

Abstract
This is an author-produced version of a manuscript accepted for publication in The Journal of Immunology (The JI). The American Association of Immunologists, Inc. (AAI), publisher of The JI, holds the copyright to this manuscript. This version of the manuscript has not yet been copyedited or subjected to editorial proofreading by The JI; hence, it may differ from the final version published in The JI (online and in print). AAI (The JI) is not liable for errors or omissions in this author-produced version of the manuscript or in any version derived from it by the U.S. National Institutes of Health or any other third party. The final, citable version of record can be found at www.jimmunol.org.
B lymphocyte stimulator (BLyS) is a well known direct costimulator of adaptive immune cells, particularly B lineage cells. However, we have reported recently that BLyS is also able to activate monocytes. Other innate immune cells such as dendritic cells (DCs), play a key role in the initiation of adaptive immune responses and the purpose of the current study was to assess whether there is a direct role for BLyS in modulating human DC functions. Here we show that BLyS induces DC activation and maturation. Thus, BLyS strongly induced upregulation of surface costimulatory molecule expression and secretion of specific cytokines and chemokines in DCs. BLyS-stimulated DCs (BLyS-DCs) were also able to augment allogeneic CD4 T cell proliferation to a greater extent than were control DCs. BLyS-DCs secreted elevated levels of the major Th1 polarizing cytokine, IL-12p70, and they promoted naïve CD4 T cell differentiation into Th1 T cells. Regarding BLyS receptor expression, DCs primarily express cytoplasmic TACI, however low levels of cell surface TACI are expressed as well. Collectively, our data suggest that BLyS may modulate adaptive immune cells indirectly by inducing DC maturation.
Keywords: Human, Dendritic Cells, Inflammation, Cell Activation, BLyS, TACI
Introduction
Dendritic cells (DCs) are professional antigen-presenting cells (APCs). Whereas immature DCs capture and process antigens in the peripheral tissues, mature DCs are specialized to present processed antigens to naïve T cells and promote their differentiation into Th1, Th2, or Th17 cells in lymphoid tissues (1-4). DCs initiate their maturation process when they encounter antigens through pattern recognition receptors, such as Toll like receptors, and/or inflammatory stimuli in the periphery. During the process of maturation, DCs lose endocytic activity, but acquire high expression levels of cell surface major histocompatibility complex (MHC) antigens and costimulatory molecules, such as CD80, CD86, and CD40, thereby allowing efficient antigen presentation. DC maturation is accompanied by expression of the chemokine receptor CCR7, which permits DC migration from the periphery to the lymph nodes. DCs also play a role in the induction of immune tolerance by presenting self-antigen to T cells without increased expression of costimulatory molecules, secreting suppressive cytokines, or inducing regulatory T cells (5-9). In certain circumstances dysregulation of DC function is often related to autoimmunity (10, 11).
B lymphocyte stimulator (BLyS) is a TNF family member, and is also named BAFF, zTNF4, THANK, and Tall-1(12, 13). BLyS is produced by myeloid lineage cells, such as monocytes, macrophages, dendritic cells, and neutrophils; malignant B cells; astrocytes; airway epithelial cells; and bone marrow stromal cells (12-20). IFN-γ, IFN-α, or IL-10 increases BLyS production by myeloid cells. Bacterial components like LPS and peptidoglycan can also up-regulate BLyS secretion by macrophages, dendritic cells and monocytes (14, 21). BLyS has three receptors: BCMA (B cell maturation antigen), TACI (transmembrane activator and CAML-interactor) and BAFF-R (BAFF receptor) and these receptors are primarily expressed in B lineage cells (22-26). TACI and BAFF-R have been shown to be expressed in a subset of T cells (25, 27-29). To date, most studies have focused on defining the biological effects of BLyS on B cells and T cells and these studies demonstrate that BLyS functions as a survival or costimulatory factor in lymphocytes (30-32). Recently, we reported that BLyS induces monocyte survival and activation, and we showed that TACI is expressed intracellularly in monocytes and also expressed on the cell surface upon activation (33).
BLyS levels have been shown to be elevated in sera and certain inflamed tissues of autoimmune diseases, such as Sjogren's syndrome (SS), systemic lupus erythematosus (SLE), and rheumatoid arthritis (RA) (34-38). Consistent results have also been obtained using various murine autoimmune disease models (39, 40). Although elevated BLyS levels are known to promote B cell survival and hence autoantibody production (41), there may be other cell types that are also modulated by BLyS in either a direct or indirect manner. In this regard, it is interesting to note that the role of DCs in autoimmune diseases such as RA, SS, or SLE has been studied, and considerable evidence suggests that altered DC function is related to the initiation or perpetuation of those diseases (42-46). These observations, taken together with our recent study demonstrating the ability of BLyS to directly activate monocytes, support the hypothesis that BLyS may play a larger role in coordinating the interplay between the innate and adaptive immune systems. Therefore, in this study we investigated the effects of BLyS on human DCs, and we show for the first time that BLyS directly promotes DC activation and maturation, which subsequently induces naïve CD4 T cell proliferation and differentiation into Th1 T cells. We also present our data concerning BLyS-binding receptor expression and demonstrate a role for TACI in mediating the direct effects of BLyS on DCs. Of interest, we demonstrate the existence of a unique intracellular pool of preformed TACI in monocyte-derived DCs, and present evidence that this pool is located within the Golgi apparatus. Thus, our data extend our understanding of the cellular targets of BLyS and suggest one more mechanism by which BLyS may play a role in modulating the adaptive immune system, i.e., indirectly through DC maturation.
Materials and Methods
Generation of human monocyte-derived DCs
Mononuclear cells (MNCs) were isolated from peripheral blood of healthy donors by Ficoll-Paque density gradient centrifugation. All patients provided written informed consent in accordance with the Declaration of Helsinki. The Mayo Clinic Rochester Institutional Review Board approved the protocol to obtain blood from volunteers. Monocytes were purified from MNCs by positive selection using the StemSep Ab enrichment cocktail (StemCell Technologies, Vancouver, BC) in combination with StemSep magnetic colloid, according to the manufacturer's instructions. Cell purity was assessed using flow cytometry and monocyte preparations were routinely >98% CD14 positive. Blood DCs were isolated from MNCs by positive selection using the blood dendritic cell isolation kit II from Miltenyi Biotech (Auburn, CA). To generate DCs, purified CD14 positive monocytes (1 × 106/ml in 3ml) were cultured for 5 days in complete RPMI medium containing 1500 U/ml rhGM-CSF (Berlex, Inc., Wayne, NJ) and 25 ng/ml rhIL-4 (Peprotech, Rocky Hill, NJ) in 6 well plates (Becton Dickinson, San Jose, CA). rhGM-CSF (1500 U/ml) and IL-4 (25 ng/ml) were added at day 0 and day 2. On day 5, cells were harvested and washed twice before further stimulation. Cell purity was again assessed and cells were found to be >99% CD11c positive and there were undetectable CD3 and CD19 positive cells. For those experiments assessing the effects of BLyS on cell morphology, culture conditions included GM-CSF and IL-4. In all other experiments, cells were stimulated with or without recombinant human BLyS (Calbiochem, La Jolla, CA), LPS (055:B5, Sigma-Aldrich, St. Louis, MO), or CD40L (Alexis Biochemicals, San Diego, CA) in the absence of GM-CSF and IL-4.
FACS analysis
Cells to be analyzed by flow cytometry were washed with PBS supplemented with 2% FBS containing 2 mM EDTA and 0.05% sodium azide. In all cases, cells were also stained with a matching isotype or fluorescent control. For detecting BLyS binding receptors, cells were incubated with biotinylated goat anti-TACI and anti-BCMA (R&D Systems, Inc., Minneapolis, MN) or PE-anti-BAFF-R (eBioscience, San Diego, CA) for 30 min at 4°C. After washing, PE-conjugated streptavidin (Caltag, Burlingame, CA) was added and cells were incubated for an additional 15 min at 4°C, and washed again. For BAFF-R staining, cells were fixed after washing without further secondary antibody staining. After washing, cells were fixed with 1% paraformaldehyde and analyzed. To detect intracellular BLyS binding receptors, cells were first fixed and permeabilized using a fixation/permeabilization kit purchased from Caltag, and then cells were stained. For BLyS binding on DCs, cells were incubated with or without 2 μg/ml BLyS for 2h at 37°C in FACS buffer, washed, and stained with biotinylated anti-BLyS (R&D Systems, Inc.) followed by PE-conjugated streptavidin. To determine the ability of BLyS to induce DC maturation markers, DCs were stimulated with or without BLyS or LPS for 24h, and cells were incubated with PE-conjugated anti-CD83, anti-CD80, anti-CD86 (BD Biosciences, San Jose, CA) or FITC-conjugated anti-DC-SIGN (AbD Serotec, Oxford, UK). Mannose receptor and CD40 were detected with unconjugated murine antibodies (AbD Serotec, Oxford, UK and Invitrogen, Carlsbad, CA, respectively) followed by PE-conjugated anti-mouse Ig (Invitrogen, Carlsbad, CA). In some experiments, BLyS or LPS were boiled for 20 min or pretreated with polymyxin B (PB) (20 µg/ml, for 20 min at room temperature) before incubating with cells. For all flow cytometric analyses, we used the FcγR blocking reagent (Miltenyi Biotech) to pretreat cells or it was included during the primary antibody incubation in DCs. Data were analyzed using FlowJo software (Tree Star, Inc., Ashland, OR).
