

Abstract
Multilocus sequence typing of 169 non-O157 enterohemorrhagic Escherichia coli (EHEC) isolated from patients with hemolytic uremic syndrome (HUS) demonstrated 29 different sequence types (STs); 78.1% of these strains clustered in 5 STs. From all STs and serotypes identified, we established a reference panel of EHEC associated with HUS (HUSEC collection).
Keywords: Enterohemorrhagic Escherichia coli, EHEC, hemolytic uremic syndrome, HUS, HUSEC, serotyping, molecular phylogeny, MLST, dispatch
Enterohemorrhagic Escherichia coli (EHEC) strains are a highly pathogenic subgroup of Shiga toxin–producing E. coli (STEC) that cause severe human diseases, including bloody diarrhea and hemolytic uremic syndrome (HUS) (1). The ability to cause severe human disease differentiates EHEC from other STEC found in the environment that are less pathogenic or nonpathogenic. E. coli O157:H7 is the most frequent EHEC implicated as a cause of HUS (2), but non–O157:H7 EHEC are variably present as the only pathogens in stools from HUS patients (1,3,4).
A recent phylogenetic analysis of E. coli isolated from humans and animals in different geographic areas by multilocus sequence typing (MLST), the current standard for phylogenetic analyses of bacteria, indicated extensive allelic variations and homolog recombinations in pathogenic lineages and demonstrated repeated and independent evolution of pathogenic strains (5,6). However, only a limited number of EHEC associated with HUS have been so investigated. Therefore, we performed a comprehensive MLST-based examination of the molecular phylogeny of EHEC isolated from HUS patients and established a collection of representative HUS-associated enterohemorrhagic E. coli (HUSEC) (www.ehec.org).
The Study
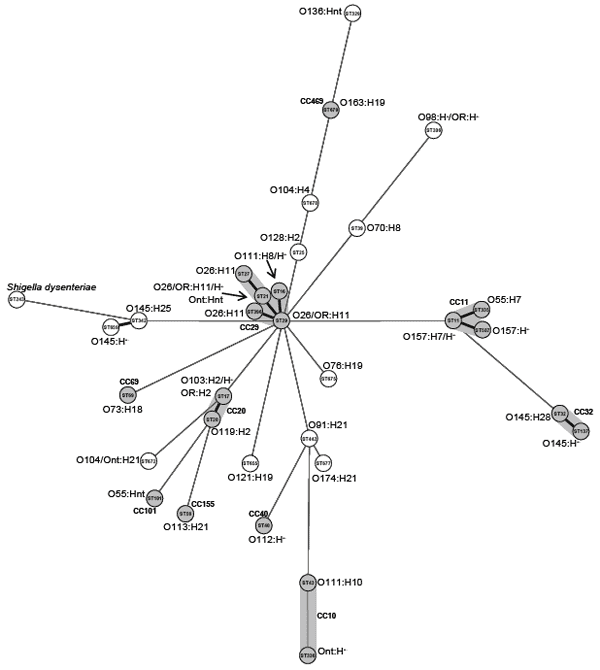
From 1996 through 2006, 524 EHEC were isolated as the only pathogens from fecal samples of epidemiologically unrelated patients with HUS (1 strain per patient). The isolation was achieved by using previously described procedures (7). The isolates were confirmed as E. coli by API 20 E (bioMérieux, Marcy l’Etoile, France) and serotyped (8) by using antisera against E. coli O antigens 1–181 and H antigens 1–56. In all nonmotile isolates from serogroups O26, O103, O111, O145, and O157, fliC genes were genotyped (9,10). MLST was performed as described previously (6) with small modifications (11). Phylogenetic analyses were based on allelic data that used the BURST algorithm (12) to achieve a more robust interpretation of the clustering and to reduce the influences by the effects of the recombination, which are widespread in E. coli (6). In addition, the stringent definition of clonal complexes (CCs), with which strains sharing at least 6 identical alleles are grouped into the same CC, was applied. The minimum spanning tree was generated from the allelic profiles by using Shigella dysenteriae strain M1354 (ST243, by using data from http://web.mpiib-berlin.mpg.de/mlst/dbs/Ecoli) as outgroup ( Appendix Figure).
