

Abstract
This study tested the hypothesis that skeletal muscles generate more mechanical energy in gait tasks that raise the center of mass compared to the mechanical energy they dissipate in gait tasks that lower the center of mass despite equivalent changes in total mechanical energy. Thirteen adults ran on a 10° decline and incline surface at a constant average velocity. Three-dimensional (3D) joint powers were calculated from ground force and 3D kinematic data using inverse dynamics. Joint work was calculated from the power curves and assumed to be due to skeletal muscle–tendon actuators. External work was calculated from the kinematics of the pelvis through the gait cycle. Incline vs. decline running was characterized with smaller ground forces that operated over longer lever arms causing larger joint torques and work from these torques. Total lower extremity joint work was 28% greater in incline vs. decline running (1.32 vs. −1.03 J/kg m, p<0.001). Total lower extremity joint work comprised 86% and 71% of the total external work in incline (1.53 J/kg m) and decline running (−1.45 J/kg m), which themselves were not significantly different (p<0.180). We conjectured that the larger ground forces in decline vs. incline running caused larger accelerations of all body tissues and initiated a greater energy-dissipating response in these tissues compared to their response in incline running. The runners actively lowered themselves less during decline stance and descended farther as projectiles than they lifted themselves during incline stance and ascended as projectiles. These data indicated that despite larger ground forces in decline running, the reduced displacement during downhill stance phases limited the work done by muscle contraction in decline compared to incline running.
Keywords: Locomotion, Gait, Joint power, Incline, Decline, Ramp, Biomechanics
1. Introduction
Locomotion is a basic human behavior that transports us through the environment in a relatively healthy and efficient manner. Locomotion is performed through a combination of energy generating, shortening contractions and energy dissipating, lengthening contractions in skeletal muscle. These contractions combine to maintain a constant amount of total body mechanical energy in level gaits (Donelan et al., 2002; Laursen et al., 2000; Minetti et al., 1993), or to increase or decrease mechanical energy in non-level, ascending or descending gaits (Daley and Biewener, 2003; Gabaldon et al., 2004; Gottschall and Kram, 2006; McIntosh et al., 2006; Roberts and Belliveau, 2005). Although non-level gaits that maintain a constant average velocity have equivalent increases and decreases in total body mechanical energy per unit of vertical displacement, recent evidence suggests that actual muscle work is biased towards greater energy generation than dissipation in inclined vs. declined gaits of equal vertical displacement. Inclined vs. declined gaits have longer stance durations and higher average joint powers (Lay et al., 2007; McFadyen and Winter, 1988; McIntosh et al., 2006; Riener et al., 2002) and this combination of factors dictates that muscle work derived from joint powers would be greater during the stance phase in ascent than in descent. When combined across all lower limb joints, data from Duncan et al. (1997) showed that positive muscle work was two- to threefold greater in stair ascent compared to negative muscle work in stair descent and our data showed a smaller but reliable 17% bias towards positive over negative muscle work in stair ascent vs. descent (DeVita et al., 2001).
These observations led us to directly investigate the nature of muscular contributions to mechanical energy fluctuations in ascending and descending stair and ramp walking (DeVita et al., 2007). We assessed the positive, negative, and net areas under the lower extremity joint power curves during the stance phases of these gaits and we summed these values across the joints assuming this work was performed by skeletal muscles (e.g. (Dutto et al., 2006; Elftman, 1939; Winter, 1983). Net muscle work in ramp ascent was 25% greater than net muscle work in ramp descent (89 vs. −71 J m−1, p<0.010) and net muscle work in stair ascent was 43% greater than net muscle work in stair descent (107 vs. −75 J step−1, p<0.000). While these data were limited to sagittal plane, lower extremity joint powers during the stance phase, they were compelling enough to lead us to hypothesize that skeletal muscles generate more mechanical energy in gait tasks that raise the center of mass compared to the mechanical energy they dissipate in gait tasks that lower the center of mass, despite equivalent changes in total mechanical energy. This hypothesis implies that muscle function is biased towards energy-generating shortening contractions over that of energy-dissipating lengthening contractions in locomotion. We now test this hypothesis with more rigorous protocols than in our previous work. The purposes of this study were to compare three-dimensional (3D), lower extremity joint powers and work throughout the entire stride cycle in incline and decline running at a constant average velocity and to compare these work values to the total external work performed during the complete strides. We expected to observe a greater magnitude of muscle work and that muscle work was a greater proportion of the total external work in incline vs. decline running.