Endocytosis
Unstimulated or stimulated DCs (1 × 105) were suspended in 100 μl of complete medium. Cells were incubated with 1 mg/ml of FITC-conjugated dextran beads (40,000 MW, Molecular Probes, Eugene, OR) for 1.5 h at 37 °C or on ice to measure nonspecific binding. Cells were washed three times, fixed with 1% paraformaldehyde, and analyzed by flow cytometry.
CD4 T cell proliferation
DCs were collected after a 24h incubation with different stimuli, washed twice to remove cytokines, irradiated (2000 rad), and were then seeded in round-bottomed 96-well plates (Corning, Corning, NY) at 10,000 to 625 cells/well (serial two-fold dilutions). Naïve CD45RA+ CD45RO− CD4 T cells were enriched to >98% purity by magnetic cell separation using StemCell Technology naïve CD4 T cell enrichment cocktail /EasySep® Magnetic Nanoparticles and the negative selection program on the Robosep Separator (StemCell Technologies). A constant number of CD4 T cells (1× 105/200μl) were added to each well and cultured for 3 days. Cells were pulsed with 1 μCi tritiated thymidine (3H-TdR; 5.0 Ci/mmol [185 GBq/mmol], Amersham, Piscataway, NJ) for the final 16h before collecting and counting radioactivity. 3H-TdR incorporation levels were determined using a Beckman scintillation counter.
Naïve CD4 T cell polarization
For detecting cytokine secretion by CD4 T cells, 2.5×105 naïve CD4 T cells were co-cultured with 0.5×105 preactivated DCs in 200 μl of complete medium in round-bottomed 96 well plates (DC:T cells = 1:5) for 48 h. Th1 or Th2 cytokines were determined using a cytometric bead assay (CBA) kit (BD Biosciences) according to the manufacturer's instructions. Data analysis was performed using BD Biosciences CBA software. Cell free supernatants were stored at −80 °C until they were analyzed. For intracellular staining, 1×105 naïve CD4 T cells were co-cultured with 0.2 ×105preactivated DCs in flat-bottomed 96 well plates (DC:T cells = 1:5). On day 5, cells were stimulated with 50 ng/ml PMA plus 1 μg/ml ionomycin in the presence of 10 μg/ml brefeldin A (Sigma-Aldrich) for the last 4 h of culture. Cells were fixed and permeabilized using a fixation/permeabilization kit (Caltag). Cells were stained with BD FastImmune™ IFN-γ/IL-4 (BD Biosciences) and APC-CD3 (BD Biosciences) in permeabilization solution. CD3 positive cells were analyzed for levels of cytoplasmic cytokine expression. FcγR blocking reagent (Miltenyi Biotech) was used in all staining procedures.
Human cytokine array
The profile of cytokines secreted by DCs (2×106/2ml in 24 well plates) was determined by using a protein microarray (RayBio Human Cytokine Ab Array V; RayBiotech, Inc., Norcross,GA). After a 24h incubation with or without stimuli, cell culture supernatants were harvested and stored at −80 °C until they were analyzed. Experiments were performed according to the manufacturer's instructions.
Quantification of cytokine secretion by ELISA
IL-6, IL-1β, IL-23, TNF-α, IL-10, (R&D Systems, Inc.), and IL-12p70 (BD Bioscience) levels were determined in cell-free culture supernatants using cytokine-specific ELISA kits. Supernatants of dendritic cell cultures were stored at −80 °C until they were analyzed. Results are shown in picograms per milliliter or nanograms per milliliter (as mean +/- SD).
Immunofluorescence analysis
In vitro generated DCs were placed into 12×75 mm FACS tubes and fixed with 4% paraformaldehyde for 20 min at room temperature (RT). After washing, cells were permeabilized with 0.1% Triton X-100 for 5 min at RT and washed with 3ml FACS buffer. After blocking with 5% normal rabbit serum in PBS (NRS/PBS) for 1 h at RT, cells were preincubated with FcγR blocking reagent (Miltenyi Biotech) in 5% NRS/PBS for 10 min. Goat anti-TACI antibody (C-20, Santacruz Biotechnology, Santa Cruz, CA) and mouse GM130 antibody (BD Bioscience) were then added and cells were incubated for an additional 1h at RT. After washing with 3ml FACS buffer, cells were stained with FITC labeled F(ab')2 rabbit anti-goat secondary antibody and Cy3 labeled F(ab')2 rabbit anti-mouse antibody (Jackson ImmunoResearch Laboratories, Inc., West Grove, PA) for 40 min at RT. Cells were then washed with 4 ml FACS buffer and fixed again with 4% paraformaldehyde for 20 min at RT. Cytospin preparations of the cells on glass slides were made using a Thermo Shandon cytospin 2 (350 rpm, 4 min). When slides were dry, a mounting solution containing DAPI (Vector Laboratories, Inc., Burlingame, CA) was applied, and images were taken using a Zeiss LSM 510 Confocal Laser Scanning Microscope (Carl Zeiss, Oberkochen, Germany).
Statistical analysis
Statistical analysis was performed using the Student's t test. Values of p < 0.05 were considered to be significant.
Results
BLyS induces DC maturation
We first determined whether BLyS has any biological effect on DCs. To accomplish this goal, we generated monocyte-derived DCs using an in vitrosystem and DCs generated in this manner were then cultured with or without BLyS or LPS. When DC cultures were analyzed by light microscopy, BLyS was observed to induce significant cell clustering (Fig. 1; bottom right panel). In addition, BLyS-stimulated DCs (BLyS-DCs) were larger than unstimulated cells and displayed more dendrites (Fig. 1; bottom left panel). To further characterize the activation of DCs by BLyS, cell surface phenotype was characterized using flow cytometry following incubation of DCs with BLyS or LPS as a control. As shown in Fig. 2A, surface expression of the costimulatory molecules, CD80, CD86, and the maturation marker, CD83, greatly increased upon stimulation with BLyS or LPS compared to unstimulated cells. BLyS also strongly induced CD40 expression, however, BLyS down regulated expression of the mannose receptor and DC-SIGN (Fig. 2B). Figure 2C demonstrates that the ability of BLyS to significantly increase CD80 and CD86 levels and induce CD83 is reproducible across multiple, independent experiments, and that it occurred in a dose dependent manner (Fig. 2D). In data not shown, HLA-DR expression was not significantly changed by BLyS stimulation. The DC maturation process is also accompanied by a decrease in phagocytic activity. We next assessed the ability of BLyS to attenuate phagocytosis by measuring DC uptake of FITC-conjugated dextran. Compared to unstimulated DCs, BLyS-DCs showed significantly reduced phagocytosis (Fig. 3). Finally, to confirm the effects of BLyS on DCs, we also analyzed directly isolated DCs from peripheral blood. Following isolation, purified blood DCs were cultured with BLyS. As shown in Fig. 4, BLyS also strongly induced CD83 expression on CD11c positive myeloid DCs.
FIGURE 1. BLyS induces DC morphological changes.
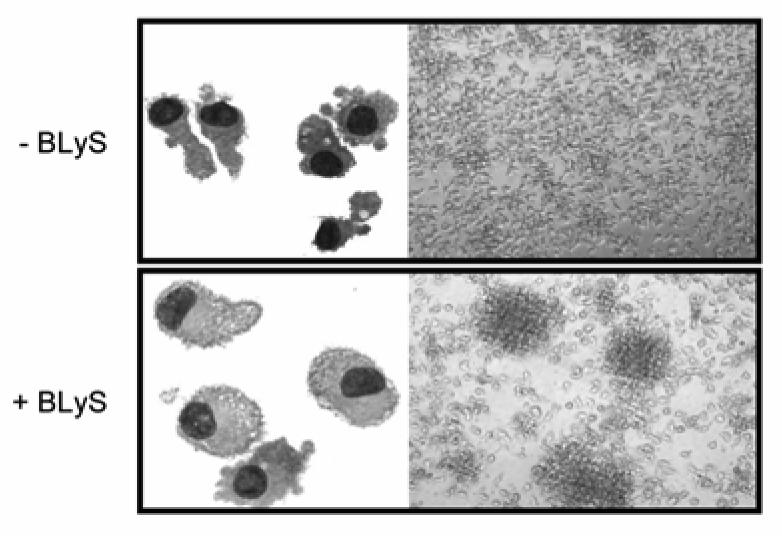
Cell morphology was analyzed by Wright-Giemsa staining (left) or light microscopy (right) after a 48h stimulation with or without BLyS in the presence of GM-CSF/ IL-4.