Among 524 EHEC isolated from HUS patients, 355 (67.7%) belonged to serotypes O157:H7/H– (249 were non–sorbitol-fermenting EHEC O157:H7/H– and 106 were sorbitol-fermenting EHEC O157:H–), and 169 (32.3%) belonged to 34 non-O157 serotypes. Because the phylogeny of E. coli O157:H7/ H- has been extensively studied and is well established (5,13), we focused on MLST analysis of the 169 non-O157 strains, which represent all non-O157 EHEC serotypes from German HUS patients during the study period. We performed MLST analysis of only a subset of 10 O157 strains as controls.
MLST analysis of 169 non-O157 EHEC isolates distinguished 29 different sequence types (STs), which clustered into 10 CCs and 12 singletons (Table 1). The predominant ST was ST21, which consisted of 43 isolates (25.4% of non-O157 EHEC), followed by ST29 (30 isolates, 17.8%), ST32 (30 isolates, 17.8%), ST17 (15 isolates, 8.9%), and ST16 (14 isolates, 8.3%) (Table 1). These 5 STs included 78.1% of all HUS-associated non-O157 EHEC. The remaining 21.9% (n = 37) of the non-O157 EHEC strains belonged to 24 other STs that comprised only 1–3 strains; 14 of these STs were found only once (Table 1). Among the 10 CCs identified, CC29 was the most frequent. It comprised 89 strains of 5 STs, corresponding to 60.1% of 148 strains that could be assigned to a CC and to 52.7% of all non-O157 EHEC (Table 1). After CC29, CC32 and CC20 were the most frequently identified CCs (Table 1).
Table 1. Clonal complexes, sequence types, and serotypes of non-O157 EHEC from patients with hemolytic uremic syndrome*.
| CC | No. strains (%)† | ST | No. strains (%)† | Serotype‡ (no. strains)§ |
|---|---|---|---|---|
| 29 | 89 (52.7) | ST21 | 43 (25.4) | O26:H11/H– (fliCH11) (41), OR:H11 (1), Ont:Hnt (1) |
| ST29 | 30 (17.8) | O26:H11/H– (fliCH11) (29), OR:H11 (1) | ||
| ST27 | 1 (0.6) | O26:H11 | ||
| ST396 | 1 (0.6) | O26:H11 | ||
|
|
|
ST16 |
14 (8.3) |
O111:H8/H– (fliCH8) |
| 32 | 32 (18.9) | ST32 | 30 (17.8) | O145:H28/H– (fliCH28) |
|
|
|
ST137 |
2 (1.2) |
O145:H– (fliCH28) |
| 20 | 16 (9.5) | ST17 | 15 (8.9) | O103:H2/H– (fliCH2) (14), OR:H2 (1) |
|
|
|
ST20 |
1 (0.6) |
O119:H2 |
| 10 | 3 (1.8) | ST43 | 1 (0.6) | O111:H10 |
|
|
|
ST330 |
2 (1.2) |
Ont:H– |
| 11 |
3 (1.8) |
ST335 |
3 (1.8) |
O55:H7 |
| 40 |
1 (0.6) |
ST40 |
1 (0.6) |
O112:H– |
| 69 |
1 (0.6) |
ST69 |
1 (0.6) |
O73:H18 |
| 101 |
1 (0.6) |
ST101 |
1 (0.6) |
O55:Hnt |
| 155 |
1 (0.6) |
ST56 |
1 (0.6) |
O113:H21 |
| 469 |
1 (0.6) |
ST679 |
1 (0.6) |
O163:H19 |
| NA |
2 (1.2) |
ST25 |
2 (1.2) |
O128:H2 |
| NA |
2 (1.2) |
ST678 |
2 (1.2) |
O104:H4 |
| NA |
2 (1.2) |
ST655 |
2 (1.2) |
O121:H19 |
| NA |
1 (0.6) |
ST329 |
1 (0.6) |
O136:Hnt |
| NA | 3 (1.8) | ST342 | 2 (1.2) | O145:H25/H– (fliCH25) |
|
|
|
ST659¶ |
1 (0.6) |
O145:H– (fliCH25) |
| NA |
1 (0.6) |
ST677 |
1 (0.6) |
O174:H21 |
| NA |
1 (0.6) |
ST39 |
1 (0.6) |
O70:H8 |
| NA |
1 (0.6) |
ST675 |
1 (0.6) |
O76:H19 |
| NA |
3 (1.8) |
ST442 |
3 (1.8) |
O91:H21 |
| NA |
3 (1.8) |
ST306 |
3 (1.8) |
O98:H– (2), OR:H–(1) |
| NA | 2 (0.6) | ST672 | 2 (1.2) | O104:H21 (1), Ont:H21 (1) |
*CC, clonal complex; ST, sequence type; EHEC, enterohemorrhagic Escherichia coli; HUS, hemolytic uremic syndrome; NA, not assigned. †% of strains of a CC and ST among all 169 non-O157 EHEC isolated from HUS patients. ‡H–, nonmotile; OR, O rough (autoagglutinable strain); nt, not typeable by the E. coli O and H antisera used. §Number of strains of the serotype that belonged to the respective ST; if no number is given, all strains of the serotype belonged to the respective ST. ¶ST659 is a single-locus variant of ST342.
