

Abstract
The genetic architecture of plant defense against microbial pathogens may be influenced by pathogen lifestyle. While plant interactions with biotrophic pathogens are frequently controlled by the action of large-effect resistance genes that follow classic Mendelian inheritance, our study suggests that plant defense against the necrotrophic pathogen Botrytis cinerea is primarily quantitative and genetically complex. Few studies of quantitative resistance to necrotrophic pathogens have used large plant mapping populations to dissect the genetic structure of resistance. Using a large structured mapping population of Arabidopsis thaliana, we identified quantitative trait loci influencing plant response to B. cinerea, measured as expansion of necrotic lesions on leaves and accumulation of the antimicrobial compound camalexin. Testing multiple B. cinerea isolates, we identified 23 separate QTL in this population, ranging in isolate-specificity from being identified with a single isolate to controlling resistance against all isolates tested. We identified a set of QTL controlling accumulation of camalexin in response to pathogen infection that largely colocalized with lesion QTL. The identified resistance QTL appear to function in epistatic networks involving three or more loci. Detection of multilocus connections suggests that natural variation in specific signaling or response networks may control A. thaliana–B. cinerea interaction in this population.
PLANT resistance to pathogens involves multiple layers of defense. These range from nonhost resistance mechanisms against nonpathogens (or innate immune responses), defense responses effective against different classes or species of pathogens, to defense strategies effective against a limited range of pathogen isolates. Isolate-specific plant defense responses have been most extensively described with regard to the interaction between plants and biotrophic plant pathogens (Gabriel 1999; Dangl and Jones 2001; Jones and Dangl 2006). These isolate-specific interactions are primarily mediated by direct or indirect recognition of pathogen avr gene products (effectors) by plant proteins, activating a salicylic acid-dependent signaling cascade within the plant that leads to localized programmed cell death (Bent and Mackey 2007). While understanding these isolate-specific, qualitative interactions provides valuable contributions to both applied and general understanding of plant defense, interest has grown in discovering the bases of “partial” or quantitative resistance, with the goal of developing durable resistance to diverse pathogens (Niks and Rubiales 2002). Naturally variable resistance to necrotrophic plant pathogens such as Botrytis cinerea, Alternaria brassisicola, Plectosphaerella cucurmerina, or Sclerotinia sclerotiorum appears to be quantitative and polygenic (Kim and Diers 2000; Micic et al. 2004; Llorente et al. 2005; Finkers et al. 2007, 2008; Maxwell et al. 2007). However, the nature and extent of isolate-specific interaction between plants and necrotrophic pathogens is relatively unknown, and no qualitative naturally variable resistance genes effective against necrotrophic pathogens have been described (Jones and Dangl 2006).
The use of structured mapping populations to identify quantitative trait loci (QTL) provides a powerful approach to the study of quantitative traits, and a counterpoint to experiments focused on large-effect single loci (Young 1996). QTL mapping allows identification of genetic contributions to plant–pathogen interactions, without constraint by prior hypotheses. This approach exploits naturally occurring variation between parents of the mapping population and thus has potential to detect effects contingent on variation in, rather than absolute disruption of, gene expression or function. A whole-genome approach can also identify epistatic interaction between plant loci and test the dependency of identified plant loci on differences in either pathogen isolate or stage of infection.
Consistent with the proposition that plant resistance to necrotrophic pathogens has a complex genetic basis, QTL studies of plant resistance to necrotrophic pathogens have generally revealed the influence of numerous QTL of small phenotypic effect. Quantitative trait mapping of resistance to S. sclerotiorum in common bean, sunflower, and rapeseed revealed numerous small-effect QTL (Park et al. 2001; Bert et al. 2002; Zhao and Meng 2003). Studies of quantitative resistance to B. cinerea in tomato and Arabidopsis found QTL explaining 12–15% (tomato) and 5–10% (Arabidopsis) of phenotypic variance for traits associated with resistance to B. cinerea (Denby et al. 2004; Finkers et al. 2007). Despite the challenges manifest in uncovering the causal polymorphisms for small-effect QTL, genomics tools available for Arabidopsis thaliana offer promising opportunities to unravel network mechanisms underlying complex quantitative traits (Tonsor et al. 2005; Keurentjes et al. 2007; Wentzell et al. 2007; Ma and Bohnert 2008). To begin elucidating the molecular mechanisms underlying quantitative resistance, we analyzed quantitative A. thaliana responses to B. cinerea infection in a structured mapping population.
A. thaliana is one of hundreds of plant hosts susceptible to infection by the necrotrophic fungus B. cinerea, and possesses the most extensive resources for biological study in plants (Shindo et al. 2007). For the A. thaliana component of this study, we used the recombinant inbred line (RIL) population generated from the natural accessions Bayreuth-0 (Bay-0) and Shahdara (Loudet et al. 2002). This population is one of the largest A. thaliana mapping populations currently available, increasing our statistical power to dissect quantitative resistance shaping a complex plant/necrotroph interaction (Vales et al. 2005). Recently published partial genomic sequences from the Bay-0 and Shahdara parents will aid in identification of candidate polymorphisms between these accessions (Clark et al. 2007). This population has also been characterized for variation in nitrogen metabolism, nutrient content, trichome density, senescence, resistance to Pseudomonas syringae, variation in global transcript accumulation, and changes in transcript accumulation in response to salicylic acid (Loudet et al. 2003a,b; Symonds et al. 2005; Diaz et al. 2006; Kliebenstein et al. 2006; Perchepied et al. 2006; West et al. 2007). Another benefit of this population is that accessions from central Asia, such as Shahdara, form the most genetically distinct geographic grouping within the species A. thaliana, thus this population samples natural variation not present in the most commonly referenced A. thaliana RIL population, Col-0 × Ler (Sharbel et al. 2000; Nordborg et al. 2005). Finally, this population is an important tool for studying quantitative traits because the development of heterogenous inbred families (HIFs) from RILs possessing residual heterozygosity in identified genomic regions facilitates validation and controlled testing of QTL effects (Loudet et al. 2005).
For the pathogen component of this quantitative resistance analysis we used B. cinerea, a necrotrophic plant pathogenic fungus that infects a taxonomically diverse array of plants including most major dicot families, as well as pectinaceous monocots, nursery-grown conifers, and moss (Elad et al. 2004; De Leon et al. 2007; Williamson et al. 2007). This pathogen shows extensive phenotypic variation and genetic evidence supports the existence of high levels of recombination and minimal species subdivision based on host or geography (Calpas et al. 2001; Albertini et al. 2002; Rowe and Kliebenstein 2007). Studies of defined genetic lesions affecting known A. thaliana defenses provide a valuable framework for the biology of B. cinerea–plant interactions, which can contribute to identification of candidate genes (Prins et al. 2000; Glazebrook 2005; Robert-Seilaniantz et al. 2007; Williamson et al. 2007). Despite the documented genetic and phenotypic diversity of B. cinerea, many studies of molecular mechanisms of plant defense against B. cinerea are limited to single pathogen isolates. However, the existence of large collections of B. cinerea diversity allows us to query the effect of the pathogen's variation on host responses and underlying genetic architecture (Cui et al. 2004; Rowe and Kliebenstein 2007). These genetic and mechanistic tools in combination with the relative ease of quantifying plant tissue loss in response to B. cinerea infection make B. cinerea an optimal pathogen for studying quantitative resistance.
Plant production of antimicrobial secondary metabolites forms an important component of quantitative resistance variation. In particular, the indole-derived phytoalexin camalexin contributes to A. thaliana resistance to the necrotrophic fungal pathogens such as B. cinerea and A. brassicicola (Thomma et al. 1999; Ferrari et al. 2003). These necrotrophic pathogens possess intraspecific variation in the inhibitory effects of camalexin on both conidial germination in vitro and pathogen growth in planta (Kliebenstein et al. 2005; Sellam et al. 2007). While genes responsible for several steps in the camalexin biosynthetic pathway have been identified, the signaling components required for induction of camalexin biosynthesis are relatively uncharacterized, although salicylic acid, glutathione, and reactive oxygen species all contribute (Jirage et al. 1999; Zhou et al. 1999; Denby et al. 2005; Qutob et al. 2006; Nafisi et al. 2007; Parisy et al. 2007). Camalexin is easily and rapidly quantifiable, allowing simultaneous analysis of A. thaliana natural variation contributing to camalexin accumulation as well as pathogen-induced tissue death. Combining both pathogen virulence and plant response traits into the same analysis will provide insight into the mechanisms of camalexin induction by necrotrophic pathogens as well as how specific plant responses contribute to quantitative resistance.