FIGURE 2. BLyS promotes DC maturation.

A and B. DCs (1×106/ml) were cultured with or without 200 ng/ml BLyS or 20 ng/ml LPS for 24 h, before analysis of expression of the indicated surface molecules on viable cells using flow cytometry. The experiments shown in A and B are representative of 9 and 4 independent experiments, respectively. ΔMFI (delta mean fluorescence intensity), is calculated from the MFI of the cells expressing the marker of interest divided by the MFI of the cells stained with the isotype control, and these numbers are shown in each histogram. C. The fold induction of ΔMFI for CD80, CD86, and CD83 in BLyS or LPS stimulated-DCs as compared with unstimulated DCs is displayed from nine independent experiments. When compared with Nil, *p=0.0002, **p=0.0063, and ***p=0.0022. D. BLyS was titrated over the indicated range of concentrations, and cell surface expression of CD80, as determined by flow cytometry, was determined following a 24 hour stimulation period. The data shown reflect the median CD80 fluorescence intensity observed in a single experiment and are representative of three independent experiments.
FIGURE 3. The phagocytic activity is decreased in BLyS-treated DCs.
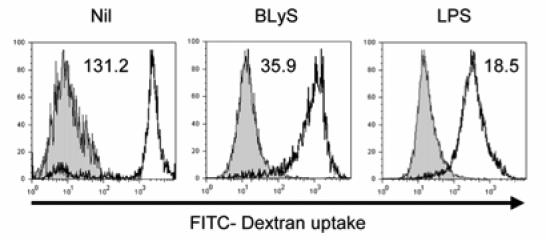
After cells were incubated with FITC-dextran for 2 h at 4°C (filled histogram, control for nonspecific binding) or at 37°C (open histogram), the levels of FITC-dextran uptake were examined by flow cytometry and the ΔMFI was calculated and is indicated on the graphs. The data shown are representative of five independent experiments.
FIGURE 4. BLyS promotes blood DC maturation.
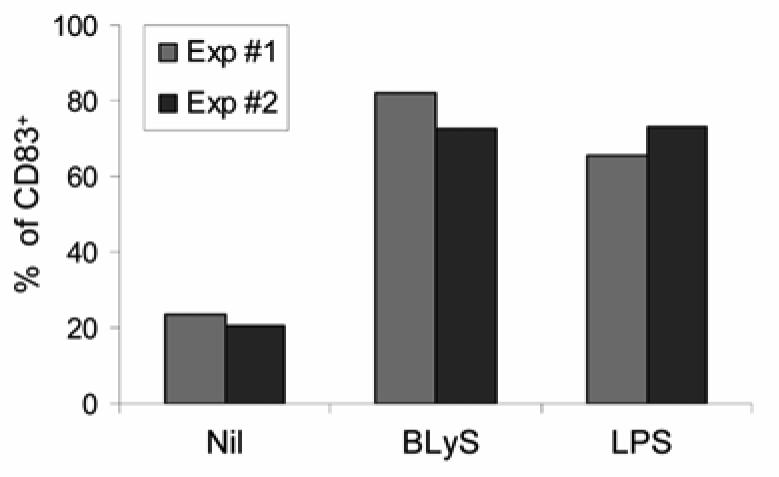
Freshly isolated blood DCs were cultured with or without 200 ng/ml BLyS or 20 ng/ml LPS for 48 h, and CD83 expression was analyzed by flow cytometry. The graph depicts the percentages of CD83-expressing cells in the CD11c positive population observed following stimulation in two independent experiments (Exp #1 and Exp #2).
BLyS stimulates DCs to secrete cytokines and chemokines
To further characterize the effects of BLyS on DCs, we used a cytokine/chemokine antibody-based array to measure protein secretion by DCs cultured in the presence or absence of BLyS or LPS. BLyS induced secretion of a variety of chemokines and cytokines to varying degrees, including IL-6, IL-8, GRO-α, IL-1α, IL-1β, TNF-α, monocyte chemoattractant protein 1 (MCP-1), MCP-2, MCP-4, macrophage inflammatory protein 1β (MIP-1β), MIP-3α, and RANTES (Fig. 5B). Compared with unstimulated cells, secretion of IL-6, IL-8, MCP-1, and growth-related oncogene (GRO) was highly induced by BLyS (Fig. 5B). MCP-1 and RANTES secretion was notable in that greater induction was observed in the BLyS-stimulated cells than in the LPS-stimulated positive control cultures. To a lesser degree, MCP-2, MCP-4, MIP-3α, and MIP-1β were also increased by BLyS stimulation. Of interest, BLyS-treated DCs produced high amounts of the proinflammatory cytokines IL-6, IL-1α, IL-1β, and the Th1 chemokine, IP-10, but not the chemokines that recruit Th2 type immune cells, such as TARC, MDC, and I-309 (Fig. 5B).
FIGURE 5. BLyS induces DC cytokine and chemokine production.

A. Cytokine array map. B. Cytokine production by BLyS-DCs (200 ng/ml, 24 h) or LPS-DCs (20 ng/ml, 24 h) was compared with unstimulated DCs by using the RayBio Human Cytokine Ab Array IV. One representative out of two independent experiments is shown. C. After stimulating cells with or without 200 ng/ml BLyS or 20 ng/ml LPS for 24 h, cell free supernatants were harvested to determine the levels of IL-6, IL-1β, and TNF-α by ELISA. Data reflect the mean +/- SD of supernatants collected from three independent experiments.
The enhanced production of IL-6, IL-1β, and TNF-α by BLyS was confirmed by direct ELISA assay. Particularly, IL-6 and TNF-α production was greatly enhanced and IL-1β was moderately increased by BLyS (Fig. 5C). Because IL-6 and IL-1β, in concert with IL-23, have been shown to induce differentiation of Th17 cells (47, 48), we next examined the ability of BLyS to induce IL-23 secretion. As shown in Table 1, we observed that BLyS stimulation resulted in only very low levels of IL-23 secretion as revealed by ELISA, and it was quite variable between donors. Despite the variable and low levels of IL-23 secretion, we did investigate whether BLyS stimulation induced the appearance of Th17 cells, characterized by the intracytoplasmic expression of IL-17, However, in results not shown, we failed to detect cells expressing intracellular IL-17 following BLyS stimulation.
Table 1.
Effects of BLyS on DC IL-23 secretion
| Experiment Number | |||||
|---|---|---|---|---|---|
| Stimulus | 1 | 2 |
3 pg/mL |
4 | 5 |
| Nil | ND* | ND | ND | ND | ND |
| BLyS | 219.5 | ND | 18.8 | 59.7 | ND |
| LPS | 270.4 | 1234.8 | 55.2 | 202.9 | 376.4 |
| CD40L | 65.6 | 18.8 | ND | ND | 34.7 |
After stimulating cells with or without 200 ng/ml BLyS or 20 ng/ml LPS for 24 h, cell free supernatants were harvested to determine the levels of IL-23 by ELISA. ND: below detection level (<15 pg/mL).
BLyS-DCs enhance allogeneic CD4 T cell proliferation
The results described above clearly demonstrate that BLyS induces DC maturation. To address the functional activity of BLyS-DCs, we next performed allogeneic mixed leukocyte reaction (MLR). BLyS-DCs were comparable to LPS-DCs in their ability to strongly augment CD4 T cell proliferation (Fig. 6). DCs stimulated with 100 ng/ml CD40L had only a moderate effect on CD4 T cell proliferation (Fig. 6; however, when a higher concentration of CD40L was used (200 ng/ml), the ability of DCs to enhance CD4 T cell proliferation was comparable to BLyS-DCs (data not shown).
FIGURE 6. BLyS-treated DCs enhance T cell proliferation in an allogeneic MLR.
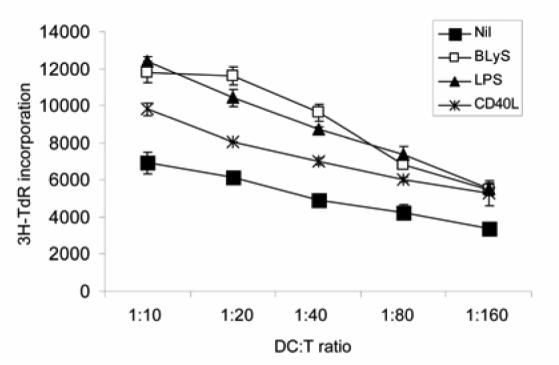
After a 24 h stimulation with or without 200 ng/ml BLyS, 100 ng/ml CD40L, or 20 ng/ml LPS, DCs were cocultured with 1×105 freshly isolated allogeneic naive CD4 T cells for 3 days in round-bottomed 96 well plates. Cells were pulsed with 1 µCi [3H]thymidine for the final 16 h, and then [3H]thymidine uptake was measured to determine CD4 T cell proliferation. Data reflect the mean +/- SD of triplicates from one experiment and are representative of six independent experiments.