The predominant serotypes identified among the 169 non–O157 HUS-associated EHEC were O26:H11/H– (n = 72; 42.6%), O145:H28/H– (n = 32; 18.9%), O111:H8/H– (n = 14; 8.3%), and O103:H2/H– (n = 14; 8.3%). The nonmotile strains within these serogroups shared the H antigen–encoding fliC gene with the motile strains that expressed the respective H antigen (Table 1). These 8 serotypes together constituted 132 (78.1%) of the non-O157 EHEC associated with HUS, whereas the other 37 strains (21.9%) belonged to 26 different serotypes, 17 of which contained only a single isolate (Table 1).
The most frequent serotypes including O26:H11/H–, O103:H2/H–, O111:H8/H–, and O145:H28/H– clustered into the 5 most prevalent STs (Table 1). However, not all isolates of the same serotype always belonged to the same ST (Table 1). One example is serotype O26:H11/H– (fliCH11), which was the most common non–O157 EHEC associated with HUS and clustered into 4 STs as single-locus variants (Table 1). Each of four O rough (OR) strains (2 OR:H11, and 1 each OR:H2 and OR:H–), none of which could be successfully serotyped, was matched by its ST to an O typeable strain, indicating a recent conversion from the smooth to the rough strain form.
The relationships among members of the different STs and CCs are demonstrated in the Appendix Figure. Within the serogroup O111, 14 isolates belonging to serotypes O111:H8 and O111:H– (fliCH8) were ST16 (CC29). In contrast, the EHEC O111:H10 isolate with ST43 (CC10) shared none of the 7 MLST loci with the O111:H8/H– strains, indicating that EHEC O111 causing HUS originate from 2 different clonal sources. Similar differences were observed between EHEC O145:H25 (ST342)/O145:H– (fliCH25) (ST659) and O145:H28 (ST32). Whereas ST659 is a single-locus variant of ST342, both allelic profiles differ in all loci from ST32.
The combination of MLST analysis and serotyping enabled us to establish the HUSEC collection. This collection comprises 41 EHEC isolated from HUS patients in Germany, which includes all 36 EHEC serotypes (O157 and non-O157) isolated from HUS patients and all 31 STs identified within these serotypes (Table 2). The strains included in this HUSEC collection were reserotyped and characterized for their stx genotypes and the presence of the eae gene (Table 2). Phenotypic characteristics and additional properties such as putative virulence determinants are available at www.EHEC.org.
Table 2. Strains of the HUSEC collection representing all serotypes of HUS-associated EHEC strains isolated in Germany, 1996–2006*.