2. Methods
2.1. Participants
Thirteen adults, 8 males and 5 females, volunteered for the study. Their mean (sd) age, mass, height, and BMI were 22.3 (2.8) yr, 69.9 (13.2) kg, 1.74 (0.10) m, and 23.1 (2.8) kg/m2. All participants were healthy, recreational athletes without history of lower extremity injury. All participants gave written informed consent before participating according to University policy.
2.2. Experimental set-up
A 5 m ramp inclined 10° was fitted with a force platform (AMTI OR6-6-2000) in the middle of the ramp. The ramp itself was located in the middle of 6 m long level runways at both ends. Three-dimensional ground forces and the free moments were measured with the force platform at 960 Hz. Three-dimensional kinematics were recorded at 240 Hz with an eight camera infrared digital system (Qualisys MacReflex 240). An infrared timing system (Brower Timing Systems, Model IRD-T175) was used to constrain running speed to 3.35 m/s in both conditions.
2.3. Testing protocol
Participants wore bicycle shorts, a tight fitting T-shirt, and running shoes. Reflective markers were placed on the participants’ right leg and on the pelvis with a modified Helen Hayes marker set to identify lower extremity and pelvic kinematics. Participants practiced running on the level approach and continued up or down the ramp to acclimate to the speed and force plate location constraints. Five trials were collected for each subject and running condition. Experimental trials were immediately re-collected if the subject’s velocity was more than 5% different than the target speed, if the foot was not completely on the force platform or if the subject made obvious stride alterations to contact the force platform. Additionally, the resultant velocity of a marker on the L5/S1 joint was inspected immediately after the trial to verify the subject maintained a relatively constant average velocity through the trial. The order of the conditions was counterbalanced among the participants.
2.4. Data reduction
Cartesian coordinates of the markers describing the flight and the stance phase on the force platform were processed through a second-order low-pass digital filter with a 6 Hz cut-off frequency. Inverse dynamics using Visual 3D software (C-Motion Inc., Rockville, MD) were used to calculate the 3D joint torques at each lower extremity joint throughout the stride cycle. Extensor angular impulses were calculated from the torque curves. Three-dimensional joint powers were calculated as the product of the joint torques and joint angular velocities. Total power curves within each anatomical plane were then calculated as the sum of the hip, knee, and ankle joint powers and used to provide visual descriptions of the simultaneous power outputs from the three joints. Positive, negative, and net joint work in all planes throughout the stride were calculated from the hip, knee, and ankle powers as the areas under the joint power–time curves. Positive and negative work indicated that the muscles generated or dissipated mechanical energy (Alexander, 2004; DeVita et al., 2007; Dutto et al., 2006; Elftman, 1939; Roberts and Belliveau, 2005; Winter, 1983). We emphasized work in the sagittal plane here because it was 89% of the total joint work while frontal and transverse plane work were 9% and 2% of the total. We use the term muscle in this study to mean the entire muscle–tendon complex.
We estimated the change in total body energy (i.e. total external work) in each running task by calculating the net change in body gravitational potential energy (ΔPE) and linear kinetic energy (ΔKE) over the stride cycle. We used the vertical position and resultant velocity of the L5/S1 joint at the start and end of the stride cycle in these calculations. We assumed the work associated with the positions and velocities of the body segments relative to the body center of mass was zero over the course of the stride cycle (i.e. the positions and velocities of the limbs relative to the body center of mass were identical at the initial and final toe off points marking the start and end of the cycle). We used ΔKE to verify that running velocity and therefore kinetic energy were constant over the stride.
Total external work and joint work were dependent upon stride length because variation in this displacement affected vertical displacement and PE. Stride length was 4% shorter in decline vs. incline running (2.37 vs. 2.48 m, p<0.025) and due to this difference in stride length, work variables were normalized to both body mass and stride length and expressed in units of J/kg m for all comparisons.
2.5. Statistical analysis
Variables describing selected kinematics, ground forces, energetic states, torque, and work were compared between decline and incline running with one tail paired t-tests. The differences between total joint work and total external work within decline and incline running were also analyzed with one tail paired t-tests. Alpha levels were set to 0.05 in all comparisons.
3. Results
The goals of the tasks were to maintain a constant running velocity (i.e. kinetic energy) over the stride cycles while deleting or adding potential energy during decline and incline running. Differences in ΔKE over the cycle were 24 J (5%) in decline and 4 J (<1%) in incline running (Fig. 1). Differences in ΔPE over the cycle were 257 J in decline and 269 J in incline running. The 4% difference in ΔPE between gaits was directly due to the 4% shorter stride length in decline vs. incline running. Overall, ΔKEs were minimal and ΔPEs were similar in incline and decline running. Thus, total external work on the runners in each gait was satisfactory for the present purpose and the small adjustment of work values to stride length further improved the quality of the data.
