

Abstract
Activation of noradrenergic locus coeruleus (LC) neurons promotes wakefulness and behavioral arousal. In rats, LC neurons receive circadian inputs via a circuit that originates in the suprachiasmatic nucleus (SCN) and relays through the dorsomedial hypothalamus (DMH) to LC; this circuit input increases LC activity during the active period. DMH neurons expressing the peptide neurotransmitter orexin / hypocretin are ideally situated to act as a relay between SCN and LC due to their synaptic inputs from SCN and innervation of LC. Here, we examined the hypothesis that orexin is involved in transmitting circadian signals to LC using single-unit recordings of LC neurons in anesthetized rats maintained in 12:12 light-dark housing. We replicated earlier findings from this lab that LC neurons fire significantly faster on average during the active compared to rest periods. Local microinjection of an orexin antagonist, SB-334867-A attenuated the impulse activities of the fastest firing population of LC neurons during the active period. We also found that DMH orexin neurons project preferentially to LC and express a diurnal rhythm of activation that correlates with LC neuronal firing frequency. Therefore, we propose that DMH orexin neurons play a role in modulating the day-night differences of LC impulse activity.
Keywords: Orexin, hypocretin, locus coeruleus, norepinephrine, dorsomedial hypothalamus, circadian, arousal, sleep, waking
Introduction, 2
Locus coeruleus (LC) neuronal impulse activity is greatest during wakefulness (Aston-Jones and Bloom, 1981) and the LC plays a role in the circadian regulation of the sleep-wake cycle (Gonzalez and Aston-Jones, 2006). Anatomically, an indirect projection from the suprachiasmatic nuclei (SCN) via a relay in the dorsomedial hypothalamic area (DMH) provides circadian inputs that increase LC impulse activity during the active phase (Aston-Jones et al., 2001). Here we examined whether orexin, a wake-regulatory neuropeptide expressed in a subpopulation of DMH neurons, mediates this cyclical influence in LC.
Orexin is thought to consolidate wakefulness by increasing neuronal activation in wake-active brain areas, including the LC (Peyron et al., 1998; Hagan et al., 1999; Horvath et al., 1999; Ivanov and Aston-Jones, 2000;Piper et al., 2000; Sakurai, 2007). Orexin precursor mRNA, and hypothalamic and CSF orexin levels increase during the active period (Taheri et al., 2000; Yoshida et al., 2001; Zeitzer et al., 2003). Moreover, elevated orexin neuronal activity in the active period persists in constant darkness (Estabrooke et al., 2001), and the fluctuation of CSF orexin levels is abolished by SCN lesions (Zhang et al., 2004b), indicating that the anatomical inputs from the SCN (Yoshida et al., 2006) influence orexin neuronal activity in a circadian fashion. As LC neurons receive dense, direct orexin innervation (Peyron et al., 1998;Date et al., 1999; Horvath et al., 1999; Zhang et al., 2004a; Deurveilher and Semba, 2005; Espana et al., 2005), express orexin 1 receptors (Trivedi et al., 1998; Greco and Shiromani, 2001; Hervieu et al., 2001; Marcus et al., 2001; Bernard et al., 2002), and are excited by orexin (Hagan et al., 1999; Ivanov and Aston-Jones, 2000; Bernard et al., 2002; Soffin et al., 2002; van den Pol et al., 2002; Hoang et al., 2003; Walling et al., 2004), we hypothesized that orexin mediates day/night fluctuations in LC impulse activity.
DMH neurons project to the LC (Aston-Jones et al., 2001; Aston-Jones, 2005; Deurveilher and Semba, 2005) and the dorsal DMH contains orexin neurons (Peyron et al., 1998; Nambu et al., 1999; Chou et al., 2003; Deurveilher and Semba, 2005). Orexin neurons in the DMH and PeF, but not in the LH, have a diurnal activity rhythm (Estabrooke et al., 2001), and DMH orexin neurons receive direct synaptic inputs from the SCN (Yoshida et al., 2006). Whereas LH orexin neurons are active during reward-conditioned cues but not during stress or arousal per se (Harris et al., 2005), more medial orexin neurons appear to be involved in responses to stress and the maintenance of arousal during the wake phase of the circadian cycle (Estabrooke et al., 2001; Harris et al., 2005; Harris and Aston-Jones, 2006).
Using extracellular single-unit recordings in vivo, we demonstrate that elevated LC impulse activity in nocturnal rats begins shortly before dark onset and persists throughout the active phase, and that SB-334867-A, a selective orexin-1 receptor antagonist (Porter et al., 2001; Smart et al., 2001; Soffin et al., 2002), attenuates firing frequencies specifically in LC neurons with elevated firing frequencies during the active phase. Finally, we demonstrate that orexin neurons in the DMH preferentially project to the LC and show increased Fos expression at times when LC impulse activity is high. These results indicate that orexin inputs may provide an excitatory signal to LC neurons during the active period, and thereby participate in circadian regulation of sleep and waking.
Results, 3
Electrophysiology
Sixty-four individual LC neurons were recorded from 23 halothane-anesthetized rats throughout the sleep / wake cycle (ZT 5 – 20). Recordings during the active period were obtained at ZT 14–20, and during the rest period at ZT 5–10. Similar to a previous report from our lab (Aston-Jones et al., 2001), average LC impulse activity was greater during the active period than during the rest period (2.9 ± 0.4 spikes/sec vs. 1.8 ± 0.2 spikes/sec, n = 19 and 30 neurons and 5 and 15 animals, respectively, p < 0.05, t = 1.9, df = 34 by t-test). Local microinjection of ACSF had no effect on LC firing frequency (1.9 ± 0.6 spikes/sec pre-injection vs 1.8 ± 0.5 spikes/sec during ACSF, n = 7 neurons and 2 animals, p = 0.36, t = 1.0, df = 6). However, microinfusion of 100 µM orexin-A during the rest period significantly increased LC firing frequency in all neurons recorded (1.5 ± 0.5 spikes/sec pre-injection vs 3.5 ± 0.9 spikes/sec during orexin, n = 6 neurons and 3 animals, p = 0.009, t = 4.2, df = 5 Fig. 1).
Figure 1.

LC neurons are activated by local microinfusion of orexin A during the rest phase in the anesthetized rat. A, raw traces of an LC neuron before (upper trace) and during (lower trace) microinfusion of 100 µM orexin A from a micropipette adjacent to the recording pipette. B, average impulse activity is significantly increased during 100 µM orexin A microinfusion.
Diurnal differences in response to SB-334867-A
We tested whether orexin inputs contribute to the difference in LC firing frequencies found between rest and active phase recordings. Figure 2 shows traces recorded during the rest and active periods before and during 100 µM SB-334867-A infusion. Significant decreases in average LC firing frequencies were observed during SB-334867-A infusion in the active period (2.9 ± 0.4 spikes/sec pre-injection, 1.8 ± 0.3 spikes/sec during SB-334867-A, n = 19 neurons and 5 animals, p = 0.01, t = 2.7, df = 18). Moreover, firing frequencies in the presence of the antagonist during the active period were not substantially different from firing frequencies during the rest period (1.8 ± 0.3 Hz vs 1.9 ± 0.3 Hz, p = 0.8, t = 0.3, df = 34). In contrast, during the rest period similar local SB-334867-A had no significant effect on LC firing frequency (1.9 ± 0.3 spikes/sec pre-injection, 2.2 ± 0.4 spikes/sec during SB-334867-A, n = 17 neurons and 10 animals, p = 0.4, t = 0.9, df = 16). However, in the hour immediately preceding the onset of darkness (ZT 11 – 12, Fig. 3), local SB-334867-A inhibited LC firing frequencies to an extent resembling the inhibition during the active period (2.9 ± 0.4 spikes/sec pre-injection, 1.7 ± 0.3 spikes/sec during SB-334867, n = 16 neurons and 6 animals, p<0.001, t = 2.1, df = 15), suggesting that orexin inputs to the LC occur independent of light inputs.