BLyS-DCs induce Th1 immune responses
Next, we questioned whether BLyS-DCs are involved in naïve CD4 T cell differentiation. To address the question, we first determined whether BLyS induces DCs to produce IL-12, a well known Th1-driving cytokine. Although BLyS-DCs did not appear to secrete IL-12 as assessed using the cytokine array (Fig. 5B), the sensitivity and sample size of this assay are limited. We next used a more sensitive ELISA that would allow us to directly measure IL-12p70, the active form of IL-12. Even though the levels of IL-12 were variable among 10 independent experiments, IL-12p70 was clearly induced by BLyS with at least a two fold induction observed when compared with cells cultured with media alone (Nil; Fig. 7A). However, BLyS failed to induce IL-10 secretion in four independent experiments (Fig. 7B).
FIGURE 7. BLyS enhances DC IL-12 production, but not IL-10.

Immature DCs were incubated with 200 ng/ml BLyS, 20 ng/ml LPS, or without any stimulus for 24 h, and then supernatants were collected to measure IL-12p70 (A) and IL-10 (B) by ELISA. * When compared with Nil, p=0.014.
To determine whether BLyS-DCs induce differentiation of naïve CD4 T cells into Th1 CD4 T cells, we isolated human naïve CD45RA+CD45RO− CD4 T cells from peripheral blood and co-cultured them with stimulated or unstimulated DCs. After 48 h, supernatants were harvested and analyzed using a cytometric bead array. BLyS-DCs induced CD4 T cells to produce large amounts of the Th1 cytokine IFN-γ (Fig. 8A), but not Th2 type cytokines, such as IL-4, IL-5, and IL-10 (Fig. 8B). T cell cytokine production induced by LPS-DCs and CD40L-DCs was very similar (Fig. 8A and 8B), with the exception of IL-10 secretion. To confirm the results above, CD4 T cell cytokine production was analyzed by intracellular staining and flow cytometry. BLyS-DCs enhanced CD4 T cell IFN-γ production, but decreased or had no effect on IL-4 production by CD4 T cells when compared with Nil-DCs (Fig. 8C). Therefore, BLyS-DCs may be involved in Th1 immune responses by inducing CD4 T cells to produce high amounts of IFN-γ and low levels of IL-4, IL-5, and IL-10.
FIGURE 8. BLyS-DCs induce naïve CD4 T cell differentiation into Th1 T cells.

After DCs (0.5×105/well) were preactivated with or without 200 ng/ml BLyS, 20 ng/ml LPS, or 100 ng/ml CD40L for 24 h, cells were cocultured with freshly isolated allogeneic naïve CD4 T cells (2.5×105/well) for 2 days in round-bottomed 96 well plates. Culture supernatants were collected and Th1/Th2 cytokine production was measured using a human CBA (A and B). Data are representative of four independent experiments. C. DCs were pre-activated with or without 200 ng/ml BLyS, 100 ng/ml LPS, or 200 ng/ml CD40L for 24 h. After three washes, DCs (0.2×105/well) were cocultured with purified allogeneic naïve CD4 T cells (1×105/ well) for 5 days in flat-bottomed 96 well plates. On day 5, cells were restimulated with PMA plus ionomycin in the presence of brefeldin A for the last 4 h of culture, and cells were analyzed for intracellular IFN-γ and IL-4 staining using flow cytometry.
BLyS-induced DC maturation is not caused by LPS contamination
Because DCs are very sensitive to LPS stimulation, we needed to exclude the possibility that endotoxin contamination in the recombinant BLyS accounted for the ability of BLyS to stimulate DC maturation. Even though we excluded LPS contamination in the recombinant BLyS in our previous study by using polymyxin B and boiling (33), we subjected both BLyS and LPS, as a control, to heat inactivation by boiling for 30 minutes. We then assessed the ability of the heat-inactivated BLyS and LPS to induce surface marker expression as well as IL-12p70 cytokine expression as compared with control, non-heat-inactivated reagents. Heat-inactivation completely blocked BLyS-induced expression of surface markers, CD80, CD86, and CD83, whereas it had no effect on the ability of LPS to enhance surface marker expression (Fig. 9A). Similarly, pretreatment of BLyS with polymyxin B did not impair its ability to upregulate CD80 expression whereas it did, as expected, impair LPS-mediated induction of DC CD80 expression (Fig. 9B). Similar results were obtained when cytokine secretion was assayed instead. Thus, IL-12p70 was still induced by boiled LPS, but not by boiled BLyS (Fig. 9C). In summary, the direct effects of BLyS on DC maturation do not result from LPS contamination.
FIGURE 9. BLyS-induced maturation of DCs is not caused by endotoxin contamination.

Cells were cultured with control or heat-inactivated BLyS (ΔBLyS, 200 ng/ml) or LPS (ΔLPS, 20 ng/ml) for 24 h. A. Cells were analyzed for cell surface molecule expression by flow cytometry and the ΔMFI is indicated. Data are representative of four independent experiments. B. BLyS or LPS were pretreated with PB (20 µg/ml) for 20 min at room temperature, and then added into cells. Surface CD80 expression was measured. Data shown are from two independent experiments. C. Cell-free supernatants were analyzed for IL-12p70 by ELISA. * When compared with Nil, p=0.031.
BLyS-binding receptors
To determine how BLyS activates DCs, we first looked at BLyS binding using flow cytometry. BLyS binding was consistently detected; however, this occurred in a temperature dependent manner. BLyS binding was detectable when cells were incubated at 37°C (Fig. 10A), but not at 4°C (data not shown). Next, we tried to identify which receptor is responsible for BLyS binding on DCs. There are three known BLyS binding receptors- TACI, BCMA, and BAFF-R. None of the receptors were detectable on the cell surface using flow cytometry (Fig. 10B). Because we have previously shown that each antibody for the three receptors does detect their targets on normal human B cells (47), the results were not simply from antibody failure to detect their targets. Using normal TACI expressing B cells as a tool, we demonstrated that preincubation of cells with exogenous BLyS did not block the ability of the anti-TACI antibody to bind to receptor (data not shown), thereby excluding the possibility that our inability to detect TACI by flow cytometry resulted from epitope blocking. In addition, DCs lacked cytoplasmic BAFF-R by flow cytometry, and BCMA expression at the mRNA level was very heterogeneous among different donors (data not shown). In contrast, TACI expression was consistently detected by RT-PCR (data not shown), intracellular flow cytometry, and immunofluorescence (Fig. 10C). Even though surface TACI was not detectable by flow cytometry, some surface TACI was detected by confocal microscope in nonpermeabilized cells (Fig. 10D). Of interest, intracellular TACI was principally localized in the perinuclear area, which colocalized with the staining observed using the cis-Golgi matrix protein (GM130)-specific monoclonal antibody. Punctuate forms of TACI were also detected in the cytoplasm albeit to varying degrees among different donors (Fig. 10D and 10E). In addition, the same staining pattern of TACI was also detected in DCs freshly isolated from peripheral blood (data not shown). Upon incubation of cells at 37°C prior to staining, there was a marked increase in levels of punctuate TACI expression (data not shown). This observation is consistent with our flow cytometric assessment of BLyS binding, i.e., ligand binding was observed at 37°C. Finally, we also investigated receptor expression following stimulation of DCs with CD40L, BLyS, or LPS; however, none of these stimuli induced surface expression of any of the three BLyS binding receptors (data not shown).
FIGURE 10. BLyS receptor expression.
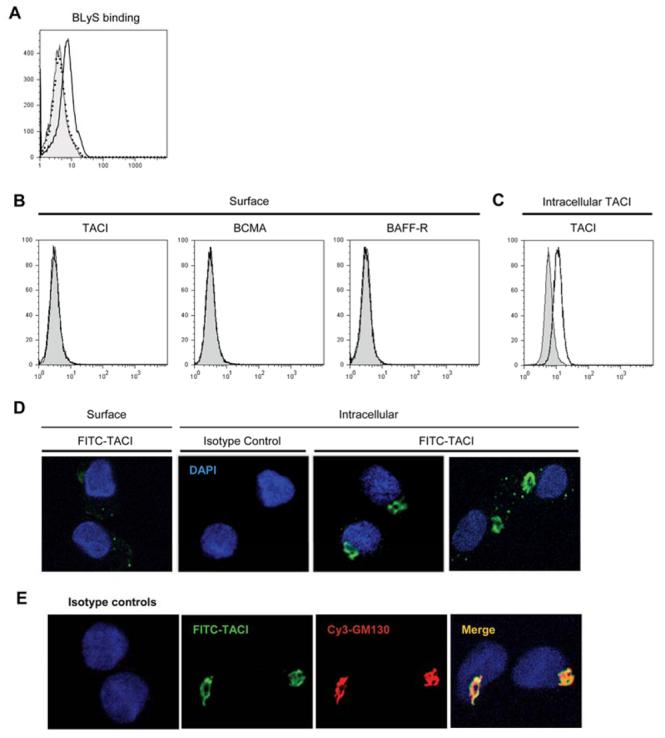
A. Soluble BLyS binding was determined by flow cytometry. DCs were incubated with (open histogram) or without (filled histogram) 2 μg/ml soluble BLyS for 2 h at 37°C, washed, incubated with biotinylated anti-BLyS antibody, and stained with streptavidin-conjugated PE. Isotype control was indicated by the dotted histogram. B. Surface staining of TACI, BCMA, and BAFF-R. C. Intracellular TACI staining. Specific receptor expression was indicated by the open histograms and isotype and fluorescence controls were indicated by the filled histograms. D. Subcellular localization of TACI in DCs by confocal immunofluorescence microscopy. Surface and intracellular TACI expression was assessed using a TACI specific goat antibody. E. Cells were costained with anti-TACI and anti-GM130 antibodies. Colocalization was confirmed when the images were merged (right panel). Isotype matched control primary antibody staining is also shown in D and E.