Fig. 1.

Means and standard deviations (n = 13) of the potential energy (PE) and kinetic energy (KE) at the beginning (initial) and end (final) of one full stride. KE was maintained through the stride cycles within 5% and 1% for decline and incline running. The changes in PE over the stride were less than 5% different between gaits (257 vs. 269 J in decline and incline). The protocol successfully manipulated PE while maintaining relatively consistent and similar KE values within and between gaits.
Vertical displacements during the strides were −0.38 and 0.40 m in decline and incline running (p<0.029, Fig. 2). These displacements were composed of projectile flight and non-projectile stance phases. Decline vs. incline running had 44% larger displacement during flight (0.13 vs. 0.09 m, p<0.001) and a 19% smaller displacement during stance (0.25 vs. 0.31 m, p<0.000).
Fig. 2.
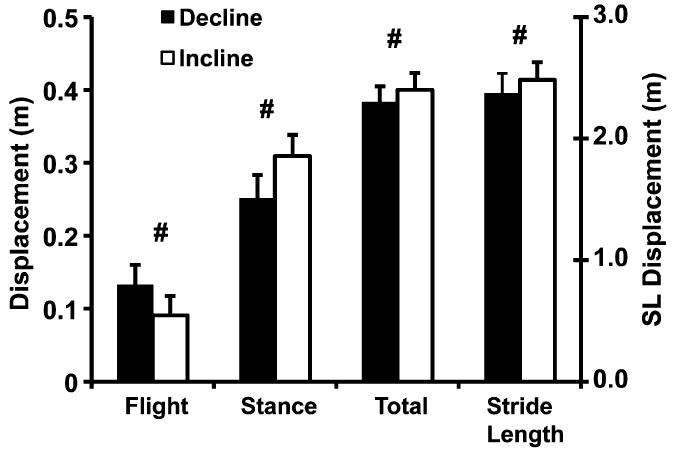
Means and standard deviations (n = 13) of the vertical displacement magnitudes (i.e. negative decline values in absolute values) during the flight, stance, and total stride phases and stride lengths in decline and incline running. Flight and stance values are doubled single limb values to account for both limbs through a complete stride. Decline vs. incline running had 44% larger displacement (p<0.001) during flight and a 19% smaller displacement (p<0.001) during stance. Decline vs. incline running also had a 4% smaller vertical displacement (p<0.029) through the total stride due to the 5% shorter stride length (p<0.026). The runners descended as projectiles farther in decline than they ascended as projectiles in incline running and they actively lowered themselves less during decline stance than they lifted themselves during incline stance.
Sagittal plane joint torques were similar between the gaits especially during swing phases (Fig. 3). The total extensor angular impulses during stance phases were 177% (p<0.000) and 49% (p<0.000) larger at the hip and ankle joints in incline vs. decline running. Decline running had 54% greater extensor angular impulse at the knee (p<0.002) compared to incline running. When summed across all joints the extensor angular impulse during stance was 27% larger in incline vs. decline running (p<0.000).
Fig. 3.

Mean (n = 13) sagittal plane joint torque– and power–time curves during decline (dashed) and incline (solid) running. Individual joint curves and the summed curves (Support and Total). Heel strike at 0.0 (vertical line) with swing followed by stance phases. Swing phase torques and powers were similar across gaits at each joint and in support and total curves. Stance phase joint torques were also similar between gaits with the largest difference observed at the hip joint. Gait inclination could be distinguished by stance phase joint powers and work which were directly related to slope such that decline running had more negative than positive work and incline running had more positive than negative work.
Hip joint power and work were biased towards positive, energy-generating roles in swing and stance phases of both gaits whereas knee joint power and work were biased towards negative, energy-dissipating roles in both gaits (Fig. 3). The ankle demonstrated similar negative power, work, and energy dissipation in the early stance phases for both gaits but altered positive work in direct relation to the slope of the running surface. Generally, swing phase powers were similar across running gaits at each joint and stance phase powers were dissimilar. Decline running was biased towards negative joint power and work whereas incline running was biased towards positive power and work during stance. Overall, the net joint work in swing was 12% and 7% of the total in decline and incline running whereas the net joint work in stance was 88% and 93% of the total net work in both gaits.