Figure 2.

LC neuronal impulse activity during the rest (A, C) and active periods (B, D). Average impulse activity was greater during the active period than during the rest period in LC neurons. Raw traces (A, B) and composite firing frequencies (C, D) of LC neural responses to local microinfusion of the orexin 1 receptor antagonist SB334867-A (upper traces in A and B are pre-microinfusion and lower traces are during microinfusion). Whereas no significant change in firing was observed in average LC activity with SB-334867-A microinfusion during the rest period, SB-334867-A microinfusion during the active period significantly decreased LC firing frequencies . Each line in panels C and D indicates data from a single neuron.
Figure 3.

LC impulse activity and responses to SB-334867-A at ZT 11 – 12. A, SB-334867-A inhibited firing frequency during the “transition” period: ZT 11 – 12. Each line indicates data from a single neuron. B, For comparison, average impulse activity during the active and rest periods (averages of all neurons depicted in Fig. 2 C, D) were plotted next to the average frequency during the transition period. Average firing frequency and magnitude of response to SB-334867-A were similar in the transition period to the active period.
A subset of “fast” firing neurons were found during the active period that were not present during the rest period (Fig. 2c and d). All neurons included here, regardless of firing frequency, met the physiological criteria (e.g., waveform, response to noxious stimuli) established for LC neurons (see Materials and Methods). We asked whether fast-firing neurons exhibited a greater sensitivity to SB-334867-A than slower firing neurons during the active period. For this analysis we performed a median split to divide neurons recorded in the active period into those that fired faster vs slower than the median rate. Neurons recorded during the active period with higher firing rates than the median firing frequency of 2.3 spike/sec exhibited decreased firing following SB-334867-A (“fast” neurons: 4.2 ± 0.6 spikes/sec pre-injection, 1.6 ± 0.5 spikes/sec during SB-334867-A, n = 9, p < 0.0001, t = 5.5, df = 8) whereas cells that fired below the median frequency did not significantly change activity following SB-334867-A (“slow” neurons: 1.7 ± 0. 1 spikes/sec pre-injection, 1.9 ± 0.3 spikes/sec during SB-334867-A, n = 10, p = 0.8, t = 0.6, df = 9). We then used the same criterion of 2.3 spikes/sec to designate “fast” vs “slow” firing neurons in other time periods. A similar difference between the response to local SB-334867-A in “fast” (3.6 ± 0.4 spikes/sec pre-injection, 2.0 ± 0.5 spikes/sec during SB-334867-A, n = 10, p = 0.01, t = 4.5, df = 9) and “slow” (1.7 ± 0.2 spikes/sec pre-injection, 1.2 ± 0.4 spikes/sec during SB-334867-A, n = 6, p = 0.2, t = 0.4, df = 5) firing LC neurons was observed during the hour immediately preceding the onset of darkness (ZT 11 – 12, Fig. 3). Notably, this difference between “fast” and “slow” firing neurons was not observed for the effects of local SB-334867-A (p = 0.3, t = 0.4, df = 4) in LC neurons recorded during the rest period: firing rates = 3.7 ± 0.3 spikes/sec pre-injection, 4.0 ± 0.5 spikes/sec during SB-334867-A, n = 5 fast-firing cells; and 1.2 ± 0.2 spikes/sec pre-injection, 1.4 ± 0.3 spikes/sec during SB-334867-A, n = 12 slow-firing cells. Thus, faster-firing LC neurons shortly before and during the active period were preferentially inhibited by the orexin-1 receptor antagonist, suggesting a role for orexin in establishing the diurnal rhythmicity of LC firing frequency.
In a separate experiment, the response to footpad stimulation in 6 “fast” neurons that responded to 100 µM SB-334867-A was tested. No differences were found between the response magnitude to footpad stimulation prior to vs. during antagonist application (7.8 ± 1.2 spikes/sec pre-injection vs 8.4 ± 2.1 spikes/sec during SB-334867-A, p = 0.8, t = 0.3, df = 5, data not shown), indicating that SB-334867-A mediated inhibition is specific to orexin inputs since the glutamate-mediated LC neuronal response to footpad stimulation (Ennis et al., 1988; Ennis et al., 1992) remained intact.
DMH orexin neuron activation at ZT 11 – 12
Because we found “fast” firing LC neurons that reduced their firing frequency in the presence of the orexin receptor antagonist SB-334867-A in the hour immediately preceding dark, we hypothesized that orexin neuron activation should be similar at this time as during the active phase (Fig. 3B). To measure orexin neuronal activation, we counted Fos immunoreactivity in three major compartments of the hypothalamus in which orexin neurons are located, the DMH, PeF and LH. As shown in Fig. 4, Fos-IR in orexin neurons was elevated during the active period compared to the rest period in DMH (58.3 ± 6.6 % active vs 28.8 ± 3.5% rest period, n = 5 animals, p = 0.004, t = 4.0, df = 9) and PeF (42.9 ± 2.4 % active vs 20.4 ± 3.0 % rest period, n = 5 animals, p = 0.0004, t = 5.9, df = 9). This difference was not evident in LH orexin neurons (26.2 ± 1.8 % active vs 20.7 ± 4.4 % rest period, n = 5 animals, p = 0.28, t = 1.2, df = 9); this overall pattern of results resembled those previously reported (Estabrooke et al., 2001). However, in animals sacrificed 1.5 hr after the transition time from rest to active periods (ZT 11 – 12), only orexin neurons located in the DMH had elevated Fos-IR (57.7 ± 2.0 % transition vs 28.8 ± 3.5 % rest period, n = 5 animals, p = 0.0001, t = 7.0, df = 9); neither PeF (23.4 ± 2.2 % transition vs 20.4 ± 3.0 % rest period n = 5 animals, p = 0.45, t = 0.8, df = 9) nor LH (19.1 ± 1.2 % transition vs 20.7 ± 4.4 % rest period n = 5 animals, p = 0.74, t = 0.4, df = 9) orexin neurons showed an increase during this transition period compared to the rest period. These results implicate orexin neurons located in the DMH as responsible for the elevated LC firing frequencies observed for ZT 11 – 12. We also observed non-orexin neurons located in the same hypothalamic regions that were Fos-activated in these experiments, but they were not analyzed in detail.
Figure 4.