| Strain | Original | Year of isolation | Serotype | ST (CC) | eae | stx 1 | stx2† |
|---|---|---|---|---|---|---|---|
| HUSEC001 | 05-946 | 2005 | O111:H10 | 43 (10) | – | – | 2 |
| HUSEC002 | 5152/97 | 1997 | Ont:H– | 330 (10) | – | – | 2 |
| HUSEC003 | 6334/96 | 1996 | O157:H7 | 11 (11) | + | – | 2 |
| HUSEC004 | 3072/96 | 1996 | O157:H– | 11 (11) | + | – | 2 |
| HUSEC005 | 2907/97 | 1997 | O55:H7 | 335 (11) | + | – | 2 |
| HUSEC006 | 5376/99 | 1999 | O157:H– | 587 (11) | + | – | 2 |
| HUSEC007 | 7382/96 | 1996 | O103:H2 | 17 (20) | + | – | 2 |
| HUSEC008 | 2791/97 | 1997 | O103:H– | 17 (20) | + | – | 2 |
| HUSEC009 | 6833/96 | 1996 | OR:H2 | 17 (20) | + | – | 2 |
| HUSEC010 | 1805/00/A | 2000 | O119:H2 | 20 (20) | + | 1 | – |
| HUSEC011 | 2516/00 | 2000 | O111:H8 | 16 (29) | + | 1 | 2 |
| HUSEC012 | 6037/96 | 1996 | O111:H– | 16 (29) | + | 1 | 2 |
| HUSEC013 | 2245/98 | 1998 | O26:H11 | 21 (29) | + | 1 | – |
| HUSEC014 | 5080/97 | 1997 | O26:H– | 21 (29) | + | 1 | 2 |
| HUSEC015 | 126814/98 | 1998 | OR:H11 | 21 (29) | + | 1 | 2 |
| HUSEC016 | 5028/97 | 1997 | Ont:Hnt | 21 (29) | + | 1 | – |
| HUSEC017 | 3319/99 | 1999 | O26:H11 | 27 (29) | + | 1 | 2 |
| HUSEC018 | 1530/99 | 1999 | O26:H11 | 29 (29) | + | – | 2 |
| HUSEC019 | 1588/98 | 1998 | OR:H11 | 29 (29) | + | 1 | – |
| HUSEC020 | 3271/00 | 2000 | O26:H11 | 396 (29) | + | – | 2 |
| HUSEC021 | 0488/99 | 1999 | O145:H28 | 32 (32) | + | – | 2 |
| HUSEC022 | 4557/99 | 1999 | O145:H– | 137 (32) | + | – | 2 |
| HUSEC023 | 1169/97/1 | 1997 | O112:H– | 40 (40) | – | – | 2dact |
| HUSEC024 | 2996/96 | 1996 | O73:H18 | 69 (69) | – | – | 2dact |
| HUSEC025 | 06-05009 | 2006 | O55:Hnt | 101 (101) | – | 1 | – |
| HUSEC026 | 99-09355 | 1999 | O113:H21 | 56 (155) | – | – | 2dact |
| HUSEC027 | 03-07727 | 2003 | O163:H19 | 679 (469) | – | – | 2dact |
| HUSEC028 | 03-06687 | 2003 | O128:H2 | 25 (NA) | – | 1c | 2d |
| HUSEC029 | 4256/99 | 1999 | O70:H8 | 39 (NA) | + | – | 2 |
| HUSEC030 | 05-03519 | 2005 | O98:H– | 306 (NA) | – | 1 | – |
| HUSEC031 | 7792/96 | 1996 | OR:H– | 306 (NA) | + | 1 | – |
| HUSEC032 | 2441/98 | 1998 | O136:Hnt | 329 (NA) | – | 1c | 2 |
| HUSEC033 | 4392/97 | 1997 | O145:H25 | 342 (NA) | + | – | 2 |
| HUSEC034 | 3332/99 | 1999 | O91:H21 | 442 (NA) | – | 1 | 2+2dact |
| HUSEC035 | 1529/98 | 1998 | O121:H19 | 655 (NA) | + | – | 2 |
| HUSEC036 | 2839/98 | 1998 | O145:H– | 659 (NA) | + | 1 | 2c |
| HUSEC037 | 02-03885 | 2002 | O104:H21 | 672 (NA) | – | 1 | 2+2dact |
| HUSEC038 | 3356/97/B | 1997 | Ont:H21 | 672 (NA) | – | 1 | 2dact |
| HUSEC039 | 3651/96 | 1996 | O76:H19 | 675 (NA) | – | 1c | – |
| HUSEC040 | 220/00 | 2000 | O174:H21 | 677 (NA) | – | – | 2c |
| HUSEC041 | 01-09591 | 2001 | O104:H4 | 678 (NA) | – | – | 2 |
*HUSEC, hemolytic uremic syndrome–associated enterohemorrhagic Escherichia coli; EHEC, enterohemorrhagic E. coli. For each serotype, the multilocus sequence type (ST) and the corresponding clonal complex (CC) are given in accordance to the E. coli multilocus sequence typing website (http://web.mpiib-berlin.mpg.de/mlst/dbs/Ecoli). Furthermore, the presence (+, present; – absent) of the intimin gene (eae), the Shiga toxin gene (stx), and its subtype(s) are specified. nt, not typeable by the O and H antisera used; H–, nonmotile; OR, O rough (autoagglutinable strain); NA, not assigned. †2dact, stx2d-activatable.