To characterize the genetic architecture of A. thaliana resistance to the necrotrophic fungus B. cinerea and simultaneously test whether pathogen genotype influences this architecture, we identified QTL affecting lesion development and A. thaliana production of camalexin in response to infection by four divergent B. cinerea isolates in the A. thaliana Bay-0 × Shahdara RIL population. Numerous small-to-moderate effect QTL were identified, which affected Arabidopsis interactions with either all or a subset of single B. cinerea isolates. These results reflect the extreme phenotypic diversity in this population more than the actual QTL effect, as calculating allelic substitution effects for a subset of loci revealed changes in trait values ranging from 10 to 50%. Consistent with the described role of camalexin in defense against B. cinerea, 9 of 15 camalexin accumulation QTL colocalized with QTL influencing lesion size. Finally, epistatic interactions among QTL controlled a large fraction of the variation in both resistance and camalexin accumulation. We validated two of the most consistent QTL using HIFs and tested the distribution of effects at these loci using eight additional diverse B. cinerea isolates.
MATERIALS AND METHODS
Plant materials and growth conditions:
To identify polymorphic regions of the A. thaliana genome associated with susceptibility to B. cinerea, we used a population of 411 recombinant inbred lines (RILs) generated from the A. thaliana accessions Bay-0 and Shahdara (Loudet et al. 2002). The data we report are from two independent experiments, labeled “A” and “B”; each experiment consisted of two replicate plantings of the entire mapping population tested against three divergent B. cinerea isolates. Experiment A used B. cinerea isolates Fresa, Grape, and DN; Experiment B used B. cinerea isolates Fresa, Grape, and 83-2. To validate QTL and assess variation in QTL effects among B. cinerea isolates, we used HIFs generated from RILs 195 and 353. The HIF lines were generated from F7 RIL seed heterozygous at a single marker adjacent to identified B. cinerea QTL, creating matched pairs of lines containing a mixture of homozygous parental genomes representative of the original RIL and differing from each other only within the QTL region (Loudet et al. 2005).
Plants used for RIL experiment B and the QTL validation/HIF analysis were grown in 36-cell flats in large environmental chambers at 25°, 50–60% relative humidity, 120 μE with a photoperiod of 12 hr:12 hr light:dark to promote vegetative growth. Plants used in RIL experiment A were grown in 104-cell flats at similar photoperiod, temperature and humidity, but a light intensity of 150 μE. For both experiments, at least one plant per genotype of parental accessions Bay-0 and Shahdara was planted in a random location within each flat. Seeds were cold stratified in 1% phytagar for 3 days and sown directly in Premier Pro-Mix B soil (Quakertown, PA). Seedlings were thinned at 3 days postgermination to one plant per cell. Plants were fertilized biweekly with a 20:20:20 N:P:K mixture at a concentration of 1 g/liter. Plants were harvested ∼5 weeks postgermination.
Fungal isolates and infection conditions:
Collection, identification, and culture of B. cinerea isolates and the phylogenetic relationship between isolates Grape, DN, and 83-2 were as previously described (Rowe and Kliebenstein 2007). The isolates used for QTL mapping experiments were: Fresa (isolated in our laboratory from strawberries grown in San Diego County, CA), Grape (originally isolated from grapes in South Africa, provided by M. Vivier, University of Stellenbosch, Stellenbosch, South Africa), 83-2 (originally isolated from roses near San Francisco, CA, provided by D. Margosan, United States Department of Agriculture, Parlier, CA), and DN (isolated in our laboratory from a navel orange grown in Davis, CA). Additional B. cinerea isolates used for HIF tests, including published references and host plant at time of collection, are listed (Table 1).
TABLE 1.
Results of QTL validation using Bay × Sha HIFs 195 (Bcr.4.40; MSAT4.15) and 353 (Bcr.3.62; MSAT3.18)
| HIF195 (Bcr.4.5.40)
|
HIF353 (Bcr.3.5.62)
|
|||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Lesion-3
|
Camalexin
|
Lesion-3
|
Camalexin
|
|||||||
| Isolate | Source | Host | % diff | P | % diff | P | % diff | P | % diff | P |
| BMM | Zimmerli et al. (2001) | Geranium | −18 | ** | 11 | NS | −3 | NS | −14 | NS |
| BO5.10 | van Kan et al. (1997) | Grape | −3 | NS | 4 | NS | −14 | M | 6 | NS |
| BPA1 | Isolated 2005, D. J. Kliebenstein | Grape | −26 | M | 4 | NS | −9 | NS | −9 | NS |
| GLO2 | Rowe and Kliebenstein (2007) | Grape | 3 | NS | −9 | NS | 13 | NS | 14 | NS |
| KB2 | Rowe and Kliebenstein (2007) | Grape | −23 | * | −1 | NS | −9 | NS | −10 | NS |
| RASP | Rowe and Kliebenstein (2007) | Raspberry | −26 | *** | 1 | NS | 5 | NS | −13 | NS |
| PEPPER | Rowe and Kliebenstein (2007) | Pepper | −27 | * | 43 | * | −21 | M | −9 | NS |
| STEAK | Isolated 2006, H. C. Rowe | Tomato | −39 | * | −1 | NS | −6 | NS | −10 | NS |
| GRAPE | Rowe and Kliebenstein (2007) | Grape | −20 | * | −24 | * | −27 | M | −26 | * |
| FRESA | Isolated 2004, H. C. Rowe | Strawberry | −10 | NS | −13 | NS | −8 | NS | −22 | * |
| 83-2 | Rowe and Kliebenstein (2007) | Rose | −12 | NS | 0 | NS | 11 | NS | −29 | *** |
| DN | Rowe and Kliebenstein (2007) | Citrus | 0 | NS | −6 | NS | 6 | NS | −37 | * |
Available published references (Source) and collection host (Host) are given for each B. cinerea isolate tested. Lesion diameter at 72 hpi (Lesion-3) and nanogram camalexin accumulation per square centimeter leaf area (Camalexin) were measured for each HIF × isolate combination. % diff indicates the percent change in trait value produced by replacing the Shahdara allele with the Bay-0 allele at the tested locus [(Bay − Sha)/Sha]. P indicates the probability that phenotypic values obtained from lines carrying the Bay allele at the tested locus differ from lines carrying the Sha allele, as determined by factorial ANOVA incorporating three independent experiments per isolate: NS = P > 0.1, M = 0.1 > P > 0.05, *P < 0.05, **P < 0.01, ***P < 0.001.
Preparation of inocula and infection of A. thaliana were as previously described (Denby et al. 2004). Briefly, conidia were collected by flooding B. cinerea cultures grown on potato-dextrose agar with sterile distilled water. After filtering through glass wool to remove mycelial fragments, conidia were quantified with a hemocytometer and stored at −80° in 25% glycerol until use. Mature rosette leaves excised from 5-week old A. thaliana plants were placed in large plastic trays filled with 1% phytagar. Use of detached leaves allowed inoculation of leaves originating from a single plant with multiple B. cinerea isolates. This improves comparison of QTL among B. cinerea isolates by avoiding confounding interplant variability with genetic variation among isolates. Leaves were inoculated with 4-μl droplets of 105 conidia/ml in half-strength filtered organic grape juice (Santa Cruz Organics, CA). Plastic tray covers maintained humidity and leaves were incubated at room temperature. Leaves were monitored for lesion initiation at 8- to 12-hr intervals.
Phenotypic measurements:
B. cinerea pathogenicity:
Pathogenicity of B. cinerea isolates on A. thaliana Bay-0 × Shahdara RILs was estimated by measuring the diameter of the developing fungal lesion at 12- to 24-hr intervals postinoculation, depending on the rate of lesion development (Denby et al. 2004). Lesion time points were (1) 48 hours postinoculation (hpi), (2) 72 hpi, and (3) 88 hpi for experiment A and (i) 40 hpi, (1) 48 hpi, (2) 64 hpi, and (3) 72 hpi for experiment B. Lesion diameter in centimeters and total leaf area in square centimeters was obtained by hand analysis of high-resolution digital images (2048 × 1536 pixels) of infected leaves using ImageJ (Abramoff et al. 2004). Included scale objects allowed standardization of measurements across images.