Both decline and incline running had negative and positive work at each joint (Fig. 4). The total net joint work was negative and positive in decline and incline running, respectively. Decline running had 38% greater total negative joint work compared to incline running (−1.06 vs. −0.77 J/kg m, p<0.000) and incline running had 265% greater total positive joint work than decline running (1.43 vs. 0.54 J/kg m, p<0.000). Negative work in both decline and incline running was performed primarily at the knee joint (decline: 63%, incline 58%) then secondarily at the ankle (decline: 23%, incline 30%) and hip joints (decline: 15%, incline 12%). Positive work was performed primarily at the hip joint in both gaits (decline: 48%, incline 55%) with smaller contributions at the ankle (decline: 32%, incline 32%) and knee (decline: 20%, incline 13%) joints.
Fig. 4.
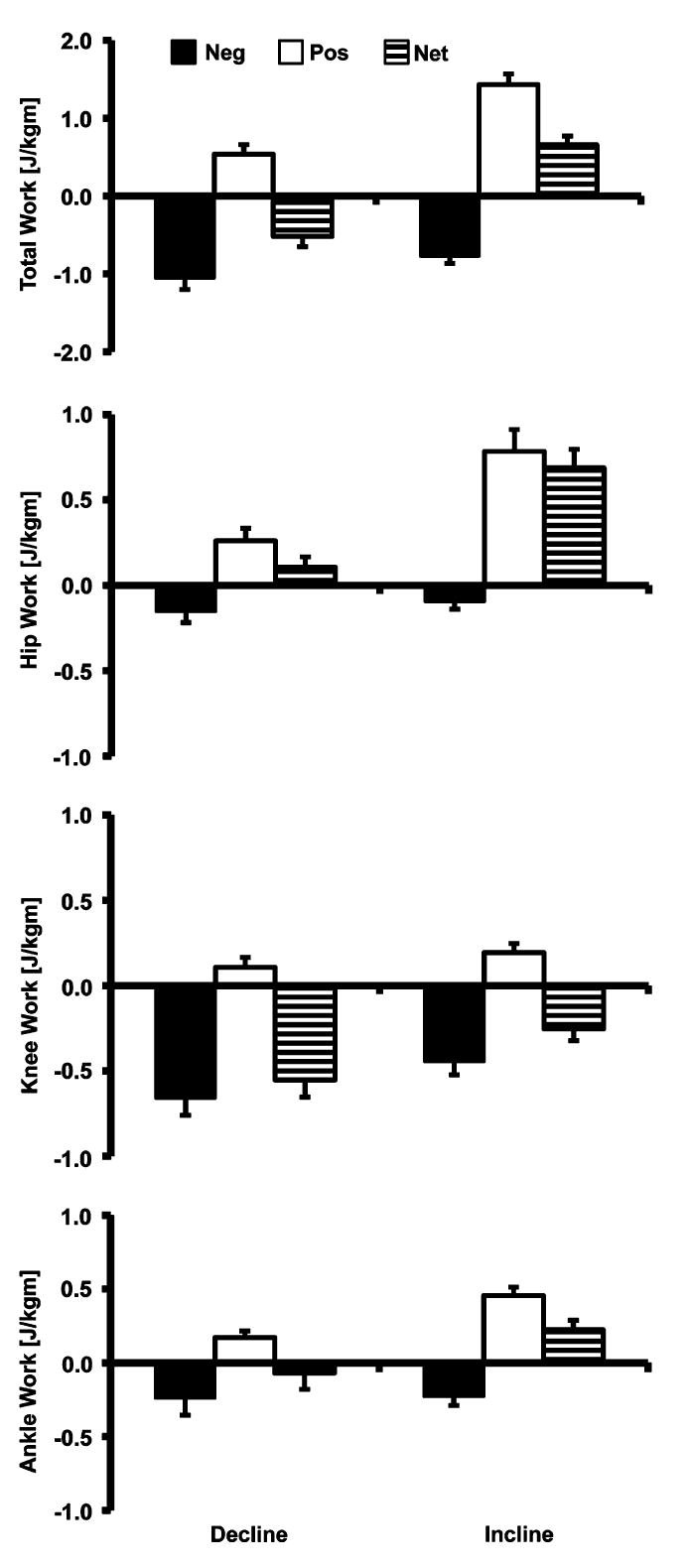
Means and standard deviations (n = 13) of the positive, negative and net joint work for the sum of all joints (Total) and hip, knee and ankle joints. Both gaits had negative and positive work phases but the net work corresponded to gait direction with decline having a net negative work and incline having a net positive work. Net work at the hip was positive in both gaits whereas net work at the knee was negative in both gaits. Ankle work corresponded to slope and was negative and positive in decline and incline running.
The magnitude of total lower extremity joint work was 28% greater in incline vs. decline running (1.32 vs. −1.03 J/kg m, p<0.001, Fig. 5). Additionally, total lower extremity joint work in decline running comprised only 71% of the total external work (−1.45 J/kg m), whereas total lower extremity joint work during incline running comprised 86% of the total external work (1.53 J/kg m).