Conclusions
Most (81.1%) of the non-O157 EHEC clustered into 3 CCs and belonged to a limited number of serotypes. These strains were recovered independently from different regions in Germany over an 11-year period. For the remaining strains, epidemiologic support is not as strong, and the clonal analysis demonstrated that their chromosomal backgrounds are highly divergent from those of CC29, CC32, and CC20. In 14 STs, we have only 1 isolate. In these cases, excluding concurrent or recent infection by E. coli O157 serologically is even more important. This exclusion was not always possible because patients’ serum for the investigation of immunoglobulin M anti-O157 lipopolysaccharide antibodies is frequently not available. However, at least some of these strains might represent emerging clones in the human population, such as O111:H10 (10), O113:H21 (14), and O121:H19 (15). Thus, strains of these serotypes included in our HUSEC collection can be used in future studies as a reference to compare EHEC isolated in other countries from HUS patients. This would allow timely discovery of the emergence of new non-O157 clones associated with HUS and the virulence traits that they contain (www.ehec.org).
Supplementary Material
Minimum spanning tree of hemolytic uremic syndrome–associated enterohemorrhagic Escherichia coli strains and Shigella dysenteriae M1354 (ST243, data from the E. coli multilocus sequence type [ST] website [http://web.mpiib-berlin.mpg.de/mlst/dbs/Ecoli]) as an outgroup generated from allelic profiles based on the eBURST algorithm (12). Each ST is represented by a circle named with its ST, and the corresponding serotypes are given (OR, O rough; H–, nonmotile; nt, not typeable with the E. coli O and H antisera used). Black lines connecting pairs of STs indicate that they share 6 (thick lines) or 5 (thin) alleles. Gray lines connecting pairs of STs of increasing line length indicate that the STs share <4 alleles. In addition, the STs and, if applicable, the connecting lines of a clonal complex are shaded in gray.
{kind=link}
Acknowledgment
We are grateful to Phillip I. Tarr for extensive discussions of the manuscript.
This study was supported by grants from the Federal Ministry of Education and Research Network Zoonoses and the Network European Research ERA–NET PathoGenoMics.
Biography
Dr Mellmann is a consultant for hygiene and infectious diseases at the University Hospital Münster. His professional interests include molecular identification and epidemiology of EHEC and other bacterial pathogens.
Footnotes
Suggested citation for this article: Mellmann A, Bielaszewska M, Köck R, Friedrich AW, Fruth A, Middendorf B, et al. Analysis of collection of hemolytic uremic syndrome–associated enterohemorrhagic Escherichia coli. Emerg Infect Dis [serial on the Internet]. 2008 Aug [date cited]. Available from http://www.cdc.gov/EID/content/14/8/1287.htm
References
- 1.Karch H, Tarr PI, Bielaszewska M. Enterohaemorrhagic Escherichia coli in human medicine. Int J Med Microbiol. 2005;295:405–18. 10.1016/j.ijmm.2005.06.009 [DOI] [PubMed] [Google Scholar]
- 2.Tarr PI, Gordon CA, Chandler WL. Shiga-toxin-producing Escherichia coli and haemolytic uraemic syndrome. Lancet. 2005;365:1073–86. [DOI] [PubMed] [Google Scholar]
- 3.Bonnet R, Souweine B, Gauthier G, Rich C, Livrelli V, Sirot J, et al. Non–O157:H7 Stx2–producing Escherichia coli strains associated with sporadic cases of hemolytic-uremic syndrome in adults. J Clin Microbiol. 1998;36:1777–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Brooks JT, Sowers EG, Wells JG, Greene KD, Griffin PM, Hoekstra RM, et al. Non-O157 Shiga toxin-producing Escherichia coli infections in the United States, 1983–2002. J Infect Dis. 2005;192:1422–9. 10.1086/466536 [DOI] [PubMed] [Google Scholar]