Camalexin:
Accumulation of the A. thaliana phytoalexin, camalexin, within individual leaves was measured by high-performance liquid chromatography (HPLC) following termination of lesion monitoring, as previously described (Kliebenstein et al. 2005). Camalexin values were standardized by total leaf area (nanogram camalexin/square centimeters leaf area) and by lesion circumference. As both standardizations, as well as raw measurements (nanogram/leaf) produced similar QTL mapping results, only analyses of camalexin standardized by leaf area are presented. As camalexin is inducible by a number of stresses, we also measured camalexin content of uninfected leaves. Concurrent with the harvesting of leaves for B. cinerea inoculation, one leaf from each plant was harvested, photographed, and placed into 90% MeOH without treatment. These leaves were extracted and analyzed as above, except that the volume of extract analyzed was increased to 100 μl to improve quantification of the low concentrations of camalexin present in uninfected leaves.
Data analysis:
RILs:
Experiments A and B were analyzed independently due to differences in rates of lesion development. Broad-sense heritability of each trait by isolate combination was estimated independently within each experiment using the general linear model procedure within SAS. Broad sense heritability was defined as 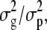 where
where 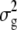 is the estimated genetic variance for the trait among different genotypes in this sample of RILs, and
is the estimated genetic variance for the trait among different genotypes in this sample of RILs, and 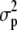 is the estimated total phenotypic variance for the trait. For this model, genotype was considered a fixed effect.
is the estimated total phenotypic variance for the trait. For this model, genotype was considered a fixed effect.
For each trait, least-squares means for each RIL × isolate combination were generated using PROC GLM (SAS Systems, Cary, NC). Genotypes for the Bay-0 × Shahdara RIL population are available at http://dbsgap.versailles.inra.fr/vnat/Documentation/33/DOC.html (Loudet et al. 2002). Composite interval mapping (CIM) and multiple interval mapping (MIM) analyses were performed using phenotypic data for each trait × isolate × experiment combination using Windows QTL Cartographer version 2.0 (Zeng et al. 1999). While the distribution of phenotypic values for camalexin showed positive skew, QTL mapping of log-transformed values did not produce significantly different results, so untransformed data is presented to simplify interpretation. Genomewide significance thresholds for CIM analyses were generated using 500 permutations of the data with α = 0.05 (Table 2) (Doerge and Churchill 1996). QTL were considered colocalized if the peak LOD score fell within the 2-LOD confidence interval calculated within the CIM module.
TABLE 2.
Trait means, estimated heritability, and QTL significance thresholds for lesion and camalexin phenotypes
| Trait | Isolate | Exp | Mean | SD | Bay | Sha | H2 | LOD |
|---|---|---|---|---|---|---|---|---|
| LESION1 | DN | A | 0.159 | 0.102 | 0.172 | 0.169 | 0.65 | 2.38 |
| LESION2 | DN | A | 0.535 | 0.216 | 0.581 | 0.514 | 0.61 | 2.25 |
| LESION3 | DN | A | 0.582 | 0.219 | 0.608 | 0.592 | 0.64 | 2.50 |
| CAMALEXIN | DN | A | 176.0 | 236.4 | 205.9 | 214.5 | 0.56 | 2.29 |
| LESION1 | FRESA | A | 0.170 | 0.110 | 0.193 | 0.154 | 0.60 | 2.43 |
| LESION2 | FRESA | A | 0.628 | 0.246 | 0.625 | 0.607 | 0.57 | 2.33 |
| LESION3 | FRESA | A | 0.691 | 0.379 | 0.699 | 0.647 | 0.54 | 2.04 |
| CAMALEXIN | FRESA | A | 131.1 | 115.5 | 142.2 | 114.4 | 0.64 | 2.37 |
| LESION1 | GRAPE | A | 0.122 | 0.085 | 0.107 | 0.130 | 0.57 | 2.20 |
| LESION2 | GRAPE | A | 0.459 | 0.200 | 0.416 | 0.388 | 0.57 | 2.32 |
| LESION3 | GRAPE | A | 0.515 | 0.233 | 0.454 | 0.412 | 0.57 | 2.30 |
| CAMALEXIN | GRAPE | A | 194.0 | 176.3 | 194.8 | 273.0 | 0.60 | 2.28 |
| LESION-i | 83-2 | B | 0.129 | 0.082 | 0.140 | 0.115 | 0.61 | 2.31 |
| LESION1 | 83-2 | B | 0.278 | 0.151 | 0.270 | 0.244 | 0.67 | 2.29 |
| LESION2 | 83-2 | B | 0.603 | 0.202 | 0.588 | 0.602 | 0.73 | 2.16 |
| LESION3 | 83-2 | B | 0.691 | 0.213 | 0.695 | 0.665 | 0.75 | 2.47 |
| CAMALEXIN | 83-2 | B | 212.4 | 153.2 | 194.9 | 199.7 | 0.72 | 2.33 |
| LESION-i | FRESA | B | 0.225 | 0.130 | 0.296 | 0.220 | 0.65 | 2.35 |
| LESION1 | FRESA | B | 0.453 | 0.155 | 0.469 | 0.445 | 0.60 | 2.48 |
| LESION2 | FRESA | B | 0.795 | 0.170 | 0.782 | 0.743 | 0.61 | 2.25 |
| LESION3 | FRESA | B | 0.942 | 0.177 | 0.941 | 0.877 | 0.63 | 2.26 |
| CAMALEXIN | FRESA | B | 114.5 | 99.3 | 92.5 | 92.0 | 0.74 | 2.29 |
| LESION-i | GRAPE | B | 0.171 | 0.123 | 0.160 | 0.147 | 0.58 | 2.37 |
| LESION1 | GRAPE | B | 0.358 | 0.164 | 0.366 | 0.311 | 0.58 | 2.24 |
| LESION2 | GRAPE | B | 0.739 | 0.174 | 0.744 | 0.733 | 0.58 | 2.33 |
| LESION3 | GRAPE | B | 0.861 | 0.181 | 0.855 | 0.821 | 0.56 | 2.45 |
| CAMALEXIN | GRAPE | B | 83.1 | 84.4 | 61.9 | 70.6 | 0.69 | 2.30 |
| CAMALEXIN | CONTROL | B | 24.7 | 84.8 | 28.8 | 0.8 | 0.67 | 2.26 |
Exp, differentiates experiment A from experiment B. Overall trait mean and standard deviation (SD) are provided for all trait-by-isolate combinations within each experiment. Bay and Sha indicate trait mean values for parental accessions interplanted with RILs. H2 is the broad-sense heritability estimate for each trait, estimated as the proportion of phenotypic variance explained by genotype in a one-way ANOVA model. LOD represents the significance threshold for QTL (P = 0.05) obtained by permutation tests.
The QTL distribution across the A. thaliana genome, including direction of allelic effects, was obtained from QTL Cartographer and visualized using EPCLUST (http://ep.ebi.ac.uk/EP/EPCLUST/), with traits clustered by location of significant QTL using the Pearson correlation distance between traits. MIM analyses employed forward and backward regression to generate an initial model. Models were subjected to at least three rounds of QTL position optimization, a new QTL search, and an epistasis search among detected QTL before QTL were tested for inclusion in the final model within the MIM algorithm. Five QTL identified consistently across isolates and experiments, as well as the epistatic interactions between these five loci, were specifically tested using ANOVA. QTL effects were estimated as the mean allelic substitution effect, or the difference attributable to the presence of Bay-0 vs. Shahdara alleles at markers adjacent to queried QTL.
HIFs:
HIFs were used to test specific genome regions associated with the Bcr.4.40 (HIF195) and Bcr.3.62 (HIF353) QTL (Loudet et al. 2005). These HIFs were tested with the four B. cinerea isolates originally used for QTL mapping, plus eight additional isolates (Table 1). Experiments employed 12 detached leaves for each genotype × isolate combination, with individual plants providing 4–6 leaves randomly assigned to different B. cinerea isolates. Each isolate was tested in three independent experiments; five experiments were required to test each isolate three times, providing 36 independent measurements per genotype × isolate combination. Phenotypic measurements for parental accessions and HIFs were obtained as described for RILs. For each HIF pair, we performed an ANOVA using a general linear model. In this model yig denotes the virulence of the plant pathogen interaction on A. thaliana genotype g, measured using the Botrytis isolate i on leaf l. The ANOVA model for the virulence, measured by lesion size or camalexin induction is: yigl = μ + Gg + Ii + (GI)gi + ɛigl, where g is Bay-0 or Sha for the HIF and the levels of i and l differ depending upon the experiment. The main effects are denoted as G and I and represent A. thaliana accession and Botrytis isolate, respectively. ɛigl represents the error and is assumed to be normally distributed with mean 0 and variance 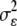 . Nested effects of flat or plant were included as random effects where appropriate. Within this model t-tests adjusted with Tukey's HSD were used to test for significant differences attributable to plant genotype for each B. cinerea isolate.