Fig. 5.
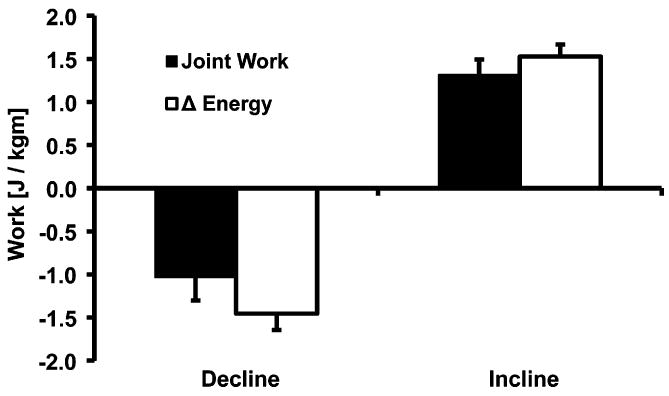
Means and standard deviations (n = 13) of the total joint work during the swing and stance phases and change in total energy (DEnergy) for decline and incline running. Joint work values are doubled to account for both lower extremities in the comparison with DEnergy values. Magnitude of total joint work was 28% greater in incline vs. decline running (1.32 vs. −1.03 J/kg m, p<0.001). Joint work in decline running comprised only 72% of the total external work (−1.45 J/kg m) whereas joint work during incline running comprised 86% of the total external work (1.53 J/kg m).
Normal and parallel ground forces were larger during the first half of stance in decline vs. incline running (Fig. 6). The normal force was 2.5-fold greater during weight acceptance in decline running (25.0 vs. 10.0 N/kg at ~0.04 s after heel strike). The direction of the resultant ground force vector relative to the joint centers was also affected by gait direction. The force vector was directed farther from the joint centers in incline vs. decline running altering the length of the moment arms for the ground force to the hip and ankle joints (Fig. 7). The average moment arm at the hip and the maximum moment arms at the hip and ankle were 29%, 50%, and 25% longer (all p<0.000) in incline vs. decline running.
Fig. 6.

Mean (n = 13) ground reaction force–time curves during decline (dashed) and incline (solid) running. The normal and parallel ground reaction forces were larger during the first half of stance in decline vs. incline running.
Fig. 7.

Means and standard deviations (n = 13) of the moment arm lengths for the ground forces during the stance phases of decline and incline running. (A) Average moment arm length over the stance phase. (B) Maximum moment arm length during stance phase. *Incline >decline, p<0.05.
4. Discussion
As in our previous work (DeVita et al., 2007), the basic assumption of this study was that work calculated from joint power was performed by skeletal muscle. Joint powers were produced by the net joint torques determined through inverse dynamics. These torques represented the muscular responses to torques applied to skeletal segments from external forces including ground and joint reaction forces, segmental weights, and inertial torques (Alexander, 1991; Bresler and Frankel, 1950; Elftman, 1939; Roberts and Belliveau, 2005; Winter, 1983). The deterministic relationship between joint torques and powers and muscle forces and work has been elegantly described in numerous studies investigating muscle mechanical advantage and alterations in muscle forces and joint torques as this advantage changes (e.g. (Biewener, 1989, 1990) and in other locomotion studies (Dutto et al., 2006; Elftman, 1939; Roberts and Belliveau, 2005; Winter, 1983). We also emphasize that our calculations do not address either muscle forces or the work produced by individual muscles. It is possible that muscle forces were larger in decline vs. incline and that through increased joint stiffness with greater muscle co-activation, the net work at each joint was reduced in decline running. Also, it is possible that individual muscles produced more negative and positive work in decline running with the net work being less in this gait than in incline running. Finally, our ground forces, joint torques and powers were highly similar to those in previous reports suggesting we successfully measured non-level running (Gottschall and Kram, 2005; Yokozawa et al., 2005, 2007).
These results closely agree with our previous work on walking on the same ramp used in this study (DeVita et al., 2007). We showed that positive muscle work was 25% greater than negative muscle work in the sagittal plane during the stance phases of incline vs. decline walking. Despite our more accurate assessment of muscle work in this study compared to our previous data, the positive biases in muscle work on the 10° slope were nearly identical in both studies. Present runners lowered themselves less during decline stance than they lifted themselves during incline stance and they descended as projectiles farther in decline running than they ascended as projectiles in incline running. Expressed in relative terms, 65% of the vertical displacement in decline running occurred during stance through primarily lengthening muscle contractions. In contrast 77% of the vertical displacement in incline running occurred during stance through primarily shortening contractions. Since external work is limited to the stance phase (i.e. vs. the flight phase), we can consider the linear mechanics due to the ground force and center of mass displacement during stance to shed light on the energetics of non-level running. Our data suggest that despite larger ground forces in decline running, the reduced displacement during downhill stance phases limits the work done by active muscle in decline compared to incline running.