Discussion
It has been shown that BLyS levels are high in certain autoimmune diseases, such as Sjogren's syndrome, systemic lupus erythematosus, and rheumatoid arthritis (34-38). Here, we show that BLyS induces DC activation and maturation as revealed by enhanced expression of costimulatory molecules and cytokine production, but reduced phagocytic activity. Moreover, BLyS-DCs promoted naive CD4 T cell proliferation and differentiation into Th1 T cells. Our observations support the proposal that high BLyS levels in patients with autoimmune diseases may result in the activation of innate immune cells to secrete large amounts of proinflammatory cytokines, and subsequently modulate adaptive immune cells, which may be involved in the initiation or perpetuation of autoimmune diseases.
BLyS induced the expression of surface costimulatory molecules, CD40, CD80 and CD86, however, MHC class II expression was not upreguated by BLyS stimulation. Although activated DCs typically do acquire higher levels of HLA class II expression, there is precedence for stimuli that do not. Thus, Radhakrishnan et al. demonstrated that cross-linking the B7-DC molecule on DCs dramatically enhanced their ability to present MHC class II-peptide complexes in the absence of upregulation of MHC class II surface expression (49, 50). It is also interesting to note that there is evidence that signaling through BCMA renders B lymphocytes more efficient antigen presenting cells. Thus, Yang et al demonstrated APRIL or agonistic BCMA antibodies activated the JNK pathway which resulted in increased expression of CD40, CD80, CD86, MHC class II, and CD54 (51). Therefore, even though the expression of MHC class II was not upregulated on DCs by BLyS stimulation, BLyS signaling may augment Ag presentation in DCs by up-regulating costimulatory molecule expression through TACI.
In this study, we present clear evidence that BLyS induces DCs to secrete a variety of cytokines and chemokines. Although the absolute levels of cytokine/chemokine secretion were different, the overall pattern of cytokine induction by BLyS is strikingly similar to LPS stimulation. BLyS-DCs highly expressed GRO-α, MCP-2, MCP-4, MIP-1β, MIP-3α, MCP-1 and RANTES, suggesting that BLyS-DCs may recruit innate immune cells into the sites of inflammation, and thus, induce or amplify the inflammatory immune responses. Of interest, MCP-1 and RANTES were induced to a greater degree in BLyS-DCs than in LPS-DCs, supporting the conclusion that BLyS strongly promotes monocyte and T cell recruitment to aid in proinflammatory responses. BLyS-DCs expressed the T cell chemokine IP-10, but did not secrete detectable levels of Th2 T cell chemokines, TARC, MDC, and I-309 (Fig. 5B) (52-54). This observation is consistent with the ability of BLyS-DCs to secrete IL-12p70, but not IL-10 (Fig. 7A and 7B).
By inducing cytokine production and costimulatory molecule expression, BLyS-DCs promote naïve CD4 T cells to secrete high amounts of IFN-γ and IL-2, but not IL-4, IL-5, or IL-10. The enhanced IFN-γ production by CD4 T cells stimulated with BLyS-DCs was confirmed by intracellular staining. Previous studies showed that BLyS is involved in Th1 related autoimmune diseases, such as SLE, Sjogren's syndrome, and RA (35-38, 55-57). Another study showed that BLyS transgenic mice showed a much greater degree of Th1-mediated inflammatory immune responses (58). Our data also suggest that BLyS is involved in Th1 immune responses, which may be mediated through DC maturation by BLyS.
Recently discovered Th17 cells are distinguished from Th1 and Th2 cells via their ability to produce IL-17, a cytokine that has been shown to play a critical role in several inflammatory and autoimmune diseases (11, 59-63). The majority of studies to date have been carried out in the murine system, and it has been reported that IL-6 and TGF-β are required to differentiate naïve CD4 T cells into Th17 cells whereas IL-23 is required for the survival and effector function of Th17 cells (64-66). However, two very recent studies demonstrated that IL-23, IL-6, and IL-1β are required for human Th17 differentiation, and only one article studied whether APCs mediate Th17 differentiation (47, 48). The authors found that activated monocytes and directly purified CD1c (BDCA1) positive myeloid DCs, but not GM-CSF/IL-4 derived DCs, promoted Th17 cell differentiation (47). Since Th17 cells are involved in autoimmune diseases, we also examined whether BLyS-DCs are also involved in Th17 differentiation. We found that BLyS induced DCs to secrete high levels of IL-6 and moderate levels of IL-1β. In addition, we observed that BLyS stimulation resulted in very low levels of DC IL-23 secretion although it was quite variable between donors. Despite production of these three cytokines, BLyS matured DCs upon coculture with T cells did not induce the appearance of T cells expressing intracellular IL-17. Thus, it is possible that the levels of IL-6, IL-1β, and/or IL-23 induction by BLyS may not be sufficient for driving Th17 differentiation. Alternatively, high IFN-γ production by CD4 T cells activated by BLyS-DCs may inhibit Th17 differentiation as has recently been demonstrated in a study of human Th17 cells (68). However, it is possible that under different culture conditions, such as using directly purified DCs and/or employing different lengths of co-culture incubation time, a role for BLyS-DC in inducing IL-17 may be discovered. Studies of this nature are currently underway.
Another major focus of our investigation was to better understand the mechanism by which DCs respond to BLyS. When these studies were initiated, DCs were not known to express any of the three BLyS-binding receptors. Our studies very interestingly demonstrated that DCs constitutively express intracellular TACI, and it is principally localized in the intense perinuclear compartment, which may reflect the Golgi apparatus (Fig. 10E). This TACI localization pattern is strikingly distinct from that observed in B cells, which express the majority of this receptor in the plasma membrane (unpublished data). Of note, DCs isolated from peripheral blood also showed intracellular localization of TACI within the same compartment (data not shown). Although one previous study demonstrated that BCMA is localized in the Golgi apparatus in the U266 myeloma cell line (67), to our knowledge, this is the first suggestion that preformed TACI exists in the Golgi in DCs. It has been shown that TNF-R1 is expressed in the plasma membrane and complexes with the intracellular signaling molecule, TRADD, however, the primary subcellular location of TNF-R1 is the trans-Golgi network (68). Recent other reports also showed that TLR4 is localized in the Golgi and recognizes internalized LPS in intestinal epithelial cells (69). In addition, neoplastic non-Hodgkins lymphoma and Hodgkins lymphoma B cells have been shown to express autocrine BLyS and APRIL, and TACI and BCMA expression may permit enhanced cell proliferation and survival by binding to autocrine or paracrine sources of BLyS and APRIL (70). These authors also showed evidence of intracellular TACI staining co-localized with BLyS, suggesting that intracellular TACI may bind to endogenous BLyS or internalized BLyS inside of the cells and signal in this manner (70). Because DCs exhibit a high level of endocytosis, it is possible that intracellular TACI may bind to internalized BLyS thus activating DCs in an intracrine manner. However, it is also interesting to note that we were able to detect low level, punctuate TACI expression on the cell membrane using a sensitive confocal microscope method. This staining pattern was reproducibly accentuated upon incubation of cells at 37 °C (data not shown). The apparent temperature dependent increase in cell surface localized TACI is coincident with our observations that BLyS binding in DCs was only detectable by flow cytometry when we incubated cells with BLyS at 37°C, but not at 4°C (data not shown). Collectively, these two observations suggest the exciting possibility that during cell activation, cell surface TACI levels are rapidly increased by the transit from the Golgi to the cell surface, thereby resulting in an enhanced ability to bind BLyS. Such a process has been previously described for CD40L (71). In addition, previous studies have shown that the mature follicular dendritic cell (FDC) network is disrupted in BLyS -/-mice, but not in BAFF-R null mice, suggesting FDCs need BLyS for their maturation possibly through TACI or BCMA, but not BAFF-R (72, 73). Impaired production of lymphotoxin β by B cells could be one possible mechanism accounting for the defective FDC network in BLyS -/-mice. However, our study suggests an alternative explanation that FDC may mature directly in response to BLyS in a TACI-dependent manner.