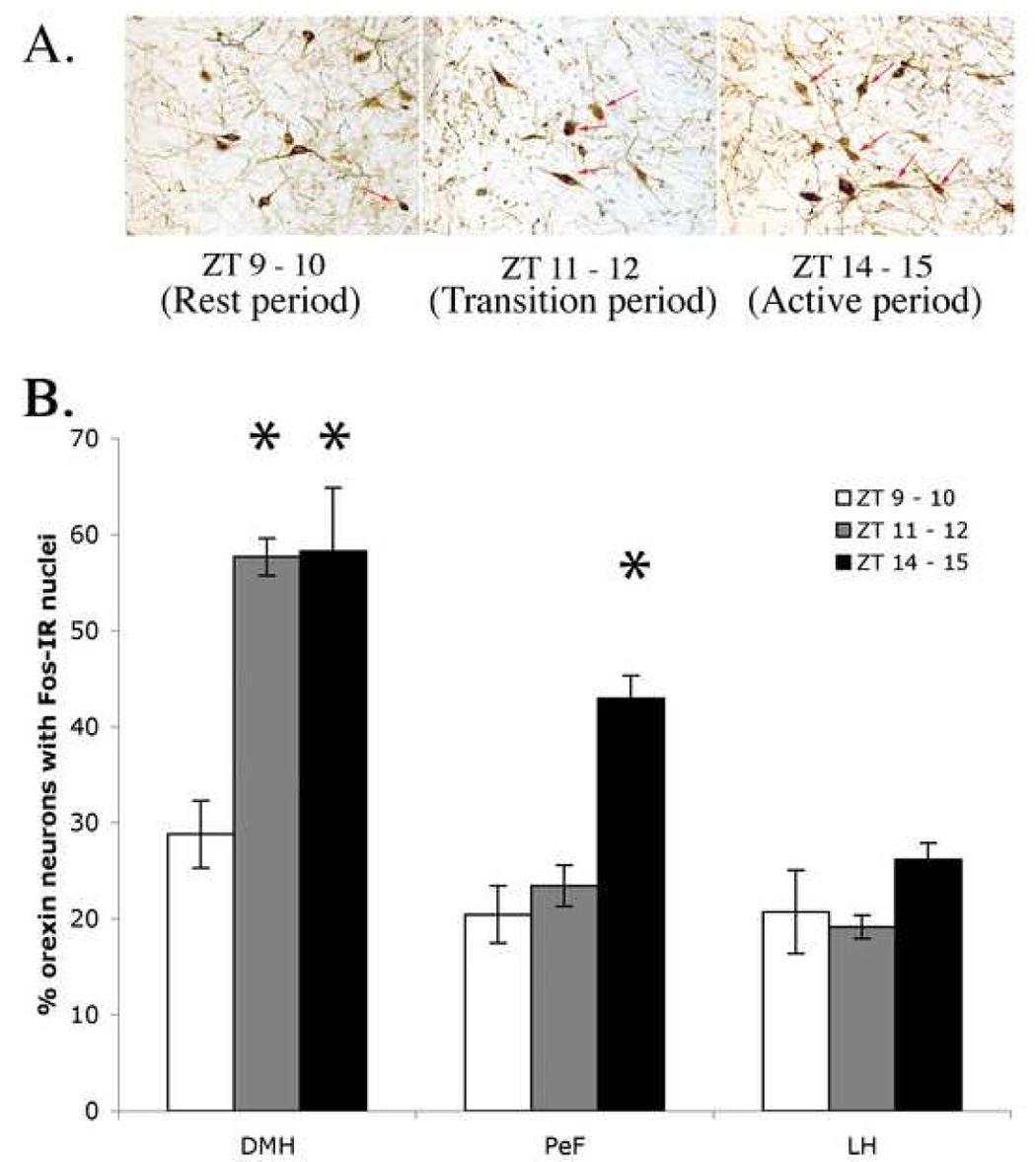
A, FOS expression in orexin-positive neurons in animals sacrificed 90 minutes after ZT 9–10, 11–12, or 14–15 in the dorsomedial hypothalamus. B, average counts of orexin-IR neurons that were also Fos-IR at ZT 9-10 (white bars), 11–12 (grey bars) and 14–15 (black bars) in the dorsomedial hypothalamus (DMH), perifornical (PeF), and lateral hypothalamus (LH). Fos-IR in orexin neurons was elevated during the active period compared to the rest period in the DMH and PeF, but not significantly in the LH. Only orexin neurons located in the DMH showed an increase in Fos immunoreactivity over ZT 9–10 at ZT 11–12 and neither PeF nor LH orexin neurons showed a similar increase. Asterisks denote significant differences from rest.
Orexin neuronal innervation of the LC
The DMH is responsible for relaying circadian temporal information from the SCN to the LC, and orexin neurons located in the DMH receive synaptic inputs from the SCN and express a circadian rhythm of activation. These findings led us to postulate that orexin neurons in the DMH provide a major input to the LC. To test this idea, we conducted retrograde tract-tracing to map inputs to LC from the different subfields of orexin neurons. Injections of the retrograde tracer cholera toxin b subunit (CTb) that encompassed the core LC nucleus and peri-LC dendritic zone yielded numerous retrogradely labeled neurons in the hypothalamus. Many of these retrogradely labeled LC afferent neurons also stained for orexin (Fig. 5). Of the total number of CTb-positive neurons counted from 8 animals in the DMH, PeF and LH, a significantly higher percentage also stained positive for orexin in the DMH and PeF (16 ± 1.1 % and 14.8 ± 5.9 %, respectively) than in the LH (9.5 ± 5.4 %, p < 0.05). Counting only orexin-positive neurons, a significantly greater percentage of orexin neurons in the DMH project to the LC (17.7 ± 1.1 %) than orexin neurons in either PeF or LH (5.9 ± 0.2 % and 5.4 ± 0.4 %, respectively, p < 0.05 each vs DMH). These results indicate that the bulk of orexin input to the rat LC originates in the DMH and PeF, and that orexin neurons in the DMH are more likely to project to the LC than orexin neurons in other hypothalamic subfields.
Figure 5.