. Nested effects of flat or plant were included as random effects where appropriate. Within this model t-tests adjusted with Tukey's HSD were used to test for significant differences attributable to plant genotype for each B. cinerea isolate.
RESULTS
Trait heritability and distributions:
Expansion of necrotic lesions and the accumulation of camalexin are both phenotypic measures of B. cinerea interaction with A. thaliana leaves. To investigate the genetic architecture of B. cinerea interactions with A. thaliana, we measured both phenotypes on 411 A. thaliana Bay-0 × Sha RILs infected with four different B. cinerea isolates. In all experiments Botrytis-infected A. thaliana leaves developed visible necrotic lesions at 40–48 hpi. Due to differences in lesion progression between independent experiments conducted on two separate RIL plantings (experiments A and B), lesion development was measured at different time points and the two experiments analyzed separately. To facilitate comparison between the experiments, similar lesion phenotypes are labeled as follows: lesion-i = 40 hpi (B only), lesion-1 = 48 hpi (both experiments), lesion-2 = 64 hpi (A) or 72 hpi (B), lesion-3 = 72 hpi (A) or 88 hpi (B). Camalexin accumulation was measured immediately following observation of the lesion-3 time point.
Estimates of the broad-sense heritability of B. cinerea lesion size were similar for early and late lesion time points (Table 2). Heritability estimates for camalexin accumulation were slightly higher than those for lesion size. The divergent B. cinerea isolates produced similar RIL phenotype distributions; lesion size showed a slight negative skew and camalexin accumulation a stronger positive skew (supplemental Figures 1 and 2). A shift in lesion size indicates greater aggressiveness of Grape and Fresa in experiment B than in experiment A; this increased lesion size was associated with identification of more QTL in experiment B. Overall, lesion traits and camalexin accumulation showed slightly higher heritability in experiment B (59.3%) than experiment A (64.6%). While lesion traits were more correlated within experiments than between experiments, camalexin accumulation per genotype showed significant positive correlation across all isolate by experiment combinations (Pearson coefficient = 0.21–0.42, P < 0.0001; supplemental Figure 3). Plants infected with Fresa accumulated higher mean levels of camalexin than those infected with Grape across both experiments, while the highest camalexin accumulation was observed in RILs infected with 83-2 (supplemental Figure 2). While mean differences between the Bay-0 and Shahdara accessions were minimal, we observed both positive and negative transgressive segregation for late lesion (72–88 hpi) diameter and camalexin accumulation (Table 2, supplemental Figures 1 and 2). This implies that the similar B. cinerea resistance phenotypes shown by the Bay-0 and Shahdara parental lines result from the presence of distinct genetic polymorphisms with antagonistic effects contributed by each parent.
Identification of QTL:
Our analyses identified 23 QTL with statistically significant influence on B. cinerea lesion size or camalexin accumulation in the A. thaliana Bay-0 × Shahdara RIL population (Figure 1, supplemental Table 1). QTL are named as Botrytis cinerea resistance loci with chromosome and approximate centimorgan location [Bcr.chromosome.cM]. Despite differences in trait distributions and heritability, QTL positions were largely in agreement between experiments A and B (Figure 1; supplemental Figures 1 and 2). Six QTL were specific to single B. cinerea isolates, while 4 QTL significantly influenced plant response to all B. cinerea isolates tested (Figures 1 and 2B). Of the 23, 9 QTL influenced both lesion size and camalexin accumulation, while 8 and 6 loci controlled only lesion size or camalexin, respectively (Figure 2A). The 4 QTL affecting all B. cinerea isolates were detected for both lesion and camalexin traits, however Bcr.3.62 affected camalexin accumulation for all isolates but lesion size for only two isolates, specifically at late lesion time points. Camalexin accumulation and lesion-size traits were negatively correlated, with the strongest correlation detected between camalexin accumulation and late lesion time points (supplemental Figure 3).
Figure 1.—

Genomic locations of QTL identified for B. cinerea lesion size and A. thaliana camalexin accumulation. A. thaliana chromosomes are identified by roman numerals (I–V), with centimorgans ascending from the left; chromosomes are separated by gray lines. Each row represents a B. cinerea isolate × trait × experiment combination, with isolate and experiment indicated by text labels, and trait indicated by text color (green, lesion-1, lesion-i; orange, lesion-2; red, lesion-3; black, camalexin). CTRL indicates noninoculated leaves. Rows are clustered to show overall similarity of QTL architecture, using the weighted group average (WPGMA) linkage based on the absolute value of Pearson correlation distances. Lesion phenotypes (top) and camalexin (bottom) are clustered separately. Colored cells indicate QTL significant at P = 0.05 in CIM models; blue indicates positive association of trait values with Bay alleles and yellow indicates positive association of trait values with Sha alleles. QTL selected for directed analysis of epistasis are shown with gray arrows at the lower edge of the heat map. QTL positions, LOD scores, additive effects, and R2 values are provided in supplemental Table 1.
Figure 2.—

Distribution of significant QTL among phenotypic traits and B. cinerea isolates. Circles or ovals represent phenotypic traits or B. cinerea isolates, respectively. Number of statistically significant QTL detected as influencing a given trait (for any isolate) or isolate (for any trait) are shown, with values located within overlapping shapes indicating shared QTL. (A) Phenotypic traits. Early lesion includes traits lesion-i (experiment B only) and lesion-1; late lesion includes lesion-2 and lesion-3. (B) B. cinerea isolates.
Detection of three QTL (Bcr.1.06-10, Bcr.2.52, and Bcr.3.00) for multiple isolates but only within single experiments suggests the influence of uncontrolled environmental factors at these loci. To explore whether QTL detection as isolate- or trait-specific occurred as an artifact caused by limited experimental power, we examined the distribution of QTL detected at LOD = 1, well below the 0.05 significance thresholds calculated empirically via permutations (LOD = 2.04–2.50; Table 2). Including QTL that were detected but not statistically significant increased the number of QTL shared among all traits and all isolates, but trait- and isolate-specific QTL were still detected (supplemental Figure 4). This suggests that some isolate-specific interactions can be attributed to variation in QTL effect that is expected in a trait controlled by numerous loci. This is unlikely to be caused by genotype × environment interactions as each isolate was tested against the same plants. Finally, genetic variation affecting B. cinerea lesion development and camalexin accumulation are closely associated in this population.
QTL effects:
As suggested by transgressive trait distributions within the RILs, both A. thaliana parental genotypes contributed to increased susceptibility to B. cinerea (Figure 3; supplemental Figures 1 and 2). Shahdara alleles at Bcr.1.80 and Bcr.4.40 were associated with larger B. cinerea lesions and, in some instances, with lower camalexin accumulation, while Bay-0 alleles at Bcr.2.61, Bcr.3.00, and Bcr.5.20 conferred larger lesion sizes (Figure 1). All lesion-size QTL that colocalized with camalexin QTL showed opposite allelic effects, consistent with the described role of camalexin in A. thaliana resistance to B. cinerea (Kliebenstein et al. 2005; Ferrari et al. 2007) (Figures 1 and 3). Using Pearson correlation distances to evaluate overall similarity in QTL patterns identified among different B. cinerea isolates, experiments, and time points, detected partial clustering of QTL affecting lesion size by time point. This was primarily driven by the detection of QTL Bcr.3.00 at early time points and detection of Bcr.4.40 primarily at later time points (Figure 1). This suggests that different A. thaliana genes are involved in early and late responses to B. cinerea infection. A trend toward clustering QTL by experiment reflects the identification of experiment-specific QTL, possibly due to environmental differences between experiments or the use of different B. cinerea isolates (DN vs. 83-2).