Since DCs are known to produce BLyS, it raises a question of whether endogenous BLyS binds to TACI and activates DCs in an autocrine manner. However, in the steady state, we did not observe DC activation in the absence of stimulation, implying that baseline levels of endogenous BLyS expression are insufficient to activate DCs. Because we demonstrate that exogenous BLyS activates DCs, it is possible that exogenous BLyS significantly enhances endogenous BLyS expression. To test this possibility, we used RT-PCR to measure BLyS mRNA levels following stimulation of DCs with BLyS, LPS, and IFNγ as a positive control. In data not shown, although LPS and IFNγ increased BLyS mRNA levels as expected, treatment with exogenous BLyS did not increase BLyS mRNA levels. Therefore, we suggest that the underlying mechanism is not interaction of endogenous BLyS with TACI, but rather reflects activation by exogenous BLyS.
DCs clearly play a pivotal role in shaping Th-mediated immunity. Environmental signals are also key in determining whether DCs orchestrate a Th1, Th2, and/or Th17 immune response. Moreover, DCs also modulate regulatory T cells to promote their homeostasis and hence induction of immune tolerance (5). In our study, we expand the knowledge of signals regulating DC function by demonstrating that BLyS strongly activates DCs to secrete inflammatory cytokines, IL-6, IL-1β, and TNF-α, as well as instructs DC to induce naïve CD4 T cell proliferation and differentiation into Th1 CD4 T cells. Thus, our observations support the speculation that elevated levels of BLyS may break peripheral tolerance via its ability to activate DCs in a manner that also results in promotion of a Th1 response, leading ultimately to Th1-type immune disorders.
Acknowledgments
We thank Dr. A. B. Dietz for discussions about DC generation and Dr. S. Chi and Dr. M. McNiven for kindly providing us with the anti-GM130 antibody and helpful discussions.
Supported by the National Institutes of Health CA105258 and CA062242 (awarded to DFJ).
Footnotes
Competing interests statement: Not applicable.
References
- 1.Banchereau J, Briere F, Caux C, Davoust J, Lebecque S, Liu YJ, Pulendran B, Palucka K. Immunobiology of dendritic cells. Annu Rev Immunol. 2000;18:767–811. doi: 10.1146/annurev.immunol.18.1.767. [DOI] [PubMed] [Google Scholar]
- 2.Lanzavecchia A, Sallusto F. Regulation of T cell immunity by dendritic cells. Cell. 2001;106:263–266. doi: 10.1016/s0092-8674(01)00455-x. [DOI] [PubMed] [Google Scholar]
- 3.Steinman RM. The dendritic cell system and its role in immunogenicity. Annu Rev Immunol. 1991;9:271–296. doi: 10.1146/annurev.iy.09.040191.001415. [DOI] [PubMed] [Google Scholar]
- 4.Moser M, Murphy KM. Dendritic cell regulation of TH1-TH2 development. Nat Immunol. 2000;1:199–205. doi: 10.1038/79734. [DOI] [PubMed] [Google Scholar]
- 5.Chen M, Wang YH, Wang Y, Huang L, Sandoval H, Liu YJ, Wang J. Dendritic cell apoptosis in the maintenance of immune tolerance. Science. 2006;311:1160–1164. doi: 10.1126/science.1122545. [DOI] [PubMed] [Google Scholar]
- 6.Enk AH. Dendritic cells in tolerance induction. Immunol Lett. 2005;99:8–11. doi: 10.1016/j.imlet.2005.01.011. [DOI] [PubMed] [Google Scholar]
- 7.Hawiger D, Inaba K, Dorsett Y, Guo M, Mahnke K, Rivera M, Ravetch JV, Steinman RM, Nussenzweig MC. Dendritic cells induce peripheral T cell unresponsiveness under steady state conditions in vivo. J Exp Med. 2001;194:769–779. doi: 10.1084/jem.194.6.769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Steinman RM, Hawiger D, Liu K, Bonifaz L, Bonnyay D, Mahnke K, Iyoda T, Ravetch J, Dhodapkar M, Inaba K, Nussenzweig M. Dendritic cell function in vivo during the steady state: a role in peripheral tolerance. Ann N Y Acad Sci. 2003;987:15–25. doi: 10.1111/j.1749-6632.2003.tb06029.x. [DOI] [PubMed] [Google Scholar]
- 9.Tarbell KV, Yamazaki S, Steinman RM. The interactions of dendritic cells with antigen-specific, regulatory T cells that suppress autoimmunity. Semin Immunol. 2006;18:93–102. doi: 10.1016/j.smim.2006.01.009. [DOI] [PubMed] [Google Scholar]
- 10.Banchereau J, Pascual V, Palucka AK. Autoimmunity through cytokine-induced dendritic cell activation. Immunity. 2004;20:539–550. doi: 10.1016/s1074-7613(04)00108-6. [DOI] [PubMed] [Google Scholar]
- 11.Gutcher I, Becher B. APC-derived cytokines and T cell polarization in autoimmune inflammation. J Clin Invest. 2007;117:1119–1127. doi: 10.1172/JCI31720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Moore PA, Belvedere O, Orr A, Pieri K, LaFleur DW, Feng P, Soppet D, Charters M, Gentz R, Parmelee D, Li Y, Galperina O, Giri J, Roschke V, Nardelli B, Carrell J, Sosnovtseva S, Greenfield W, Ruben SM, Olsen HS, Fikes J, Hilbert DM. BLyS: member of the tumor necrosis factor family and B lymphocyte stimulator. Science. 1999;285:260–263. doi: 10.1126/science.285.5425.260. [DOI] [PubMed] [Google Scholar]
- 13.Schneider P, MacKay F, Steiner V, Hofmann K, Bodmer JL, Holler N, Ambrose C, Lawton P, Bixler S, Acha-Orbea H, Valmori D, Romero P, Werner-Favre C, Zubler RH, Browning JL, Tschopp J. BAFF, a novel ligand of the tumor necrosis factor family, stimulates B cell growth. J Exp Med. 1999;189:1747–1756. doi: 10.1084/jem.189.11.1747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Nardelli B, Belvedere O, Roschke V, Moore PA, Olsen HS, Migone TS, Sosnovtseva S, Carrell JA, Feng P, Giri JG, Hilbert DM. Synthesis and release of B-lymphocyte stimulator from myeloid cells. Blood. 2001;97:198–204. doi: 10.1182/blood.v97.1.198. [DOI] [PubMed] [Google Scholar]
- 15.Scapini P, Nardelli B, Nadali G, Calzetti F, Pizzolo G, Montecucco C, Cassatella MA. G-CSF-stimulated neutrophils are a prominent source of functional BLyS. J Exp Med. 2003;197:297–302. doi: 10.1084/jem.20021343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Novak AJ, Bram RJ, Kay NE, Jelinek DF. Aberrant expression of B-lymphocyte stimulator by B chronic lymphocytic leukemia cells: a mechanism for survival. Blood. 2002;100:2973–2979. doi: 10.1182/blood-2002-02-0558. [DOI] [PubMed] [Google Scholar]
- 17.Novak AJ, Grote DM, Stenson M, Ziesmer SC, Witzig TE, Habermann TM, Harder B, Ristow KM, Bram RJ, Jelinek DF, Gross JA, Ansell SM. Expression of BLyS and its receptors in B-cell non-Hodgkin lymphoma: correlation with disease activity and patient outcome. Blood. 2004;104:2247–2253. doi: 10.1182/blood-2004-02-0762. [DOI] [PubMed] [Google Scholar]
- 18.Krumbholz M, Theil D, Derfuss T, Rosenwald A, Schrader F, Monoranu CM, Kalled SL, Hess DM, Serafini B, Aloisi F, Wekerle H, Hohlfeld R, Meinl E. BAFF is produced by astrocytes and up-regulated in multiple sclerosis lesions and primary central nervous system lymphoma. J Exp Med. 2005;201:195–200. doi: 10.1084/jem.20041674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kato A, Truong-Tran AQ, Scott AL, Matsumoto K, Schleimer RP. Airway epithelial cells produce B cell-activating factor of TNF family by an IFN-beta-dependent mechanism. J Immunol. 2006;177:7164–7172. doi: 10.4049/jimmunol.177.10.7164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Gorelik L, Gilbride K, Dobles M, Kalled SL, Zandman D, Scott ML. Normal B cell homeostasis requires B cell activation factor production by radiation-resistant cells. J Exp Med. 2003;198:937–945. doi: 10.1084/jem.20030789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Craxton A, Magaletti D, Ryan EJ, Clark EA. Macrophage- and dendritic cell--dependent regulation of human B-cell proliferation requires the TNF family ligand BAFF. Blood. 2003;101:4464–4471. doi: 10.1182/blood-2002-10-3123. [DOI] [PubMed] [Google Scholar]