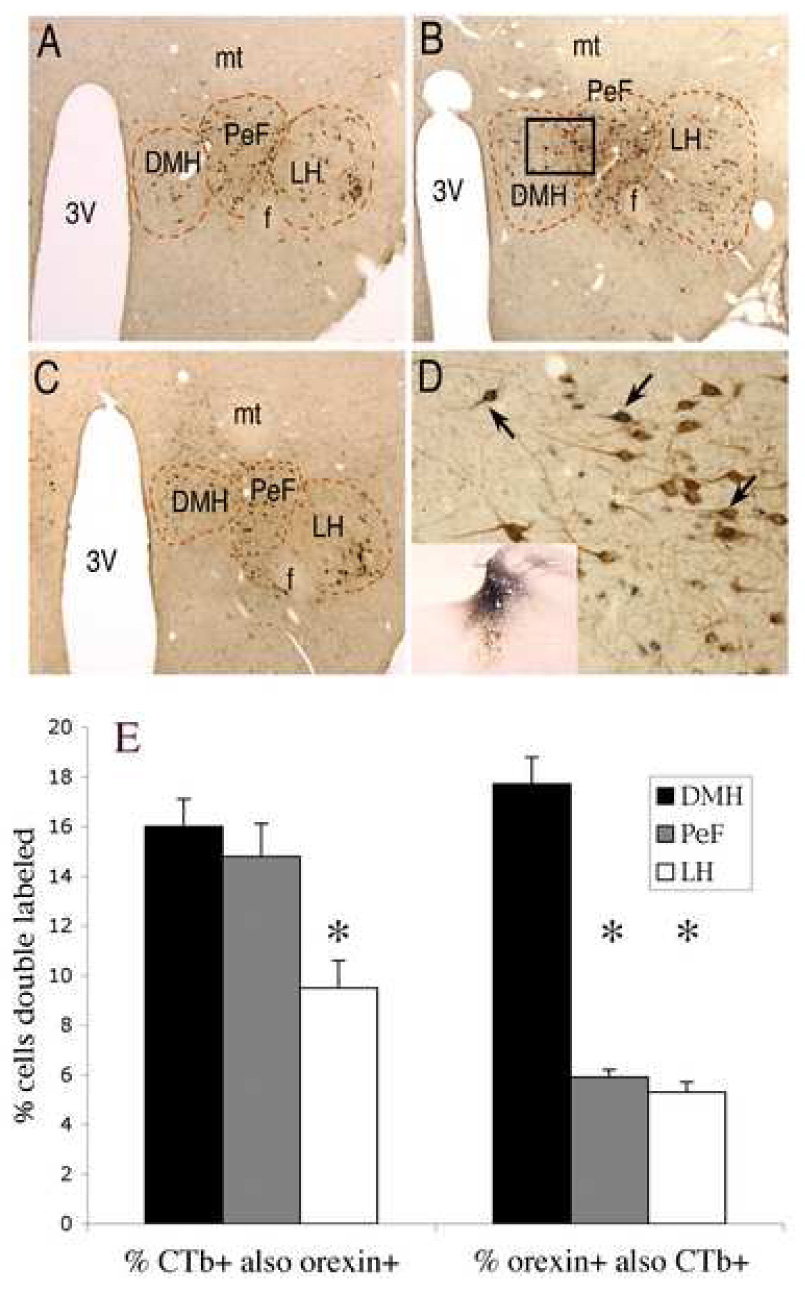
Double labeling of orexin afferents to the LC. A – C, Frontal sections at low-power through different levels of the rat hypothalamus (ordered caudal to rostral) double stained for orexin (brown) and CTb (retrogradely transported from the LC, dark blue). These images depict the different areas that were defined for counting orexin neurons: DMH, PeF and LH. Abbreviations: 3V – third ventricle; mt – mammillothalamic tract; f – fornix. D, High-power photo taken from box in panel B,showing doubly labeled neurons in the DMH and PeF regions (at arrows). Inset: example injection site of CTb in the LC. E: Bar graph summarizing counts of neurons labeled for orexin, CTb, or both. Note that most LC-afferent orexin neurons are located in the DMH and PeF with fewer in the LH. In addition, a igher percentage of orexin neurons in the DMH contain CTb, indicating that they project to LC, than in the PeF or the LH. Asterisks denote significant differenes (p <0.05) from DMH.
Discussion, 4
The data presented here indicate that orexin is involved in transmitting temporal information from the hypothalamus to noradrenergic LC neurons. The orexin receptor antagonist, SB-334867-A, microinfused onto LC neurons in vivo attenuated firing frequencies only in neurons recorded during the active period with relatively fast firing frequencies; LC neurons with lower impulse activity, or those recorded during the rest period, did not respond in any significant fashion to the antagonist. This finding, along with the complementary findings presented here that Fos expression in orexin neurons of the DMH coincides with increased LC neuronal activation and that these neurons preferentially project to the LC, suggest a role for DMH orexin neurons in the previously observed (Aston-Jones et al., 2001) increase in excitability of LC neurons during the active period.
Methodological considerations
Animals were not kept in constant darkness (DD), so any differences between the active and rest periods can only be described as diurnal and not circadian. However, the original work which serves as a basis for this study (Aston-Jones et al., 2001) showed that differences in impulse activity persisted in animals kept in constant darkness and can thus be considered circadian. Moreover, LC neurons recorded immediately preceding the active period had similar firing frequencies and responses to the orexin antagonist as neurons recorded in the middle of the active period. Since these neurons were recorded while animals were kept in the light, it’s likely that the underlying cause of the switch from rest to active period phenotype is mediated not by a light-sensing pathway but rather by an internal clock that anticipates the active period.
Our results are consistent with previous findings showing a diurnal rhythm in DMH orexin neuronal activation (Estabrooke et al., 2001). Here, we expand on these results to include a time-point just prior to the active period, when ambient lighting conditions are the same as during the rest phase, finding that orexin neurons anticipate the active period with elevated activity. Also, dorsal orexin neurons previously have been demonstrated to preferentially project to the LC (Espana et al., 2005). Here, we expand on these results to show that, when the orexin field is divided into three subdivisions, those orexin neurons located in the DMH preferentially project to the LC. These results, along with the previously documented lack of circadian activity rhythm in the LC following DMH lesions (Aston-Jones et al., 2001), are consistent with our view that DMH orexin neurons mediate circadian influences on LC. However, because we did not selectively lesion LH orexin neurons in this study, we cannot exclude the possibility that local network interactions between DMH and LH orexin neurons might relay temporal information from DMH to LC via the LH.
The vast majority of orexin neurons co-express dynorphin and glutamate (Abrahamson et al., 2001; Chou et al., 2001), yet in this case application of an orexin antagonist alone decreased LC firing during the active period. This suggests that orexin is the primary neurotransmitter for the transmission of temporal input to the LC. However, we did not examine the effects of either κ or µ-opioid, or ionotropic glutamate (NMDA or AMPA), receptor antagonists either alone or in conjunction with the orexin antagonist. It remains to be determined what role if any dynorphin and glutamate might play in the transmission of temporal information to the LC.
The recording conditions themselves might have affected the results as well. The advantage of using the anesthetized whole animal preparation is that, as opposed to slice electrophysiology of the LC (Soffin et al., 2002), the circuit output from the SCN remains sufficiently intact that its tonic effect on LC activity can be recorded under a similar behavioral state during either the active or rest periods (Aston-Jones et al., 2001). It is possible, however, that another anesthetic might alter the results—thus either increasing or decreasing the likelihood of recording neurons with elevated impulse activity during the active period. Further study might elucidate whether and how anesthetics affect the arousal-promoting properties of orexin neurons, and the effects of orexin inputs on LC neurons.
Role of orexin in transmitting temporal signals to the LC
There are numerous functional similarities between hypothalamic orexin neurons and LC neurons in their control of sleep-wake state. LC neuronal firing frequency is tightly associated with the organism’s sleep-wake state, being highest during wake, lower during slow wave sleep and almost silent during paradoxical (REM) sleep (Aston-Jones and Bloom, 1981;Pace-Schott and Hobson, 2002; Berridge and Waterhouse, 2003). Similarly, DMH orexin neuronal activation is correlated with sleep and waking (Estabrooke et al., 2001; Lee et al., 2005; Mileykovskiy et al., 2005), and extracellular orexin levels fluctuate with the diurnal and circadian period such that they are highest during the active period (Taheri et al., 2000;Lin et al., 2002). Moreover, orexin increases LC neuronal activation (Horvath et al., 1999; Ivanov and Aston-Jones, 2000) and decreases REM sleep duration when microinjected into the LC (Bourgin et al., 2000). The fact that the LC receives copious orexinergic innervation (Peyron et al., 1998; Espana et al., 2005) further emphasizes the functional relationship between these two areas in sleep-wake state control.