We also conjectured that the reduced work in decline vs. incline gaits was due to the larger ground forces and the altered muscle mechanical advantage in decline vs. incline gaits (DeVita et al., 2007). The resultant ground forces at 0.04 s after heel strike were 25.5 and 10.0 N/kg body mass in decline and incline running, respectively. The much greater force in decline running most likely caused larger accelerations of all body tissues initiating a greater energy-dissipating response in non-muscular tissues including the heel pad, bones, and spinal discs (Li et al., 1995; Pain and Challis, 2001, 2002; Paul et al., 1978; Weijers et al., 2005) compared to their response in incline running. Additionally, vibrational movements of muscle bellies and tendons in response to the ground forces elicit energy dissipation but through a non-contractile mechanism (Pain and Challis, 2002; Wakeling et al., 2003; Wakeling and Nigg, 2001). We observed a similar result in landings from vertical falls in which landings with 23% greater impact forces had 18% less joint work (DeVita and Skelly, 1992) despite nearly identical changes in total body energy. Along with the difference in magnitude of the external ground forces between gaits, their directions relative to the joint centers were also affected by gait direction. The longer moment arms at the hip and ankle in incline running reduced the muscle mechanical advantage at these joints and ultimately led to larger total torque, power, and work despite having lower ground forces. Inclined running has been shown to increase the moment arm of the ground force at the hip compared to level running and this change was associated with greater torque and work output at the hip (Roberts and Belliveau, 2005) as seen presently. The reduced moment arms of the larger ground force in decline vs. incline running reduced the work demand on the contractile function of muscle (as in Alexander, 1991; Biewener, 1989; Chang et al., 2000); however, this force was still applied to the skeleton directly through foot contact with the floor and to muscle and other tissues through their attachments to the skeleton. In summary, despite larger ground forces in decline vs. incline running, the contractile load on skeletal muscles was reduced and compressive or vibrational loads on skeletal muscle and other tissues were most likely increased in decline compared to incline running.
Neural mechanisms may underlie the differences between ascending and descending gaits, linking mechanics to function. Although differences in environmental and mechanical circumstances between decline and incline running may pre-determine the biomechanics of the two movements, stopping the lowering of the center of mass during decline running after a shorter descent than the ascent in incline running can serve as an intrinsic safety mechanism. Pre-activation of muscles is one mechanism of reducing uncertainty associated with preparation for footfall during descending gaits such as downward stepping and landing (Hortobagyi and DeVita, 2000; Santello, 2005). Another mechanism mediating the earlier stopping of the center mass’s lowering during descending gaits, including decline running is the stretch reflex (Kamibayashi and Muro, 2006). In decline running, the rapid rate of change in the vertical ground reaction force peaked about 50 ms after heel strike (Fig. 6) and caused muscles to stretch, evoking, most likely a medium-latency volley (50–60 ms) (Matthews, 1991). These neural mechanisms in descent increase safety and also facilitate the early stopping of the center of mass’s lowering.
The data supported the hypothesis that skeletal muscles generate more mechanical energy in gait tasks that raise the center of mass compared to the mechanical energy they dissipate in gait tasks that lower the center of mass, despite equivalent changes in total mechanical energy. Incline vs. decline running was characterized with smaller ground forces that operated over longer lever arms causing larger joint torques and work from these torques. Based on the literature, we conjectured that the larger ground forces in decline running caused larger accelerations of all body tissues and initiated a greater energy-dissipating response in these tissues compared to their response in incline running. The runners actively lowered themselves less during decline stance and descended farther as projectiles than they lifted themselves during incline stance and ascended as projectiles. Despite larger ground forces in decline running, the reduced displacement during downhill stance phases limited the work done by muscle contraction in decline compared to incline running.
Acknowledgments
The authors thank Allison Gruber, Ben Long, and Kevin Via for their careful work with the study participants and in subsequent data analysis. This work was supported by NIH R01AG024161.
Footnotes
Conflict of interest statement The authors have no conflicts of interest interfering with this work.