- 22.Wu Y, Bressette D, Carrell JA, Kaufman T, Feng P, Taylor K, Gan Y, Cho YH, Garcia AD, Gollatz E, Dimke D, LaFleur D, Migone TS, Nardelli B, Wei P, Ruben SM, Ullrich SJ, Olsen HS, Kanakaraj P, Moore PA, Baker KP. Tumor necrosis factor (TNF) receptor superfamily member TACI is a high affinity receptor for TNF family members APRIL and BLyS. J Biol Chem. 2000;275:35478–35485. doi: 10.1074/jbc.M005224200. [DOI] [PubMed] [Google Scholar]
- 23.Thompson JS, Schneider P, Kalled SL, Wang L, Lefevre EA, Cachero TG, MacKay F, Bixler SA, Zafari M, Liu ZY, Woodcock SA, Qian F, Batten M, Madry C, Richard Y, Benjamin CD, Browning JL, Tsapis A, Tschopp J, Ambrose C. BAFF binds to the tumor necrosis factor receptor-like molecule B cell maturation antigen and is important for maintaining the peripheral B cell population. J Exp Med. 2000;192:129–135. doi: 10.1084/jem.192.1.129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Thompson JS, Bixler SA, Qian F, Vora K, Scott ML, Cachero TG, Hession C, Schneider P, Sizing ID, Mullen C, Strauch K, Zafari M, Benjamin CD, Tschopp J, Browning JL, Ambrose C. BAFF-R, a newly identified TNF receptor that specifically interacts with BAFF. Science. 2001;293:2108–2111. doi: 10.1126/science.1061965. [DOI] [PubMed] [Google Scholar]
- 25.von Bulow GU, Bram RJ. NF-AT activation induced by a CAML-interacting member of the tumor necrosis factor receptor superfamily. Science. 1997;278:138–141. doi: 10.1126/science.278.5335.138. [DOI] [PubMed] [Google Scholar]
- 26.Yan M, Marsters SA, Grewal IS, Wang H, Ashkenazi A, Dixit VM. Identification of a receptor for BLyS demonstrates a crucial role in humoral immunity. Nat Immunol. 2000;1:37–41. doi: 10.1038/76889. [DOI] [PubMed] [Google Scholar]
- 27.Ng LG, Sutherland AP, Newton R, Qian F, Cachero TG, Scott ML, Thompson JS, Wheway J, Chtanova T, Groom J, Sutton IJ, Xin C, Tangye SG, Kalled SL, Mackay F, Mackay CR. B cell-activating factor belonging to the TNF family (BAFF)-R is the principal BAFF receptor facilitating BAFF costimulation of circulating T and B cells. J Immunol. 2004;173:807–817. doi: 10.4049/jimmunol.173.2.807. [DOI] [PubMed] [Google Scholar]
- 28.Ye Q, Wang L, Wells AD, Tao R, Han R, Davidson A, Scott ML, Hancock WW. BAFF binding to T cell-expressed BAFF-R costimulates T cell proliferation and alloresponses. Eur J Immunol. 2004;34:2750–2759. doi: 10.1002/eji.200425198. [DOI] [PubMed] [Google Scholar]
- 29.Huard B, Schneider P, Mauri D, Tschopp J, French LE. T cell costimulation by the TNF ligand BAFF. J Immunol. 2001;167:6225–6231. doi: 10.4049/jimmunol.167.11.6225. [DOI] [PubMed] [Google Scholar]
- 30.Schneider P. The role of APRIL and BAFF in lymphocyte activation. Curr Opin Immunol. 2005;17:282–289. doi: 10.1016/j.coi.2005.04.005. [DOI] [PubMed] [Google Scholar]
- 31.Ng LG, Mackay CR, Mackay F. The BAFF/APRIL system: life beyond B lymphocytes. Mol Immunol. 2005;42:763–772. doi: 10.1016/j.molimm.2004.06.041. [DOI] [PubMed] [Google Scholar]
- 32.Bossen C, Schneider P. BAFF, APRIL and their receptors: structure, function and signaling. Semin Immunol. 2006;18:263–275. doi: 10.1016/j.smim.2006.04.006. [DOI] [PubMed] [Google Scholar]
- 33.Chang SK, Arendt BK, Darce JR, Wu X, Jelinek DF. A role for BLyS in the activation of innate immune cells. Blood. 2006;108:2687–2694. doi: 10.1182/blood-2005-12-017319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Mackay F, Silveira PA, Brink R. B cells and the BAFF/APRIL axis: fast-forward on autoimmunity and signaling. Curr Opin Immunol. 2007;19:327–336. doi: 10.1016/j.coi.2007.04.008. [DOI] [PubMed] [Google Scholar]
- 35.Groom J, Kalled SL, Cutler AH, Olson C, Woodcock SA, Schneider P, Tschopp J, Cachero TG, Batten M, Wheway J, Mauri D, Cavill D, Gordon TP, Mackay CR, Mackay F. Association of BAFF/BLyS overexpression and altered B cell differentiation with Sjogren's syndrome. J Clin Invest. 2002;109:59–68. doi: 10.1172/JCI14121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Seyler TM, Park YW, Takemura S, Bram RJ, Kurtin PJ, Goronzy JJ, Weyand AC. BLyS and APRIL in rheumatoid arthritis. J Clin Invest. 2005 doi: 10.1172/JCI25265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Szodoray P, Jellestad S, Teague MO, Jonsson R. Attenuated apoptosis of B cell activating factor-expressing cells in primary Sjogren's syndrome. Lab Invest. 2003;83:357–365. [PubMed] [Google Scholar]
- 38.Zhang J, Roschke V, Baker KP, Wang Z, Alarcon GS, Fessler BJ, Bastian H, Kimberly RP, Zhou T. Cutting edge: a role for B lymphocyte stimulator in systemic lupus erythematosus. J Immunol. 2001;166:6–10. doi: 10.4049/jimmunol.166.1.6. [DOI] [PubMed] [Google Scholar]
- 39.Gross JA, Johnston J, Mudri S, Enselman R, Dillon SR, Madden K, Xu W, Parrish-Novak J, Foster D, Lofton-Day C, Moore M, Littau A, Grossman A, Haugen H, Foley K, Blumberg H, Harrison K, Kindsvogel W, Clegg CH. TACI and BCMA are receptors for a TNF homologue implicated in B-cell autoimmune disease. Nature. 2000;404:995–999. doi: 10.1038/35010115. [DOI] [PubMed] [Google Scholar]
- 40.Zhang M, Ko KH, Lam QL, Lo CK, Srivastava G, Zheng B, Lau YL, Lu L. Expression and function of TNF family member B cell-activating factor in the development of autoimmune arthritis. Int Immunol. 2005;17:1081–1092. doi: 10.1093/intimm/dxh287. [DOI] [PubMed] [Google Scholar]
- 41.Kalled SL. The role of BAFF in immune function and implications for autoimmunity. Immunol Rev. 2005;204:43–54. doi: 10.1111/j.0105-2896.2005.00219.x. [DOI] [PubMed] [Google Scholar]
- 42.Vogelsang P, Jonsson MV, Dalvin ST, Appel S. Role of dendritic cells in Sjogren's syndrome. Scand J Immunol. 2006;64:219–226. doi: 10.1111/j.1365-3083.2006.01811.x. [DOI] [PubMed] [Google Scholar]
- 43.Blanco P, Palucka AK, Gill M, Pascual V, Banchereau J. Induction of dendritic cell differentiation by IFN-alpha in systemic lupus erythematosus. Science. 2001;294:1540–1543. doi: 10.1126/science.1064890. [DOI] [PubMed] [Google Scholar]
- 44.Gottenberg JE, Cagnard N, Lucchesi C, Letourneur F, Mistou S, Lazure T, Jacques S, Ba N, Ittah M, Lepajolec C, Labetoulle M, Ardizzone M, Sibilia J, Fournier C, Chiocchia G, Mariette X. Activation of IFN pathways and plasmacytoid dendritic cell recruitment in target organs of primary Sjogren's syndrome. Proc Natl Acad Sci U S A. 2006;103:2770–2775. doi: 10.1073/pnas.0510837103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Santiago-Schwarz F, Anand P, Liu S, Carsons SE. Dendritic cells (DCs) in rheumatoid arthritis (RA): progenitor cells and soluble factors contained in RA synovial fluid yield a subset of myeloid DCs that preferentially activate Th1 inflammatory-type responses. J Immunol. 2001;167:1758–1768. doi: 10.4049/jimmunol.167.3.1758. [DOI] [PubMed] [Google Scholar]
- 46.Thomas R, Quinn C. Functional differentiation of dendritic cells in rheumatoid arthritis: role of CD86 in the synovium. J Immunol. 1996;156:3074–3086. [PubMed] [Google Scholar]
- 47.Acosta-Rodriguez EV, Napolitani G, Lanzavecchia A, Sallusto F. Interleukins 1beta and 6 but not transforming growth factor-beta are essential for the differentiation of interleukin 17-producing human T helper cells. Nat Immunol. 2007;8:942–949. doi: 10.1038/ni1496. [DOI] [PubMed] [Google Scholar]