Although other neuronal phenotypes also project to LC from DMH, the fact that DMH lesions reduced LC firing frequency during the active period (Aston-Jones et al., 2001) indicates that excitatory DMH-LC projections (such as orexin) may be most important for circadian regulation of LC activity. However, a subset of LC neurons responded to the Orexin 1 receptor antagonist with a slight increase in firing frequency. Although we did not examine this possibility here, it is possible that this may reflect orexin innervation of GABAergic interneurons located in the pericerulear dendritic zone, that in turn inhibit LC neurons (Aston-Jones et al., 2004). Finally, as orexin neuronal activation has been linked to specific wakefulness events regardless of circadian time, it might be surmised that orexin has little to do with the circadian timing of sleep and wakefulness. However, the circadian fluctuation of CSF and extracellular orexin levels seems to suggest otherwise. Recently, a model that incorporates both slow, circadian and fast,phasic orexin release has been proposed which states that the background (slow, circadian) level of orexin may determine the physiological response to the fast, phasic release (Adamantis and de Lecea, 2008). Here, we provide evidence that the LC receives orexinergic circadian inputs and thus might be a site of action of such signals.
The excitability of LC neurons is influenced by indirect inputs from the SCN via the DMH (Aston-Jones et al., 2001) and LC lesions decrease the amplitude of the circadian rhythm of the sleep-wake cycle (Gonzalez and Aston-Jones, 2006), indicating that the circadian clock may influence behavior associated with wakefulness through the LC. The observation that the subpopulation of orexin neurons located in the DMH receives direct SCN inputs and projects to the LC indicates that orexin inputs to LC might be involved in this circadian function (Deurveilher and Semba, 2005; Yoshida et al., 2006). Here we expand on this by showing that the day-night difference in LC impulse activity, when the animal’s arousal is held constant by anesthesia, is due in large part to orexin. We further demonstrate that the subpopulation of orexin neurons located in the DMH preferentially projects to the LC. Together, these and other data indicate that orexin neurons in the DMH provide circadian regulation of LC impulse activity, and thereby influence circadian regulation of sleep and waking.
Experimental Procedure
Animals
Adult male Sprague Dawley rats (250 g, Harlan, Indianapolis, IN) were kept on a 12 hr LD cycle (lights on at 0800) for at least one week after arriving in the lab before experiments, and were given food and water ad libidum. Before electrophysiological recordings, animals were anesthetized with halothane in air via spontaneous respiration. Animals were intubated with a tracheal cannula, continuously anesthetized with 0.9 – 1.2 % halothane, and placed in a stereotaxic frame with the incisor bar lowered to place the skull approximately 12° from the horizontal plane. Body temperature was maintained at 34 – 36°C with a thermistor-controlled heating pad. The eyes of animals that were recorded during their dark period were covered with black tape to avoid any light input during set-up, surgery or recording. All procedures were approved by the Institutional Animal Care and Use Committee of the University of Pennsylvania.
Electrophysiology
LC recordings and microinjections were performed using standard methods as previously described (Shiekhattar and Aston-Jones, 1992; Jodo and Aston-Jones, 1997). In brief, a 5-mm-diameter hole was drilled in the skull above the LC and the dura was reflected. A glass recording micropipette (tip diameter, 2 – 4 µm; 10 – 20 MΩ impedance) was filled with 2% pontamine sky blue dye in 0.5 M Na-acetate and aimed at the LC (4 mm caudal to lambda, and 1.2 mm lateral to midline). Signals were amplified and continuously displayed on an oscilloscope as unfiltered and filtered (0.3 – 10 kHz bandpass). LC neurons (5.8 – 6.5 mm ventral to the skull surface) were tentatively identified during recording using well established criteria: an entirely positive, notched waveform 2 ms or more in duration in unfiltered records; a slow, tonic spontaneous discharge (0.5 – 6.0 Hz); and a strong phasic activation followed immediately by long-duration inhibition (0.5 – 1.0 s) in response to noxious stimuli (for example, tail or hindpaw compression). Following LC identification, recordings were continuously monitored for changes in spike amplitude to ensure stable recordings. Footpad stimulation (FS; to activate the sciatic nerve) was delivered through paired 26-gauge needles inserted in the medial aspect of the contralateral hindpaw and was given as monophasic 0.5 ms pulses of 90 V. Stimuli were given once every 2 s for a total of 50 repetitions. Spikes of single neurons were discriminated, and digital pulses were led to a computer for on-line data collection using a laboratory interface and software (CED 1401, SPIKE2; Cambridge Electronic Design, Cambridge, England). Response magnitudes to FS were calculated as the total number of spikes recorded from 20 to 100 ms following stimulation minus the expected number of spikes (average number of spikes / sec recorded 1 min prior to FS).
Injection pipettes (tip diameter 12 – 20 µm) were fashioned from microdispensing capillary tubes as described previously (Akaoka and Aston-Jones, 1991; Shiekhattar et al., 1991). The thin lumen of these pipettes allowed accurate measurement of microinjections volume (accuracy within 15 nl). The shank of the pipette was bent and attached to the recording pipette, with its tip recessed approximately 100 µm, using UV light-curing dental resin (Filtek Supreme, Scotchbrand multi purpose adhesive, 3M, St. Paul, MN).
Microinfusions were made by pneumatic pressure through the infusion barrel of the compound recording/infusion microelectrode (Picospritzer III, Parker Instruments, Cleveland, OH). After isolating a single LC neuron, baseline firing frequencies were recorded for at least 150 sec. Following this baseline period, solutions were administered by applying pressure pulses of 30 – 100 msec duration at 40 psi every 15 seconds for a total of 150 – 225 sec. With this procedure, each pulse delivered approximately 12 nl and the total volume applied was 120 – 180 nl. Care was taken to insure that microinfusion did not interfere with unit recordings by constantly monitoring spike amplitude during delivery. Typically, neurons could be held long enough to observe firing frequencies return to baseline (washout) within 30 – 300 sec.
SB-334867-A (100 µM; Tocris, Ellisville MO) and orexin-A (100 µM; Tocris, Ellisville MO) were dissolved at their final concentrations in artificial cerebrospinal fluid (ACSF), consisting of (in mM): 112 NaCl, 3.1 Kcl, 1.2 MgSO4, 0.4 NaH2PO4, and 25 NaHCO3, and were either used fresh or stored at −20 °C for no longer than one week before use. The basis for choosing these concentrations was previous in vivo electrophysiological work (Yamuy et al., 2004) and personal communications with the authors.
At the end of recordings, micropipette penetrations were marked by iontophoretic ejection of dye from the recording pipette (−20 µA, 50 % duty cycle for 15 min). Brains were snap-frozen in a solution of isopentane at −80 °C and coronal sections through the LC (40 µm thick) were cut on a cryostat, mounted on gelatinized glass slides, and stained with neutral red.
Data analysis
Spike frequencies were averaged into 10-sec bins using recording software (SPIKE 2; Cambridge Electronic Design, Cambridge, England) and exported to an Excel spreadsheet. After stable recordings were established and before drug application, a 150 sec window of time was chosen to average baseline impulse activity and compare with average frequencies during drug application. Significance was tested by paired t-tests before and during drug application.