Figure 3.—

Estimated phenotypic effect of substituting the Bay-0 allele for the Shahdara allele at markers associated with five QTL clusters. The vertical axis shows the percent difference in trait mean between all RILs carrying the Bay-0 allele vs. all RILs carrying the Shahdara allele at the queried locus. Bar shading differentiates B. cinerea isolates, as indicated in the figure legend. Two values each are presented for isolates GRAPE and FRESA, representing experiments A and B. Significance of QTL as determined by ANOVA (Tukey's HSD) is indicated by asterisks above bars (*P < 0.05; **P < 0.01; ***P < 0.001). Trait means, standard error, and specific P-values for Bay-0 and Shahdara alleles at these QTL regions are presented in supplemental Table 2. Traits: (A) early (48 hpi) B. cinerea lesion (centimeter lesion diameter); (B) late (72–88 hpi) B. cinerea lesion (centimeter lesion diameter); and (C) camalexin accumulation (nanograms/square centimeter leaf area).
Genotype at a given QTL explained only a small proportion of genetic variance within QTL models (2–12%; supplemental Table 1). However, given the wide distribution of phenotypes in the RILs, allelic substitution values may provide a more biologically meaningful estimate of QTL effect. We calculated allelic substitution values, or the percent change in phenotype associated with substitution of the Bay-0 allele for the Shahdara allele at a given locus, for five QTL clusters identified across multiple experiments and B. cinerea isolates, Bcr.1.80, Bcr.2.61, Bcr.3.62, Bcr.4.40, and Bcr.5.20-27 (Figure 3). Bcr.5.20-27 represents two loci, Bcr.5.20 and Bcr.5.27, detected as separate QTL but sharing association with the marker NGA139. Given their genetic proximity and this population size we could not accurately partition them as two QTL and thereby treated this as one locus. Allelic substitution values for late lesion time points showed significant effects from 4 to 13%, but stronger phenotypic effects for early lesion time points (12–28%) and camalexin accumulation (5–49%) (Figure 3, supplemental Table 2). The observation that allelic effects are larger than the estimated R2 is consistent with the broad distribution of phenotypic values (supplemental Figures 1 and 2), and suggests that additional, undetected loci (or interactions among loci) influence A. thaliana–B. cinerea interaction in this population.
Identification of epistasis:
Recent studies have shown that allele-specific epistatic interactions involving two or more loci control plant/pathogen resistance gene networks or central metabolism (Bomblies et al. 2007; Rowe et al. 2008). These epistatic interactions contribute to phenotypic variability but hinder detection and effect estimation for QTL examined singly. A survey of epistasis with the MIM module of QTL Cartographer detected few instances of epistasis in our experiments, but this module only tests pairwise interactions involving two loci. MIM models generally explained a substantially lower proportion of observed trait variance (11–40%) than could be attributed to plant genotype on the basis of heritability estimates (Table 2). This suggested a failure of MIM models to identify and include higher-order interactions among loci. To assess the contribution of higher-order epistases via a targeted search, the five main QTL clusters identified across multiple experiments and B. cinerea isolates, Bcr.1.80, Bcr.2.61, Bcr.3.62, Bcr.4.40, and Bcr.5.20-27, were specifically tested for epistatic interaction via factorial ANOVA (Figures 1 and 4). These analyses tested all pairwise and three-way combinations of the markers closest to each target QTL, but could not test interactions of higher orders due to limited allele combinations present in 411 RILs. This identified significant pairwise and three-way epistases for lesion traits and camalexin accumulation with epistatic interactions more frequently detected for camalexin accumulation than lesion-size traits (Figure 4). The higher heritability of camalexin accumulation in comparison to lesion development likely provides greater statistical power to detect epistases.
Figure 4.—

Epistatic interactions among five main-effect QTL. Tested QTL are shown as ovals. Arrows indicate significant pairwise interactions; triangles show significant three-way interactions. Significance of interactions was determined by ANOVA at α = 0.05. Line types denote isolates: solid, GRAPE; dashed, FRESA; dotted (round), DN (A and C only); dotted (square), 83-2 (B and D only). (A) Late lesion (experiment A); (B) Late lesion (experiment B); (C) Camalexin (experiment A); (D) Camalexin (experiment B).
Limited analysis of interaction among these five QTL revealed several instances where epistatic interactions among QTL may obscure relationships between loci and phenotypes. In experiment A, analysis of resistance to B. cinerea Fresa detected Bcr.1.80 as significantly affecting early and late lesion size and a significant interaction between Bcr.1.80 and Bcr.2.61 altered late lesion development (supplemental Table 1, Figure 1). However, Bcr.1.80 was not identified via CIM, MIM, or ANOVA as individually influencing camalexin accumulation in response to Fresa in this experiment (Figure 3, supplemental Table 1). Instead, Bcr.1.80 significantly alters camalexin accumulation in response to Fresa via epistatic interactions with both Bcr.2.61 and Bcr.5.20 (Figure 4). Similarly, Bcr.2.61, which was not detected as influencing A. thaliana interaction with B. cinerea 83-2 in the original single-locus analysis, participates in a three-way epistatic interaction with Bcr.1.80 and Bcr.3.62 that influences lesion size (Figures 3 and 4). Accounting for these complex interactions is not only essential for developing strategies to clone B. cinerea resistance QTL, but may also allow the useful inclusion of coexpression and protein interaction data in formulation of hypotheses regarding mechanisms of A. thaliana resistance to B. cinerea.
Priming of defense:
To test whether QTL detected for camalexin accumulation related to differences in response to B. cinerea infection, rather than differences in basal camalexin levels, we measured camalexin accumulation in untreated leaves and mapped QTL. Colocalization of basal camalexin QTL with QTL controlling camalexin accumulation in response to B. cinerea infection would suggest that the causal polymorphisms for these QTL alter “priming” or constitutive stress responses. The proportion of RILs accumulating detectable levels of camalexin in leaves not inoculated with B. cinerea (∼1/3) was similar between experiments A and B. However the level of camalexin accumulation in uninfected leaves was extremely low in experiment A, yielding insufficient power to detect QTL.
Noninoculated experiment B plants accumulated an average of 25 ng camalexin/cm2 leaf area, levels dramatically lower than those found in B. cinerea-infected leaves (Table 2). Genotypic means for camalexin accumulation in noninoculated plants showed positive correlation with values obtained for B. cinerea-treated plants from the same experiment (Pearson coefficients = 0.12–0.19, P = 0.02–0.003), but camalexin values among B. cinerea-treated leaves were more strongly correlated, even across experiments (Pearson coefficients = 0.47–0.66 within experiment B, Pearson coefficients = 0.21–0.42 across experiments) (supplemental Figure 3). This suggests that genetic control of basal camalexin accumulation is distinct from control of B. cinerea-induced camalexin accumulation, but some elements may be shared.
CIM results suggested minimal overlap between genetic control of differences in camalexin accumulation between Bay-0 and Shahdara with and without B. cinerea infection (Figure 1). MIM analysis of basal camalexin levels produced a QTL model explaining 91% of genetic variance, predominantly through epistatic interactions. This model contains four QTL, each interacting epistatically with two to three other QTL in the model (supplemental Figure 5). While one of these loci, Bcr1.80, was identified as influencing camalexin and/or lesion development in B. cinerea-infected leaves, two QTL on chromosome 2, controlling 16% of phenotypic variation additively and 57% of phenotypic variance through epistatic interactions, were not detected for any B. cinerea phenotypes (Figure 1, supplemental Table 1). The fourth interacting QTL, Bcr4.36, detected as contributing a significant main effect in the CIM analysis, colocalizes with a QTL affecting lesion size for Fresa and may not be distinct from the QTL cluster Bcr4.40. The influence of Bcr.2.13 and Bcr.2.39 on camalexin accumulation in infected plants may be hidden by responses induced by pathogen infection, or these loci may represent polymorphism in a response pathway that is not activated or inhibited by B. cinerea infection. In contrast, Bcr1.80 and Bcr4.40 may represent polymorphisms in core defense signaling mechanisms that, when combined, lead to inappropriate stimulation of defense responses such as camalexin accumulation.
QTL validation and B. cinerea isolate dependency:
HIFs generated from selected Bay × Sha RILs were used to confirm two consistently identified QTL, Bcr.4.40 (HIF195) and Bcr.3.62 (HIF353). These HIFs were also used to test the influence of these QTL on A. thaliana interactions with a broader sample of B. cinerea isolates. Both lesion size and camalexin accumulation were tested for these two HIFs, despite Bcr.4.40 being primarily detected as influential for lesion size and Bcr.3.62 primarily influencing camalexin accumulation.