References
- Alexander RM. Energy-saving mechanisms in walking and running. Journal of Experimental Biology. 1991;160:55–69. doi: 10.1242/jeb.160.1.55. [DOI] [PubMed] [Google Scholar]
- Alexander RM. Bipedal animals, and their differences from humans. Journal of Anatomy. 2004;204:321–330. doi: 10.1111/j.0021-8782.2004.00289.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Biewener AA. Scaling body support in mammals: limb posture and muscle mechanics. Science. 1989;245:45–48. doi: 10.1126/science.2740914. [DOI] [PubMed] [Google Scholar]
- Biewener AA. Biomechanics of mammalian terrestrial locomotion. Science. 1990;250:1097–1103. doi: 10.1126/science.2251499. [DOI] [PubMed] [Google Scholar]
- Bresler B, Frankel JP. The forces and moments in the leg during level walking. Transactions of the American Society of Engineers. 1950;72:27–36. [Google Scholar]
- Chang YH, Huang HW, Hamerski CM, Kram R. The independent effects of gravity and inertia on running mechanics. Journal of Experimental Biology. 2000;203(Pt 2):229–238. doi: 10.1242/jeb.203.2.229. [DOI] [PubMed] [Google Scholar]
- Daley MA, Biewener AA. Muscle force-length dynamics during level versus incline locomotion: a comparison of in vivo performance of two guinea fowl ankle extensors. Journal of Experimental Biology. 2003;206:2941–2958. doi: 10.1242/jeb.00503. [DOI] [PubMed] [Google Scholar]
- DeVita P, Skelly WA. Effect of landing stiffness on joint kinetics and energetics in the lower extremity. Medicine and Science in Sports and Exercise. 1992;24:108–115. [PubMed] [Google Scholar]
- DeVita P, Mizelle C, Vestal A, Beam S, Jolla J, Smith K, Hortobagyi T. Neuromuscular reorganization during stairway locomotion in old adults. Medicine and Science in Sports and Exercise. 2001;33:s344. [Google Scholar]
- DeVita P, Helseth J, Hortobagyi T. Muscles do more positive than negative work in human locomotion. Journal of Experimental Biology. 2007;210:3361–3373. doi: 10.1242/jeb.003970. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Donelan JM, Kram R, Kuo AD. Simultaneous positive and negative external mechanical work in human walking. Journal of Biomechanics. 2002;35:117–124. doi: 10.1016/s0021-9290(01)00169-5. [DOI] [PubMed] [Google Scholar]
- Duncan JA, Kowalk DL, Vaughan CL. Six degree of freedom joint power in stair climbing. Gait and Posture. 1997;5:204–210. [Google Scholar]
- Dutto DJ, Hoyt DF, Clayton HM, Cogger EA, Wickler SJ. Joint work and power for both the forelimb and hindlimb during trotting in the horse. Journal of Experimental Biology. 2006;209:3990–3999. doi: 10.1242/jeb.02471. [DOI] [PubMed] [Google Scholar]
- Elftman H. The function of muscles in locomotion. American Journal of Physiology. 1939;125:357–366. [Google Scholar]
- Gabaldon AM, Nelson FE, Roberts TJ. Mechanical function of two ankle extensors in wild turkeys: shifts from energy production to energy absorption during incline versus decline running. Journal of Experimental Biology. 2004;207:2277–2288. doi: 10.1242/jeb.01006. [DOI] [PubMed] [Google Scholar]
- Gottschall JS, Kram R. Ground reaction forces during downhill and uphill running. Journal of Biomechanics. 2005;38:445–452. doi: 10.1016/j.jbiomech.2004.04.023. [DOI] [PubMed] [Google Scholar]
- Gottschall JS, Kram R. Mechanical energy fluctuations during hill walking: the effects of slope on inverted pendulum exchange. Journal of Experimental Biology. 2006;209:4895–4900. doi: 10.1242/jeb.02584. [DOI] [PubMed] [Google Scholar]
- Hortobagyi T, DeVita P. Muscle pre- and coactivity during downward stepping are associated with leg stiffness in aging. Journal of Electromyography and Kinesiology. 2000;10:117–126. doi: 10.1016/s1050-6411(99)00026-7. [DOI] [PubMed] [Google Scholar]
- Kamibayashi K, Muro M. Modulation of pre-programmed muscle activation and stretch reflex to changes of contact surface and visual input during movement to absorb impact. Journal of Electromyography and Kinesiology. 2006;16:432–439. doi: 10.1016/j.jelekin.2005.09.004. [DOI] [PubMed] [Google Scholar]