Fos and orexin immunohistochemistry
Immunohistochemistry for Fos and orexin were as recently described (Harris et al., 2005). Animals were euthanized for Fos studies at ZT 10.5–11.5, 12.5–13.5 or 15.5–16.5. These times are 1.5 hrs following the time periods of interest (ZT 9–10, 11–12, 14–15, see results) to allow for the accumulation of Fos protein. Rats were deeply anaesthetized with Nembutal (50 mg / kg), and transcardially perfused with 0.9 % saline followed by ice-cold 4 % paraformaldehyde in 0.1 M phosphate buffered saline (PBS), pH 7.4. The brains were removed and postfixed for 2 hours in 2 % paraformaldehyde. They were then transferred to a 20 % sucrose solution and stored at 4° C for 5 days.
Coronal sections (40 µm thick) that included the DMH, PeF and LH were cut on a cryostat (Leica CM3050, Wetzlar, Germany) and collected in 0.1 M PBS. Sections were incubated in PBS containing 0.1 % H2O2 for 15 min and then for two h in PBS containing Triton X-100 (0.3 %) in normal donkey serum (2 %) (PBST / NDS). Sections were subsequently incubated overnight at room temperature in rabbit anti-cFOS antibody (1:10,000, oncogene, Cambridge, MA) in the presence of 0.1 % Na-Azide (PBST-Az / NDS). Following this, slices were incubated in biotinylated donkey anti rabbit (1:500, Jackson ImmunoResearch, West Grove, PA) for two hours at room temperature in PBS-AZ / NDS. Tissue was transferred to an avidin-biotin complex (ABC, 1:500 in PBST; Vectastain ABC Elite kit; Vector Laboratories, Burlingame, CA) for 2 h at room temperature, rinsed once in 0.01 M PBS, then visualized by reaction with 0.02 % 3,3’-diaminobenzidine (DAB, Sigma), 0.01 % H2O2, and 0.6 % nickel ammonium sulfate in 0.05 M Tris buffer (pH 7.4) for 2 min. The reaction was terminated by transferring slices to 0.01 M PBS.
Next, these slices were incubated in goat anti-orexin-A (1:1000; Santa Cruz Biotechnology, Santa Cruz, CA) and subsequently in biotinylated donkey anti-goat (1:500, Jackson ImmunoResearch, West Grove, PA), and then processed for visualization s described above except without the nickel ammonium sulfate. Slices were mounted on microscope slides (Fisher Superfrost / Plus, Waltham, MA), dehydrated through graded alcohols, cleared in xylene, and coverslipped with Permount. .
Fos Counting and analysis
A single examiner, who was blinded to treatment conditions, performed all counts using a Leitz Laborlux microscope. Orexin-IR neurons and orexin-Fos double-labeled neurons were counted in caudal sections of the hypothalamus (3 – 4 per animal) containing the DMH, PeF and LH areas. DMH orexin neurons were counted using a vertically oriented 0.4 × 0.6 mm box, PeF orexin-IR neurons located dorsal to the fornix were counted using a horizontally oriented 1 × 0.4 mm box, and LH orexin-IR neurons a vertically oriented 0.7 × 1.0 mm box, analogous to Estabrooke et al. (2001). Criteria for DMH, PeF and DMH locations were approximately as previously described (Harris et al., 2005), and special care was taken to ensure that counting boxes did not overlap with each other. The percentage of orexin neurons that also stained for Fos for each animal was then calculated as a measure of orexin neuron activation.
Orexinergic afferents to the LC
For retrograde tracing experiments, 8 rats were anesthetized with halothane and the same stereotaxic coordinates and electrophysiological criteria as in the above “Electrophysiology” methods section were used to advance a glass micropipette (tip diameter 12 – 20 µm) to the LC. Over the course of 5 – 10 min, 40 – 60 nl CTb (0.05 % in 0.1 M phosphate buffer, pH 6) was injected unilaterally by brief pneumatic pulses from a controlled pressure source (Picrospritzer, Parker Instruments, Cleveland, OH). The pipette was left in place for 10 min thereafter to limit leakage of the tracer along the pipette track. Following post-operative procedures, rats survived 2 – 3 days before transcardial perfusion.
Perfusion and tissue slicing was performed as described above for Fos immunohistochemistry. CTb immunohistochemistry included an overnight incubation in goat anti-CTb antibody (1:20,000; List Biological Labs, Hornby, Canada) followed by incubation in secondary biotinylated donkey anti-goat IgG (1:1000; Jackson, ImmunoResearch, West Grove, PA) antibody for 90 min at room temperature. CTb was visualized using the ABC Elite kit (1:500; 90 min at room temperature) and the Blue substrate SG (Vector Laboratories, Burlingame, CA). Hypothalamic slices were subsequently stained for orexin as described above. Slices containing the LC were additionally stained for tyrosine hydroxylase (TH) using a monoclonal mouse anti-TH antibody (1:2000; Incstar, Stillwater, MN) primary antibody incubated overnight at room temperature and a biotinylated donkey anti-mouse IgG (1:500; Jackson, ImmunoResearch, West Grove, PA) secondary antibody incubated for 90 min at room temperature. TH was visualized using mouse PAP (1:250; Jackson, ImmunoResearch, West Grove, PA) followed by a DAB reaction.
Slices containing the LC were used to verify injection sites. Slices containing the DMH, PeF, and LH were used to count total the number of CTb positive neurons, orexin neurons, and neurons positive for both markers. The areas used for counting are similar to those described before (Harris et al., 2005), and are illustrated in Figure 5. In brief, LH was the area of orexin neurons lateral to the fornix, PeF consisted of orexin neurons dorsal to and extending ~400 um medial to the fornix, and DMH was defined as more medical orexin neurons, corresponding approximately to the DMH nucleus. Color images were taken on a digital camera and saved to disk for quantification using Openlab image processing software (Improvision, Ltd.; Coventry, UK) on a Macintosh computer.
Acknowledgements
We would like to thank Yan Zhu for technical assistance. This work was supported by PHS grants NS024698, DA017289 and MH14654.