HIF195 was the only HIF available to test Bcr.4.40. The Bay allele at this locus is associated with decreased lesion size at late time points for all 4 isolates used for QTL mapping (Figures 1 and 3B). While 10 of 12 B. cinerea isolates tested (including the original 4 isolates) developed smaller lesions on plants carrying the Bay allele at Bcr.4.40 (MSAT4.15), this difference was only statistically significant for 6 isolates (Table 1). Interestingly, these 6 isolates included only one of the original QTL isolates, Grape. As a limited analysis of epistasis identified complex interactions between Bcr.4.40 and several other loci, we hypothesized that HIF195 does not possess the optimal genetic background to observe the phenotypic impact of Bcr.4.40 (Figure 4). Examining the experimental data for the RIL genotypic classes delineated by the five QTL used for epistasis analysis suggests that this is the case, as RILs sharing background genotypes with HIF195 show no statistically significant difference in late lesion size between lines possessing the Bay-0 or Shahdara allele at MSAT4.15. The B. cinerea isolate DN provides the strongest example of this, as Bcr.4.40 was detected as a significant lesion size QTL but RILs containing the two HIF195 genotypes showed no phenotypic difference for this isolate (Table 1; supplemental Figure 6A). Alternatively, the QTL detected for DN near MSAT4.15 might represent a different influential polymorphism than the QTL detected for the other B. cinerea isolates, which is located outside the recombination breakpoints of HIF195. Nevertheless, HIF195 confirms the presence of at least one locus for B. cinerea resistance in this region. Isolates showed no consistent pattern in camalexin accumulation between the two HIF195 genotypes, with two isolates, Grape and Pepper, identifying opposing allelic effects at this locus. Together, the HIF195 data support the presence of genetic loci within A. thaliana that influence plant interaction with only a subset of Botrytis isolates, which may also separate control of the correlated traits lesion size and camalexin accumulation.
HIF353 was used to test the Bcr.3.62 region (MSAT3.18), for which the Bay allele was associated with lower camalexin accumulation across isolates. Again, 10 of 12 tested B. cinerea isolates showed a trend of lower camalexin accumulation associated with the Bay allele at MSAT3.18, but this difference was only statistically significant for 3 isolates, all isolates used in the original QTL mapping experiments (Table 1). The DN isolate identified the largest effect of Bcr.3.62 on camalexin accumulation. While participation of the Bcr.3.62 locus in multilocus epistatic interactions may hinder observation of the full phenotypic impact of this locus on camalexin accumulation, examination of potential interactions of Bcr.3.62 with the four other QTL used for epistasis analysis suggests that HIF353 provides a favorable, if not optimal, genetic background for detecting the effects of this locus on B. cinerea-mediated camalexin accumulation (supplemental Figure 6B). This does not preclude the possibility that such effects may apply to other B. cinerea isolates, or involve loci not included in our minimal analysis of epistasis. The HIF353 results confirm that this region contains at least one locus for camalexin accumulation in response to B. cinerea infection. No clear trend could be distinguished regarding the effect of this locus on lesion size; both positive and negative allelic effects were detected among isolates, with no statistically significant differences.
DISCUSSION
This study supports the hypothesis that genetic control of A. thaliana response to B. cinerea infection is complex. Consistent with prior observations, we detected numerous QTL with moderate phenotypic effects as influencing A. thaliana interactions with Botrytis cinerea. Contributions to B. cinerea resistance from both A. thaliana parental genotypes produced transgressive segregation for both lesion and camalexin traits. In contrast to previous analysis, we identified QTL that may control resistance to numerous B. cinerea isolates and found significant evidence for pairwise and three-way epistatic interactions (Figure 4) (Denby et al. 2004). Differences in QTL detection among B. cinerea isolates, as well as among infection time points, add new dimensions to this complexity. Recognition and assessment of sources of variation in biotic interactions is essential for developing a realistic understanding of how plants interact with Botrytis cinerea and other necrotrophic pathogens, with the ultimate goal of obtaining durable resistance in crop plants.
Camalexin QTL:
Colocalization of roughly half of significant QTL affecting lesion size with camalexin accumulation QTL of opposing allelic effects highlights the positive relationship between camalexin accumulation and A. thaliana resistance to B. cinerea. None of the identified camalexin QTL colocalized with previously described camalexin biosynthetic genes (Zhou et al. 1999; Mikkelsen et al. 2003; Nafisi et al. 2007), suggesting that these QTL may influence camalexin signaling. Previously described elements of camalexin signaling have been identified through study of mutagenized or genetically manipulated A. thaliana, thus analysis of natural variation for camalexin signaling may provide new insight into the evolution and regulation of this defense response (Glazebrook and Ausubel 1994; Denby et al. 2005; Ren et al. 2008).
In addition to variation in plant regulation of camalexin biosynthesis or accumulation, our study indicates that pathogen variation alters camalexin accumulation. The B. cinerea isolate 83-2 induced higher levels of camalexin than the other isolates tested, but QTL analysis with 83-2 did not identify any camalexin accumulation QTL that were strictly unique to this isolate. Of significant QTL, only Bcr1.19 appeared unique to camalexin induced by the B. cinerea isolate 83-2, however a nearby, potentially overlapping QTL also affects early lesion formation by Grape (Figure 1, supplemental Table 1). This suggests that genetic variation in the pathogen, rather than the plant, controls this difference in camalexin induction among B. cinerea isolates. B. cinerea isolate variation, as identified in the 83-2 isolate, can provide a useful tool for refining the necrotrophic pathogen signals that lead to the regulation of specific defense responses by the plant. Additionally, B. cinerea isolate variation improves our analysis of QTL position for both camalexin and lesion traits—higher levels of camalexin accumulation provided greater measurement accuracy and statistical power for this trait, and linkage of camalexin accumulation to lesion size may allow this biochemical trait to serve as a surrogate trait for lesion size in early stages of fine mapping.
Lesion development:
Detection of QTL specific to early vs. late lesion time points suggests that genetic contributions to plant defense may change as infection develops (Figures 1 and 2A). Studies of Botrytis pathology commonly differentiate between “primary” and “secondary” (or “spreading”) lesions (van Kan 2006). Primary lesion initiation is believed to be affected by the ability of the fungus to overcome preformed plant defenses. This includes the ability of the fungus to penetrate the leaf surface, relevant to this study as plants were not wounded prior to inoculation, and possibly the extent of plant defense activation by attempted fungal penetration of the leaf surface. However, the loss of penetration-related enzymes, such as cutinase, or the ability to form penetration structures, such as appressoria, have not impaired B. cinerea pathogenicity in experimental settings (van der Vlugt-Bergmans et al. 1997; van Kan et al. 1997; Gourgues et al. 2004). Loci associated with A. thaliana “penetration” mutants, whose defects allow inappropriate penetration of A. thaliana leaves by non-Arabidopsis pathogens, are not colocalized with B. cinerea early lesion QTL detected in this study (Collins et al. 2003; Assaad et al. 2004; Stein et al. 2006). Additionally, pen3 mutants did not show increased susceptibility to a single B. cinerea isolate tested (Stein et al. 2006). As such, it is currently unknown how the initial stages of B. cinerea infection are established. Identifying the molecular basis of early lesion QTL may provide biological insight into A. thaliana responses to early stages of B. cinerea infection.
Transition from primary lesions to secondary (or spreading) lesions requires the pathogen to overcome inducible plant defenses. B. cinerea lesion outgrowth has been shown to be inhibited by accumulation of camalexin (Ferrari et al. 2003, 2007; Kliebenstein et al. 2005). This agrees with the negative association of effects for B. cinerea lesion QTL vs. camalexin accumulation QTL, but it is difficult to infer the direction of causality in a natural variation study (Hammerschmidt 2003). Additional molecular plant defense mechanisms for postinitiation lesion inhibition have not been explicitly described, but are believed to be largely regulated via jasmonate and ethylene signaling (Thomma et al. 1998, 1999; Mengiste et al. 2003; Thaler et al. 2004). Recent work has described a strong effect of wound-induced defenses on spreading of B. cinerea lesions; these defenses include, but are not limited to, increased and more rapid camalexin accumulation (Chassot et al. 2008). Characterizing molecular regulation of a known induced defense, camalexin, may shed light on broader mechanisms of defense against B. cinerea that could be applicable to plant species other than A. thaliana.