- 5.Reid SD, Herbelin CJ, Bumbaugh AC, Selander RK, Whittam TS. Parallel evolution of virulence in pathogenic Escherichia coli. Nature. 2000;406:64–7. 10.1038/35017546 [DOI] [PubMed] [Google Scholar]
- 6.Wirth T, Falush D, Lan R, Colles F, Mensa P, Wieler LH, et al. Sex and virulence in Escherichia coli: an evolutionary perspective. Mol Microbiol. 2006;60:1136–51. 10.1111/j.1365-2958.2006.05172.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Mellmann A, Bielaszewska M, Zimmerhackl LB, Prager R, Harmsen D, Tschäpe H, et al. Enterohemorrhagic Escherichia coli in human infection: in vivo evolution of a bacterial pathogen. Clin Infect Dis. 2005;41:785–92. 10.1086/432722 [DOI] [PubMed] [Google Scholar]
- 8.Prager R, Strutz U, Fruth A, Tschäpe H. Subtyping of pathogenic Escherichia coli strains using flagellar (H)-antigens: serotyping versus fliC polymorphisms. Int J Med Microbiol. 2003;292:477–86. 10.1078/1438-4221-00226 [DOI] [PubMed] [Google Scholar]
- 9.Sonntag AK, Prager R, Bielaszewska M, Zhang W, Fruth A, Tschäpe H, et al. Phenotypic and genotypic analyses of enterohemorrhagic Escherichia coli O145 strains from patients in Germany. J Clin Microbiol. 2004;42:954–62. 10.1128/JCM.42.3.954-962.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Zhang W, Mellmann A, Sonntag A, Wieler L, Bielaszewska M, Tschäpe H, et al. Structural and functional differences between disease-associated genes of enterohaemorrhagic Escherichia coli O111. Int J Med Microbiol. 2007;297:17–26. 10.1016/j.ijmm.2006.10.004 [DOI] [PubMed] [Google Scholar]
- 11.Bielaszewska M, Prager R, Köck R, Mellmann A, Zhang W, Tschäpe H, et al. Shiga toxin gene loss and transfer in vitro and in vivo during enterohemorrhagic Escherichia coli O26 infection in humans. Appl Environ Microbiol. 2007;73:3144–50. 10.1128/AEM.02937-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Feil EJ, Li BC, Aanensen DM, Hanage WP, Spratt BG. eBURST: inferring patterns of evolutionary descent among clusters of related bacterial genotypes from multilocus sequence typing data. J Bacteriol. 2004;186:1518–30. 10.1128/JB.186.5.1518-1530.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Feng PCH, Monday SR, Lacher DW, Allison L, Siitonen A, Keys C, et al. Genetic diversity among clonal lineages within Escherichia coli O157:H7 stepwise evolutionary model. Emerg Infect Dis. 2007;13:1701–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Paton AW, Woodrow MC, Doyle RM, Lanser JA, Paton JC. Molecular characterization of a Shiga toxigenic Escherichia coli O113:H21 strain lacking eae responsible for a cluster of cases of hemolytic-uremic syndrome. J Clin Microbiol. 1999;37:3357–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.McCarthy TA, Barrett NL, Hadler JL, Salsbury B, Howard RT, Dingman DW, et al. Hemolytic-uremic syndrome and Escherichia coli O121 at a lake in Connecticut, 1999. Pediatrics. 2001;108:E59. 10.1542/peds.108.4.e59 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Minimum spanning tree of hemolytic uremic syndrome–associated enterohemorrhagic Escherichia coli strains and Shigella dysenteriae M1354 (ST243, data from the E. coli multilocus sequence type [ST] website [http://web.mpiib-berlin.mpg.de/mlst/dbs/Ecoli]) as an outgroup generated from allelic profiles based on the eBURST algorithm (12). Each ST is represented by a circle named with its ST, and the corresponding serotypes are given (OR, O rough; H–, nonmotile; nt, not typeable with the E. coli O and H antisera used). Black lines connecting pairs of STs indicate that they share 6 (thick lines) or 5 (thin) alleles. Gray lines connecting pairs of STs of increasing line length indicate that the STs share <4 alleles. In addition, the STs and, if applicable, the connecting lines of a clonal complex are shaded in gray.