- 48.Wilson NJ, Boniface K, Chan JR, McKenzie BS, Blumenschein WM, Mattson JD, Basham B, Smith K, Chen T, Morel F, Lecron JC, Kastelein RA, Cua DJ, McClanahan TK, Bowman EP, de Waal Malefyt R. Development, cytokine profile and function of human interleukin 17-producing helper T cells. Nat Immunol. 2007;8:950–957. doi: 10.1038/ni1497. [DOI] [PubMed] [Google Scholar]
- 49.Nguyen LT, Radhakrishnan S, Ciric B, Tamada K, Shin T, Pardoll DM, Chen L, Rodriguez M, Pease LR. Cross-linking the B7 family molecule B7-DC directly activates immune functions of dendritic cells. J Exp Med. 2002;196:1393–1398. doi: 10.1084/jem.20021466. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 50.Radhakrishnan S, Celis E, Pease LR. B7-DC cross-linking restores antigen uptake and augments antigen-presenting cell function by matured dendritic cells. Proc Natl Acad Sci U S A. 2005;102:11438–11443. doi: 10.1073/pnas.0501420102. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 51.Yang M, Hase H, Legarda-Addison D, Varughese L, Seed B, Ting AT. B cell maturation antigen, the receptor for a proliferation-inducing ligand and B cell-activating factor of the TNF family, induces antigen presentation in B cells. J Immunol. 2005;175:2814–2824. doi: 10.4049/jimmunol.175.5.2814. [DOI] [PubMed] [Google Scholar]
- 52.Bonecchi R, Bianchi G, Bordignon PP, D'Ambrosio D, Lang R, Borsatti A, Sozzani S, Allavena P, Gray PA, Mantovani A, Sinigaglia F. Differential expression of chemokine receptors and chemotactic responsiveness of type 1 T helper cells (Th1s) and Th2s. J Exp Med. 1998;187:129–134. doi: 10.1084/jem.187.1.129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Siveke JT, Hamann A. T helper 1 and T helper 2 cells respond differentially to chemokines. J Immunol. 1998;160:550–554. [PubMed] [Google Scholar]
- 54.Zingoni A, Soto H, Hedrick JA, Stoppacciaro A, Storlazzi CT, Sinigaglia F, D'Ambrosio D, O'Garra A, Robinson D, Rocchi M, Santoni A, Zlotnik A, Napolitano M. The chemokine receptor CCR8 is preferentially expressed in Th2 but not Th1 cells. J Immunol. 1998;161:547–551. [PubMed] [Google Scholar]
- 55.Mackay F, Sierro F, Grey ST, Gordon TP. The BAFF/APRIL system: an important player in systemic rheumatic diseases. Curr Dir Autoimmun. 2005;8:243–265. doi: 10.1159/000082106. [DOI] [PubMed] [Google Scholar]
- 56.Tan SM, Xu D, Roschke V, Perry JW, Arkfeld DG, Ehresmann GR, Migone TS, Hilbert DM, Stohl W. Local production of B lymphocyte stimulator protein and APRIL in arthritic joints of patients with inflammatory arthritis. Arthritis Rheum. 2003;48:982–992. doi: 10.1002/art.10860. [DOI] [PubMed] [Google Scholar]
- 57.Zheng Y, Gallucci S, Gaughan JP, Gross JA, Monestier M. A role for B cell-activating factor of the TNF family in chemically induced autoimmunity. J Immunol. 2005;175:6163–6168. doi: 10.4049/jimmunol.175.9.6163. [DOI] [PubMed] [Google Scholar]
- 58.Sutherland AP, Ng LG, Fletcher CA, Shum B, Newton RA, Grey ST, Rolph MS, Mackay F, Mackay CR. BAFF augments certain Th1-associated inflammatory responses. J Immunol. 2005;174:5537–5544. doi: 10.4049/jimmunol.174.9.5537. [DOI] [PubMed] [Google Scholar]
- 59.Stockinger B, Veldhoen M. Differentiation and function of Th17 T cells. Curr Opin Immunol. 2007;19:281–286. doi: 10.1016/j.coi.2007.04.005. [DOI] [PubMed] [Google Scholar]
- 60.Kolls JK, Linden A. Interleukin-17 family members and inflammation. Immunity. 2004;21:467–476. doi: 10.1016/j.immuni.2004.08.018. [DOI] [PubMed] [Google Scholar]
- 61.Harrington LE, Hatton RD, Mangan PR, Turner H, Murphy TL, Murphy KM, Weaver CT. Interleukin 17-producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages. Nat Immunol. 2005;6:1123–1132. doi: 10.1038/ni1254. [DOI] [PubMed] [Google Scholar]
- 62.Bettelli E, Carrier Y, Gao W, Korn T, Strom TB, Oukka M, Weiner HL, Kuchroo VK. Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature. 2006;441:235–238. doi: 10.1038/nature04753. [DOI] [PubMed] [Google Scholar]
- 63.Park H, Li Z, Yang XO, Chang SH, Nurieva R, Wang YH, Wang Y, Hood L, Zhu Z, Tian Q, Dong C. A distinct lineage of CD4 T cells regulates tissue inflammation by producing interleukin 17. Nat Immunol. 2005;6:1133–1141. doi: 10.1038/ni1261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Mangan PR, Harrington LE, O'Quinn DB, Helms WS, Bullard DC, Elson CO, Hatton RD, Wahl SM, Schoeb TR, Weaver CT. Transforming growth factor-beta induces development of the T(H)17 lineage. Nature. 2006;441:231–234. doi: 10.1038/nature04754. [DOI] [PubMed] [Google Scholar]
- 65.Langrish CL, Chen Y, Blumenschein WM, Mattson J, Basham B, Sedgwick JD, McClanahan T, Kastelein RA, Cua DJ. IL-23 drives a pathogenic T cell population that induces autoimmune inflammation. J Exp Med. 2005;201:233–240. doi: 10.1084/jem.20041257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Veldhoen M, Hocking RJ, Atkins CJ, Locksley RM, Stockinger B. TGFbeta in the context of an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells. Immunity. 2006;24:179–189. doi: 10.1016/j.immuni.2006.01.001. [DOI] [PubMed] [Google Scholar]
- 67.Gras MP, Laabi Y, Linares-Cruz G, Blondel MO, Rigaut JP, Brouet JC, Leca G, Haguenauer-Tsapis R, Tsapis A. BCMAp: an integral membrane protein in the Golgi apparatus of human mature B lymphocytes. Int Immunol. 1995;7:1093–1106. doi: 10.1093/intimm/7.7.1093. [DOI] [PubMed] [Google Scholar]
- 68.Jones SJ, Ledgerwood EC, Prins JB, Galbraith J, Johnson DR, Pober JS, Bradley JR. TNF recruits TRADD to the plasma membrane but not the trans-Golgi network, the principal subcellular location of TNF-R1. J Immunol. 1999;162:1042–1048. [PubMed] [Google Scholar]
- 69.Hornef MW, Frisan T, Vandewalle A, Normark S, Richter-Dahlfors A. Toll-like receptor 4 resides in the Golgi apparatus and colocalizes with internalized lipopolysaccharide in intestinal epithelial cells. J Exp Med. 2002;195:559–570. doi: 10.1084/jem.20011788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Chiu A, Xu W, He B, Dillon SR, Gross JA, Sievers E, Qiao X, Santini P, Hyjek E, Lee JW, Cesarman E, Chadburn A, Knowles DM, Cerutti A. Hodgkin lymphoma cells express TACI and BCMA receptors and generate survival and proliferation signals in response to BAFF and APRIL. Blood. 2007;109:729–739. doi: 10.1182/blood-2006-04-015958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Koguchi Y, Thauland TJ, Slifka MK, Parker DC. Preformed CD40 ligand exists in secretory lysosomes in effector and memory CD4+ T cells and is quickly expressed on the cell surface in an antigen specific manner. Blood. 2007 doi: 10.1182/blood-2007-03-081299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Rahman ZS, Rao SP, Kalled SL, Manser T. Normal induction but attenuated progression of germinal center responses in BAFF and BAFF-R signaling-deficient mice. J Exp Med. 2003;198:1157–1169. doi: 10.1084/jem.20030495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Vora KA, Wang LC, Rao SP, Liu ZY, Majeau GR, Cutler AH, Hochman PS, Scott ML, Kalled SL. Cutting edge: germinal centers formed in the absence of B cell-activating factor belonging to the TNF family exhibit impaired maturation and function. J Immunol. 2003;171:547–551. doi: 10.4049/jimmunol.171.2.547. [DOI] [PubMed] [Google Scholar]