Isolate specificity of QTL:
Several QTL studies have used multiple pathogen isolates in attempts to identify genetic elements contributing to durable resistance. These studies have focused on biotrophic pathogens, where qualitative race-specific resistance conferred by major resistance genes is vulnerable to changes in the genetic composition of the pathogen population. The identification of both broad-spectrum and isolate specific resistance QTL suggests that significant variation for mechanisms of pathogenesis exists within many pathogen species and that qualitative isolate-specific resistance conferred by major resistance genes accounts for only a portion of plant defense against naturally variable pathogen populations (Caranta et al. 1997; Arru et al. 2003; Calenge et al. 2004; Talukder et al. 2004; Diener and Ausubel 2005; Jorge et al. 2005). Understanding differences and similarities in mechanisms of partial or quantitative resistance to pathogens of different lifestyles, including biotrophs and necrotrophs, is vital to developing long-term strategies for pathogen control in a complex environment.
Previous efforts to map A. thaliana–B. cinerea interactions using 104 genotypes sampled from the Col × Ler RIL population identified small- to moderate-effect QTL, with no overlapping QTL detected between the 2 isolates tested (Denby et al. 2004). In contrast, the larger population size used in this study (>400 lines) provided greater power to detect QTL and epistatic interactions. The majority of QTL in this study were identified for multiple pathogen isolates, with four QTL identified as affecting A. thaliana interactions with all four B. cinerea isolates tested. These four QTL thus identify the strongest potential contributions of these two A. thaliana genomes to broad-spectrum B. cinerea resistance. Specific tests of two of these isolate-shared QTL regions (Bcr.3.62 and Bcr.4.40) using HIFs supported the overall trends identified in the QTL studies, with 10 of 12 isolates showing directional allelic effects consistent with predicted QTL in the tested regions (Table 1).
Epistasis and quantitative resistance networks:
The identification of significant epistasis controlling plant–pathogen interactions both benefits and complicates this analysis. Epistasis may identify genes that function together in distinct genetic networks, potentially providing valuable insight into function (Alvarez-Castro and Carlborg 2007; Gjuvsland et al. 2007). This potential for epistatic networks to provide molecular network information is highlighted by the recent observation of a two-gene epistatic hybrid necrosis, the inappropriate stimulation of plant disease networks caused by allele-specific interactions in plant disease genes (Bomblies et al. 2007). While the use of epistasis in network analysis has frequently been limited to pairwise analysis, recent metabolomic analysis has shown that higher-order interactions exist and also exhibit allele-specific interactions (Rowe et al. 2008). Our identification of higher-order epistatic networks that control quantitative pathogen resistance in A. thaliana suggests that these QTL may be caused by polymorphism in genes that function in a coordinated defense network. Validation of this epistatic network hypothesis will require cloning the full complement of interacting QTL. Further, detection of three-way epistases suggests that pairwise interactions, such as those producing hybrid necrosis, may present only the simplest and most experimentally accessible examples of interaction; much larger populations will be required to test the true size of epistatic networks.
Epistasis complicates analysis by decreasing power for QTL detection and HIF validation. This is shown by the lack of statistical significance for many of the HIF results in spite of the correct directionality of detected effects. This may be partially caused by the muffling of phenotypic differences between genotypes at the targeted locus by epistatic influences from other loci in the specific HIF background. The use of near isogenic lines, rather than the parentally mixed HIF backgrounds, would not have improved analysis as the parental genotypes are often suboptimal for observing the effects of individual QTL (supplemental Figure 6). Furthermore, the number of QTL and epistatic interactions suggest that significant increases in the size of mapping populations will be required to fully understand quantitative resistance in plant–pathogen interactions (Melchinger et al. 1998; Mihaljevic et al. 2005; Stich et al. 2007). An ideal analysis would employ large structured populations to identify all epistatic interactions and use this information to select optimal genetic backgrounds for either breeding maximally resistant genotypes or guiding future efforts to isolate causal polymorphisms.
Candidate genes and molecular mechanisms:
A rapidly expanding number of associations between specific phenotypes and genetic loci have been detected within the Bay × Sha RIL population. These (and future) studies provide additional data for hypotheses regarding the bases of A. thaliana–B. cinerea interaction. For example, although it has been proposed that plant senescence plays a role in susceptibility to B. cinerea, QTL in this study do not colocalize with senescence QTL previously identified in Bay × Sha RILs (Diaz et al. 2006; Swartzberg et al. 2008). In contrast, Bcr.4.40, identified as influencing lesion size for all B. cinerea isolates tested, occupies the same genomic region as a QTL affecting nitrogen content and total amino acid content (Loudet et al. 2003a,b). This region was recently identified as controlling differences between Bay-0 and Shahdara in accumulation of >50 metabolites; as such, pleiotropic effects of this locus will complicate efforts to predict specific candidate genes controlling differences in B. cinerea susceptibility (Rowe et al. 2008).
Recent studies have identified strong resistance to Botrytis cinerea associated with A. thaliana defects in cuticle production, hypothesized to accelerate plant detection of the pathogen and activation of defense response (Bessire et al. 2007). Comparison of transcript accumulation over the course of B. cinerea infection between cuticle-defective mutants and wild-type A. thaliana identified a number of gene candidates potentially contributing to defense against B. cinerea (Chassot et al. 2007). A cluster of these Botrytis-induced genes, including a chitinase and a trypsin inhibitor, colocalize with Bcr.2.61; however no major sequence polymorphisms at these loci were detected in the recently published genome sequences for the Bay-0 and Shahdara accessions (Clark et al. 2007). Another identified candidate gene, annotated as cysteine lyase COR13 (At4g23600), colocalizes with Bcr.4.40 and sequence data suggests that Shahdara possesses a nonconservative amino acid substitution at this locus. This locus is transcriptionally responsive to jasmonate and wounding and is predicted to influence availability of cysteine (Leon et al. 1998; Jones et al. 2003; Devoto et al. 2005). Cysteine availability has the potential to influence camalexin accumulation through both camalexin biosynthesis and glutathione-mediated signaling (Zook and Hammerschmidt 1997; Parisy et al. 2007). Biochemical testing will be required to determine whether the single amino acid substitution in Shahdara significantly alters activity or regulation of this gene product.
Some of the QTL identified in this study may overlap with B. cinerea resistance QTL previously identified in the A. thaliana Col × Ler RIL population (Denby et al. 2004). In particular, QTL were found at the top and bottom of chromosome 3 in both Col-0 × Ler and Bay-0 × Shahdara, although Bcr.3.62 was found to primarily influence camalexin accumulation in Bay-0 × Shahdara, and that trait was not measured in the Col-0 × Ler RILs (Figure 1). While these QTL shared between populations may represent different polymorphisms, analysis of these QTL regions for genes polymorphic in both pairs of A. thaliana accessions may yield strong candidate genes. Neither study identified a significant QTL at the erecta locus, a QTL influencing plant resistance to a necrotrophic pathogen for which the molecular basis is known (Llorente et al. 2005).
Conclusion:
This study identifies numerous QTL contributing to variation in plant–B. cinerea interactions between the A. thaliana accessions Bay-0 and Shahdara. Genome-scale prediction of large-effect DNA sequence and transcript accumulation polymorphisms differentiating Bay-0 and Shahdara permit an informed approach to selection and investigation of candidate genes in identified QTL regions (Kliebenstein et al. 2006; Clark et al. 2007). While identification of causal polymorphisms for QTL influencing a majority of B. cinerea isolates may provide potential breeding targets for enhanced B. cinerea resistance in related Brassica species, validation and further investigation of isolate-specific QTL may provide valuable insight into variation in mechanisms of pathogenesis within Botrytis cinerea.
Acknowledgments
We thank Michael Kuwabara and Katherine Denby for technical assistance with RIL experiments. We particularly acknowledge the contribution of Bay × Sha HIFs from Olivier Loudet at Institut National de la Recherche Agronomique, Versailles, France. Purified camalexin standards were the generous gift of Jane Glazebrook. Daniel P. Koenig, Charles Hagen, Adam Wentzell, and Rachel Kerwin provided critical reviews of this manuscript prior to submission for publication. We additionally thank two anonymous reviewers for clarifying aspects of this manuscript. The University of California, Davis Department of Plant Sciences and the Henry A. Jastro and Peter J. Shields Research Scholarship Fund provided financial support to H.C.R. during completion of this research.