- Laursen B, Ekner D, Simonsen EB, Voigt M, Sjogaard G. Kinetics and energetics during uphill and downhill carrying of different weights. Applied Ergonomics. 2000;31:159–166. doi: 10.1016/s0003-6870(99)00036-8. [DOI] [PubMed] [Google Scholar]
- Lay AN, Hass CJ, Nichols TR, Gregor RJ. The effects of sloped surfaces on locomotion: an electromyographic analysis. Journal of Biomechanics. 2007;40:1276–1285. doi: 10.1016/j.jbiomech.2006.05.023. [DOI] [PubMed] [Google Scholar]
- Li S, Patwardhan AG, Amirouche FM, Havey R, Meade KP. Limitations of the standard linear solid model of intervertebral discs subject to prolonged loading and low-frequency vibration in axial compression. Journal of Biomechanics. 1995;28:779–790. doi: 10.1016/0021-9290(94)00140-y. [DOI] [PubMed] [Google Scholar]
- Matthews PB. The human stretch reflex and the motor cortex. Trends in Neurosciences. 1991;14:87–91. doi: 10.1016/0166-2236(91)90064-2. [DOI] [PubMed] [Google Scholar]
- McFadyen BJ, Winter DA. An integrated biomechanical analysis of normal stair ascent and descent. Journal of Biomechanics. 1988;21:733–744. doi: 10.1016/0021-9290(88)90282-5. [DOI] [PubMed] [Google Scholar]
- McIntosh AS, Beatty KT, Dwan LN, Vickers DR. Gait dynamics on an inclined walkway. Journal of Biomechanics. 2006;39:491–502. doi: 10.1016/j.jbiomech.2005.07.025. [DOI] [PubMed] [Google Scholar]
- Minetti AE, Ardigo LP, Saibene F. Mechanical determinants of gradient walking energetics in man. Journal of Physiology. 1993;472:725–735. doi: 10.1113/jphysiol.1993.sp019969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pain MT, Challis JH. The role of the heel pad and shank soft tissue during impacts: a further resolution of a paradox. Journal of Biomechanics. 2001;34:327–333. doi: 10.1016/s0021-9290(00)00199-8. [DOI] [PubMed] [Google Scholar]
- Pain MT, Challis JH. Soft tissue motion during impacts: their potential contributions to energy dissipation. Journal of Applied Biomechanics. 2002;18:231–242. [Google Scholar]
- Paul IL, Munro MB, Abernethy PJ, Simon SR, Radin EL, Rose RM. Musculo-skeletal shock absorption: relative contribution of bone and soft tissues at various frequencies. Journal of Biomechanics. 1978;11:237–239. doi: 10.1016/0021-9290(78)90049-0. [DOI] [PubMed] [Google Scholar]
- Riener R, Rabuffetti M, Frigo C. Stair ascent and descent at different inclinations. Gait and Posture. 2002;15:32–44. doi: 10.1016/s0966-6362(01)00162-x. [DOI] [PubMed] [Google Scholar]
- Roberts TJ, Belliveau RA. Sources of mechanical power for uphill running in humans. Journal of Experimental Biology. 2005;208:1963–1970. doi: 10.1242/jeb.01555. [DOI] [PubMed] [Google Scholar]
- Santello M. Review of motor control mechanisms underlying impact absorption from falls. Gait and Posture. 2005;21:85–94. doi: 10.1016/j.gaitpost.2004.01.005. [DOI] [PubMed] [Google Scholar]
- Wakeling JM, Nigg BM. Soft-tissue vibrations in the quadriceps measured with skin mounted transducers. Journal of Biomechanics. 2001;34:539–543. doi: 10.1016/s0021-9290(00)00203-7. [DOI] [PubMed] [Google Scholar]
- Wakeling JM, Liphardt AM, Nigg BM. Muscle activity reduces soft-tissue resonance at heel-strike during walking. Journal of Biomechanics. 2003;36:1761–1769. doi: 10.1016/s0021-9290(03)00216-1. [DOI] [PubMed] [Google Scholar]
- Weijers RE, Kessels AG, Kemerink GJ. The damping properties of the venous plexus of the heel region of the foot during simulated heelstrike. Journal of Biomechanics. 2005;38:2423–2430. doi: 10.1016/j.jbiomech.2004.10.006. [DOI] [PubMed] [Google Scholar]
- Winter DA. Moments of force and mechanical power in jogging. Journal of Biomechanics. 1983;16:91–97. doi: 10.1016/0021-9290(83)90050-7. [DOI] [PubMed] [Google Scholar]
- Yokozawa T, Fujii N, Ae M. Kinetic characteristics of distance running on downhill slope. International Journal of Sport and Health Science. 2005;3:35–45. [Google Scholar]
- Yokozawa T, Fujii N, Ae M. Muscle activities of the lower limb during level and uphill running. Journal of Biomechanics. 2007;40:3467–3475. doi: 10.1016/j.jbiomech.2007.05.028. [DOI] [PubMed] [Google Scholar]