Abbreviations
- LC
locus coeruleus
- DMH
dorsomedial hypothalamus
- LH
lateral hypothalamus
- PeF
Perifornical area
- SCN
suprachiasmatic nucleus
- FS
footpad stimulation
- ZT
Zeitgeber time
- PBS
phosphate buffered saline
- CTb
Cholera toxin b subunit
- TH
tyrosine hydroxylase
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Literature References
- Abrahamson EE, Leak RK, Moore RY. The suprachiasmatic nucleus projects to posterior hypothalamic arousal systems. Neuroreport. 2001;12:435–440. doi: 10.1097/00001756-200102120-00048. [DOI] [PubMed] [Google Scholar]
- Adamantis A, de Lecea L. Physiological arousal: a role for hypothalamic systems. Cell Mol Life Sci. 2008 doi: 10.1007/s00018-008-7521-8. E-pub. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akaoka H, Aston-Jones G. Opiate withdrawal-induced hyperactivity of locus coeruleus neurons is substantially mediated by augmented excitatory amino acid input. J Neurosci. 1991;11:3830–3839. doi: 10.1523/JNEUROSCI.11-12-03830.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aston-Jones G. Brain structures and receptors involved in alertness. Sleep Med. 2005;6 Suppl 1:S3–S7. doi: 10.1016/s1389-9457(05)80002-4. [DOI] [PubMed] [Google Scholar]
- Aston-Jones G, Bloom FE. Activity of norepinephrine-containing locus coeruleus neurons in behaving rats anticipates fluctuations in the sleep-waking cycle. J Neurosci. 1981;1:876–886. doi: 10.1523/JNEUROSCI.01-08-00876.1981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aston-Jones G, Chen S, Zhu Y, Oshinsky ML. A neural circuit for circadian regulation of arousal. Nat Neurosci. 2001;4:732–738. doi: 10.1038/89522. [DOI] [PubMed] [Google Scholar]
- Aston-Jones G, Zhu Y, Card JP. Numerous GABAergic afferents to locus ceruleus in the pericerulear dendritic zone: possible interneuronal pool. J Neurosci. 2004;24:2313–2321. doi: 10.1523/JNEUROSCI.5339-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bernard R, Lydic R, Baghdoyan HA. Hypocretin-1 activates G proteins in arousal-related brainstem nuclei of rat. Neuroreport. 2002;13:447–450. doi: 10.1097/00001756-200203250-00017. [DOI] [PubMed] [Google Scholar]
- Berridge CW, Waterhouse BD. The locus coeruleus-noradrenergic system:modulation of behavioral state and state-dependent cognitive processes. Brain Res Brain Res Rev. 2003;42:33–84. doi: 10.1016/s0165-0173(03)00143-7. [DOI] [PubMed] [Google Scholar]
- Bourgin P, Huitron-Resendiz S, Spier AD, Fabre V, Morte B, Criado JR, Sutcliffe JG, Henriksen SJ, de Lecea L. Hypocretin-1 modulates rapid eye movement sleep through activation of locus coeruleus neurons. J Neurosci. 2000;20:7760–7765. doi: 10.1523/JNEUROSCI.20-20-07760.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chou TC, Scammell TE, Gooley JJ, Gaus SE, Saper CB, Lu J. Critical role of dorsomedial hypothalamic nucleus in a wide range of behavioral circadian rhythms. J Neurosci. 2003;23:10691–10702. doi: 10.1523/JNEUROSCI.23-33-10691.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chou TC, Lee CE, Lu J, Elmquist JK, Hara J, Willie JT, Beuckmann CT, Chemelli RM, Sakurai T, Yanagisawa M, Saper CB, Scammell TE. Orexin (hypocretin) neurons contain dynorphin. J Neurosci. 2001;21:RC168. doi: 10.1523/JNEUROSCI.21-19-j0003.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Date Y, Ueta Y, Yamashita H, Yamaguchi H, Matsukura S, Kangawa K, Sakurai T, Yanagisawa M, Nakazato M. Orexins, orexigenic hypothalamic peptides, interact with autonomic, neuroendocrine and neuroregulatory systems. Proc Natl Acad Sci U S A. 1999;96:748–753. doi: 10.1073/pnas.96.2.748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deurveilher S, Semba K. Indirect projections from the suprachiasmatic nucleus to major arousal-promoting cell groups in rat: implications for the circadian control of behavioural state. Neuroscience. 2005;130:165–183. doi: 10.1016/j.neuroscience.2004.08.030. [DOI] [PubMed] [Google Scholar]
- Ennis M, Aston-Jones G. Activation of locus coeruleus from nucleus paragigantocellularis: a new excitatory amino acid pathway in brain. J Neurosci. 1988;8:3644–3657. doi: 10.1523/JNEUROSCI.08-10-03644.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ennis M, Aston-Jones G, Shiekhattar R. Activation of locus coeruleus neurons by nucleus paragigantocellularis or noxious sensory timulation is mediated by intraceorulear excitatory amino acid neurotransmission. Brain Res. 1992;598:185–195. doi: 10.1016/0006-8993(92)90182-9. [DOI] [PubMed] [Google Scholar]
- Espana RA, Reis KM, Valentino RJ, Berridge CW. Organization of hypocretin/orexin efferents to locus coeruleus and basal forebrain arousal-related structures. J Comp Neurol. 2005;481:160–178. doi: 10.1002/cne.20369. [DOI] [PubMed] [Google Scholar]
- Estabrooke IV, McCarthy MT, Ko E, Chou TC, Chemelli RM, Yanagisawa M, Saper CB, Scammell TE. Fos expression in orexin neurons varies with behavioral state. J Neurosci. 2001;21:1656–1662. doi: 10.1523/JNEUROSCI.21-05-01656.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gonzalez MM, Aston-Jones G. Circadian regulation of arousal: role of the noradrenergic locus coeruleus system and light exposure. Sleep. 2006;29:1327–1336. doi: 10.1093/sleep/29.10.1327. [DOI] [PubMed] [Google Scholar]
- Greco MA, Shiromani PJ. Hypocretin receptor protein and mRNA expression in the dorsolateral pons of rats. Brain Res Mol Brain Res. 2001;88:176–182. doi: 10.1016/s0169-328x(01)00039-0. [DOI] [PubMed] [Google Scholar]
- Hagan JJ, Leslie RA, Patel S, Evans ML, Wattam TA, Holmes S, Benham CD, Taylor SG, Routledge C, Hemmati P, Munton RP, Ashmeade TE, Shah AS, Hatcher JP, Hatcher PD, Jones DN, Smith MI, Piper DC, Hunter AJ, Porter RA, Upton N. Orexin A activates locus coeruleus cell firing and increases arousal in the rat. Proc Natl Acad Sci U S A. 1999;96:10911–10916. doi: 10.1073/pnas.96.19.10911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harris GC, Wimmer M, Aston-Jones G. A role for lateral hypothalamic orexin neurons in reward seeking. Nature. 2005;437:556–559. doi: 10.1038/nature04071. [DOI] [PubMed] [Google Scholar]
- Hervieu GJ, Cluderay JE, Harrison DC, Roberts JC, Leslie RA. Gene expression and protein distribution of the orexin-1 receptor in the rat brain and. Neuroscience. 2001;103:777–797. doi: 10.1016/s0306-4522(01)00033-1. [DOI] [PubMed] [Google Scholar]
- Hoang QV, Bajic D, Yanagisawa M, Nakajima S, Nakajima Y. Effects of orexin (hypocretin) on GIRK channels. J Neurophysiol. 2003;90:693–702. doi: 10.1152/jn.00001.2003. [DOI] [PubMed] [Google Scholar]
- Horvath TL, Peyron C, Diano S, Ivanov A, Aston-Jones G, Kilduff TS, van Den Pol AN. Hypocretin (orexin) activation and synaptic innervation of the locus coeruleus noradrenergic system. J Comp Neurol. 1999;415:145–159. [PubMed] [Google Scholar]