References
- Abramoff, M. D., P. J. Magelhaes and S. J. Ram, 2004. Image processing with ImageJ. Biophoton. Int. 11 36–42. [Google Scholar]
- Albertini, C., G. Thebaud, E. Fournier and P. Leroux, 2002. Eburicol 14α-demethylase gene (CYP51) polymorphism and speciation in Botrytis cinerea. Mycol. Res. 106 1171–1178. [Google Scholar]
- Alvarez-Castro, J. M., and O. Carlborg, 2007. A unified model for functional and statistical epistasis and its application in quantitative trait loci analysis. Genetics 176 1151–1167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arru, L., E. Francia and N. Pecchioni, 2003. Isolate-specific QTL of resistance to leaf stripe (Pyrenophora graminea) in the ‘Steptoe’ × ‘Morex’ spring barley cross. Theor. Appl. Genet. 106 668–675. [DOI] [PubMed] [Google Scholar]
- Assaad, F. F., J. L. Qiu, H. Youngs, D. Ehrhardt, L. Zimmerli et al., 2004. The PEN1 syntaxin defines a novel cellular compartment upon fungal attack and is required for the timely assembly of papillae. Mol. Biol. Cell 15 5118–5129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bent, A. F., and D. Mackey, 2007. Elicitors, effectors, and R genes: the new paradigm and a lifetime supply of questions. Annu. Rev. Phytopathol. 45 399–436. [DOI] [PubMed] [Google Scholar]
- Bert, P. F., I. Jouan, D. T. de Labrouhe, F. Serre, P. Nicolas et al., 2002. Comparative genetic analysis of quantitative traits in sunflower (Helianthus annuus L.) 1. QTL involved in resistance to Sclerotinia sclerotiorum and Diaporthe helianthi. Theor. Appl. Genet. 105 985–993. [DOI] [PubMed] [Google Scholar]
- Bessire, M., C. Chassot, A. C. Jacquat, M. Humphry, S. Borel et al., 2007. A permeable cuticle in Arabidopsis leads to a strong resistance to Botrytis cinerea. EMBO J. 26 2158–2168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bomblies, K., J. Lempe, P. Epple, N. Warthmann, C. Lanz et al., 2007. Autoimmune response as a mechanism for a Dobzhansky-Muller-type incompatibility syndrome in plants. PloS Biol. 5 1962–1972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Calenge, F., A. Faure, M. Goerre, C. Gebhardt, W. E. Van de Weg et al., 2004. Quantitative trait loci (QTL) analysis reveals both broad-spectrum and isolate-specific QTL for scab resistance in an apple progeny challenged with eight isolates of Venturia inaequalis. Phytopathology 94 370–379. [DOI] [PubMed] [Google Scholar]
- Calpas, J. T., M. N. Konschuh, C. C. Toews and J. P. Tewari, 2001. Virulence and molecular diversity in isolates of Botrytis cinerea recovered from greenhouse tomato in Alberta. Can. J. Plant Pathol. 23 184. [Google Scholar]
- Caranta, C., V. Lefebvre and A. Palloix, 1997. Polygenic resistance of pepper to potyviruses consists of a combination of isolate-specific and broad-spectrum quantitative trait loci. Mol. Plant-Microbe Interact. 10 872–878. [Google Scholar]
- Chassot, C., A. Buchala, H.-J. Schoonbeek, J.-P. Metraux and O. Lamotte, 2008. Wounding of Arabidopsis leaves causes a powerful but transient protection against Botrytis infection. Plant J. 55 555–567. [DOI] [PubMed] [Google Scholar]
- Chassot, C., C. Nawrath and J. P. Metraux, 2007. Cuticular defects lead to full immunity to a major plant pathogen. Plant J. 49 972–980. [DOI] [PubMed] [Google Scholar]
- Clark, R. M., G. Schweikert, C. Toomajian, S. Ossowski, G. Zeller et al., 2007. Common sequence polymorphisms shaping genetic diversity in Arabidopsis thaliana. Science 317 338–342. [DOI] [PubMed] [Google Scholar]
- Collins, N. C., H. Thordal-Christensen, V. Lipka, S. Bau, E. Kombrink et al., 2003. SNARE-protein-mediated disease resistance at the plant cell wall. Nature 425 973–977. [DOI] [PubMed] [Google Scholar]
- Cui, W., R. E. Beever, S. L. Parkes and M. D. Templeton, 2004. Evolution of an osmosensing histidine kinase in field strains of Botryotinia fuckeliana (Botrytis cinerea) in response to dicarboximide fungicide usage. Phytopathology 94 1129–1135. [DOI] [PubMed] [Google Scholar]
- Dangl, J. L., and J. D. G. Jones, 2001. Plant pathogens and integrated defence responses to infection. Nature 411 826–833. [DOI] [PubMed] [Google Scholar]
- de Leon, I. P., J. P. Oliver, A. Castro, C. Gaggero, M. Bentancor et al., 2007. Erwinia carotovora elicitors and Botrytis cinerea activate defense responses in Physcomitrella patens. BMC Plant Biol. 7 52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Denby, K. J., L. J. M. Jason, S. L. Murray and R. L. Last, 2005. ups1, an Arabidopsis thaliana camalexin accumulation mutant defective in multiple defence signalling pathways. Plant J. 41 673–684. [DOI] [PubMed] [Google Scholar]
- Denby, K. J., P. Kumar and D. J. Kliebenstein, 2004. Identification of Botrytis cinerea susceptibility loci in Arabidopsis thaliana. Plant J. 38 473–486. [DOI] [PubMed] [Google Scholar]
- Devoto, A., C. Ellis, A. Magusin, H. S. Chang, C. Chilcott et al., 2005. Expression profiling reveals COI1 to be a key regulator of genes involved in wound- and methyl jasmonate-induced secondary metabolism, defence, and hormone interactions. Plant Mol. Biol. 58 497–513. [DOI] [PubMed] [Google Scholar]
- Diaz, U., V. Saliba-Colombani, O. Loudet, P. Belluomo, L. Moreau et al., 2006. Leaf yellowing and anthocyanin accumulation are two genetically independent strategies in response to nitrogen limitation in Arabidopsis thaliana. Plant Cell Physiol. 47 74–83. [DOI] [PubMed] [Google Scholar]
- Diener, A. C., and F. M. Ausubel, 2005. RESISTANCE TO FUSARIUM OXYSPORUM 1, a dominant Arabidopsis disease-resistance gene, is not race specific. Genetics 171 305–321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doerge, R. W., and G. A. Churchill, 1996. Permutation tests for multiple loci affecting a quantitative character. Genetics 142 285–294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elad, Y., B. Williamson, P. Tudzynski and N. Delen (Editors), 2004. Botrytis: Biology, Pathology and Control. Kluwer Academic Publishers, Dordrecht, The Netherlands.
- Ferrari, S., R. Galletti, C. Denoux, G. De Lorenzo, F. M. Ausubel et al., 2007. Resistance to Botrytis cinerea induced in Arabidopsis by elicitors is independent of salicylic acid, ethylene, or jasmonate signaling but requires PHYTOALEXIN DEFICIENT3. Plant Physiol. 144 367–379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferrari, S., J. M. Plotnikova, G. De Lorenzo and F. M. Ausubel, 2003. Arabidopsis local resistance to Botrytis cinerea involves salicylic acid and camalexin and requires EDS4 and PAD2 but not SID2, EDS5, or PAD4. Plant J. 35 193–205. [DOI] [PubMed] [Google Scholar]
- Finkers, R., P. van den Berg, R. van Berloo, A. ten Have, A. W. van Heusden et al., 2007. Three QTLs for Botrytis cinerea resistance in tomato. Theor. Appl. Genet. 114 585–593. [DOI] [PubMed] [Google Scholar]
- Finkers, R., Y. L. Bai, P. van den Berg, R. van Berloo, F. Meijer-Dekens et al., 2008. Quantitative resistance to Botrytis cinerea from Solanum neorickii. Euphytica 159 83–92. [Google Scholar]
- Gabriel, D. W., 1999. Why do pathogens carry avirulence genes? Physiol. Mol. Plant Pathol. 55 205–214. [Google Scholar]
- Gjuvsland, A. B., B. J. Hayes, T. H. E. Meuwissen, E. Plahte and S. W. Omholt, 2007. Nonlinear regulation enhances the phenotypic expression of trans-acting genetic polymorphisms. BMC Syst. Biol. 1 32–43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glazebrook, J., 2005. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol. 43 205–227. [DOI] [PubMed] [Google Scholar]
- Glazebrook, J., and F. M. