006. doi: 10.1046/j.1460-9568.2000.00302.x. [DOI] [PubMed] [Google Scholar]
- 45.Kawamoto K, Nagano M, Kanda F, Chihara K, Shigeyoshi Y, Okamura H. Two types of VIP neuronal components in rat suprachiasmatic nucleus. J Neurosci Res. 2003;74:852–857. doi: 10.1002/jnr.10751. [DOI] [PubMed] [Google Scholar]
- 46.Shigeyoshi Y, Taguchi K, Yamamoto S, Takekida S, Yan L, Tei H, Moriya T, Shibata S, Loros JJ, Dunlap JC, Okamura H. Light-induced resetting of a mammalian circadian clock is associated with rapid induction of the mPer1 transcript. Cell. 1997;91:1043–1053. doi: 10.1016/s0092-8674(00)80494-8. [DOI] [PubMed] [Google Scholar]
- 47.Jasper MS, Engeland WC. Splanchnic neural activity modulates ultradian and circadian rhythms in adrenocortical secretion in awake rats. Neuroendocrinology. 1994;59:97–109. doi: 10.1159/000126645. [DOI] [PubMed] [Google Scholar]
- 48.Jasper MS, Engeland WC. Splanchnicotomy increases adrenal sensitivity to ACTH in nonstressed rats. Am J Physiol. 1997;273:E363–368. doi: 10.1152/ajpendo.1997.273.2.E363. [DOI] [PubMed] [Google Scholar]
- 49.Bittman EL, Doherty L, Huang L, Paroskie A. Period gene expression in mouse endocrine tissues. Am J Physiol Regul Integr Comp Physiol. 2003;285:R561–569. doi: 10.1152/ajpregu.00783.2002. [DOI] [PubMed] [Google Scholar]
- 50.Oster H, Damerow S, Hut RA, Eichele G. Transcriptional profiling in the adrenal gland reveals circadian regulation of hormone biosynthesis genes and nucleosome assembly genes. J Biol Rhythms. 2006;21:350–361. doi: 10.1177/0748730406293053. [DOI] [PubMed] [Google Scholar]
- 51.Yoo SH, Yamazaki S, Lowrey PL, Shimomura K, Ko CH, Buhr ED, Siepka SM, Hong HK, Oh WJ, Yoo OJ, Menaker M, Takahashi JS. PERIOD2::LUCIFERASE real-time reporting of circadian dynamics reveals persistent circadian oscillations in mouse peripheral tissues. Proc Natl Acad Sci U S A. 2004;101:5339–5346. doi: 10.1073/pnas.0308709101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Kalsbeek A, Palm IF, La Fleur SE, Scheer FAJL, Perreau-Lenz S, Ruiter M, Kreier F, Cailotto C, Buijs RM. SCN outputs and the hypothalamic balance of life. Journal of Biological Rhythms. 2006;21:458–469. doi: 10.1177/0748730406293854. [DOI] [PubMed] [Google Scholar]
- 53.Le Minh N, Damiola F, Tronche F, Schutz G, Schibler U. Glucocorticoid hormones inhibit food-induced phase-shifting of peripheral circadian oscillators. Embo J. 2001;20:7128–7136. doi: 10.1093/emboj/20.24.7128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Oishi K, Amagai N, Shirai H, Kadota K, Ohkura N, Ishida N. Genome-wide Expression Analysis Reveals 100 Adrenal Gland-dependent Circadian Genes in the Mouse Liver. DNA Res. 2005;12:191–202. doi: 10.1093/dnares/dsi003. [DOI] [PubMed] [Google Scholar]
- 55.Segall LA, Perrin JS, Walker CD, Stewart J, Amir S. Glucocorticoid rhythms control the rhythm of expression of the clock protein, Period2, in oval nucleus of the bed nucleus of the stria terminalis and central nucleus of the amygdala in rats. Neuroscience. 2006;140:753–757. doi: 10.1016/j.neuroscience.2006.03.037. [DOI] [PubMed] [Google Scholar]