- Ivanov A, Aston-Jones G. Hypocretin/orexin depolarizes and decreases potassium conductance in locus coeruleus neurons. Neuroreport. 2000;11:1755–1758. doi: 10.1097/00001756-200006050-00031. [DOI] [PubMed] [Google Scholar]
- Jodo E, Aston-Jones G. Activation of locus coeruleus by prefrontal cortex is mediated by excitatory amino acid inputs. Brain Res. 1997;768:327–332. doi: 10.1016/s0006-8993(97)00703-8. [DOI] [PubMed] [Google Scholar]
- Lee MG, Hassani OK, Jones BE. Discharge of identified orexin/hypocretin neurons across the sleep-waking cycle. J Neurosci. 2005;25:6716–6720. doi: 10.1523/JNEUROSCI.1887-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin L, Wisor J, Shiba T, Taheri S, Yanai K, Wurts S, Lin X, Vitaterna M, Takahashi J, Lovenberg TW, Koehl M, Uhl G, Nishino S, Mignot E. Measurement of hypocretin/orexin content in the mouse brain using an enzymeimmunoassay: the effect of circadian time, age and genetic background. Peptides. 2002;23:2203–2211. doi: 10.1016/s0196-9781(02)00251-6. [DOI] [PubMed] [Google Scholar]
- Marcus JN, Aschkenasi CJ, Lee CE, Chemelli RM, Saper CB, Yanagisawa M, Elmquist JK. Differential expression of orexin receptors 1 and 2 in the rat brain. J Comp Neurol. 2001;435:6–25. doi: 10.1002/cne.1190. [DOI] [PubMed] [Google Scholar]
- Mileykovskiy BY, Kiyashchenko LI, Siegel JM. Behavioral correlates of activity in identified hypocretin/orexin neurons. Neuron. 2005;46:787–798. doi: 10.1016/j.neuron.2005.04.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nambu T, Sakurai T, Mizukami K, Hosoya Y, Yanagisawa M, Goto K. Distribution of orexin neurons in the adult rat brain. Brain Res. 1999;827:243–260. doi: 10.1016/s0006-8993(99)01336-0. [DOI] [PubMed] [Google Scholar]
- Pace-Schott EF, Hobson JA. The neurobiology of sleep: genetics, cellular physiology and subcortical networks. Nat Rev Neurosci. 2002;3:591–605. doi: 10.1038/nrn895. [DOI] [PubMed] [Google Scholar]
- Peyron C, Tighe DK, van den Pol AN, de Lecea L, Heller HC, Sutcliffe JG, Kilduff TS. Neurons containing hypocretin (orexin) project to multiple neuronal systems. J Neurosci. 1998;18:9996–10015. doi: 10.1523/JNEUROSCI.18-23-09996.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piper DC, Upton N, Smith MI, Hunter AJ. The novel brain neuropeptide, orexin-A, modulates the sleep-wake cycle of rats. Eur J Neurosci. 2000;12:726–730. doi: 10.1046/j.1460-9568.2000.00919.x. [DOI] [PubMed] [Google Scholar]
- Porter RA, Chan WN, Coulton S, Johns A, Hadley MS, Widdowson K, Jerman JC, Brough SJ, Coldwell M, Smart D, Jewitt F, Jeffrey P, Austin N. 1,3-Biarylureas as selective non-peptide antagonists of the orexin-1 receptor. Bioorg Med Chem Lett. 2001;11:1907–1910. doi: 10.1016/s0960-894x(01)00343-2. [DOI] [PubMed] [Google Scholar]
- Sakurai T. The neural circuit of orexin (hypocretin): maintaining sleep and wakefulness. Nat Rev Neurosci. 2007;8:171–181. doi: 10.1038/nrn2092. [DOI] [PubMed] [Google Scholar]
- Shiekhattar R, Aston-Jones G. NMDA-receptor-mediated sensory responses of brain noradrenergic neurons are suppressed by in vivo concentrations of extracellular magnesium. Synapse. 1992;10:103–109. doi: 10.1002/syn.890100204. [DOI] [PubMed] [Google Scholar]
- Shiekhattar R, Aston-Jones G, Chiang C. Local infusion of calcium-free solutions in vivo activates locus coeruleus neurons. Brain Res Bull. 1991;27:5–12. doi: 10.1016/0361-9230(91)90273-m. [DOI] [PubMed] [Google Scholar]
- Smart D, Sabido-David C, Brough SJ, Jewitt F, Johns A, Porter RA, Jerman JC. SB-334867-A: the first selective orexin-1 receptor antagonist. Br J Pharmacol. 2001;132:1179–1182. doi: 10.1038/sj.bjp.0703953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soffin EM, Evans ML, Gill CH, Harries MH, Benham CD, Davies CH. SB-334867-A antagonises orexin mediated excitation in the locus coeruleus. Neuropharmacology. 2002;42:127–133. doi: 10.1016/s0028-3908(01)00156-3. [DOI] [PubMed] [Google Scholar]
- Taheri S, Sunter D, Dakin C, Moyes S, Seal L, Gardiner J, Rossi M, Ghatei M, Bloom S. Diurnal variation in orexin A immunoreactivity and preproorexin mRNA in the rat central nervous system. Neurosci Lett. 2000;279:109–112. doi: 10.1016/s0304-3940(99)00955-6. [DOI] [PubMed] [Google Scholar]
- Trivedi P, Yu H, MacNeil DJ, Van der Ploeg LH, Guan XM. Distribution of orexin receptor mRNA in the rat brain. FEBS Lett. 1998;438:71–75. doi: 10.1016/s0014-5793(98)01266-6. [DOI] [PubMed] [Google Scholar]
- van den Pol AN, Ghosh PK, Liu RJ, Li Y, Aghajanian GK, Gao XB. Hypocretin (orexin) enhances neuron activity and cell synchrony in developing mouse GFP-expressing locus coeruleus. J Physiol. 2002;541:169–185. doi: 10.1113/jphysiol.2002.017426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walling SG, Nutt DJ, Lalies MD, Harley CW. Orexin-A infusion in the locus ceruleus triggers norepinephrine (NE) release and NE-induced long-term potentiation in the dentate gyrus. J Neurosci. 2004;24:7421–7426. doi: 10.1523/JNEUROSCI.1587-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamuy J, Fung SJ, Xi M, Chase MH. Hypocretinergic control of spinal cord motoneurons. J Neurosci. 2004;24:5336–5345. doi: 10.1523/JNEUROSCI.4812-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshida K, McCormack S, Espana RA, Crocker A, Scammell TE. Afferents to the orexin neurons of the rat brain. J Comp Neurol. 2006;494:845–861. doi: 10.1002/cne.20859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshida Y, Fujiki N, Nakajima T, Ripley B, Matsumura H, Yoneda H, Mignot E, Nishino S. Fluctuation of extracellular hypocretin-1 (orexin A) levels in the rat in relation to the light-dark cycle and sleep-wake activities. Eur J Neurosci. 2001;14:1075–1081. doi: 10.1046/j.0953-816x.2001.01725.x. [DOI] [PubMed] [Google Scholar]
- Zeitzer JM, Buckmaster CL, Parker KJ, Hauck CM, Lyons DM, Mignot E. Circadian and homeostatic regulation of hypocretin in a primate model: implications for the consolidation of wakefulness. J Neurosci. 2003;23:3555–3560. doi: 10.1523/JNEUROSCI.23-08-03555.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang JH, Sampogna S, Morales FR, Chase MH. Distribution of hypocretin (orexin) immunoreactivity in the feline pons and medulla. Brain Res. 2004a;995:205–217. doi: 10.1016/j.brainres.2003.10.004. [DOI] [PubMed] [Google Scholar]
- Zhang S, Zeitzer JM, Yoshida Y, Wisor JP, Nishino S, Edgar DM, Mignot E. Lesions of the suprachiasmatic nucleus eliminate the daily rhythm of hypocretin-1 release. Sleep. 2004b;27:619–627. doi: 10.1093/sleep/27.4.619. [DOI] [PubMed] [Google Scholar]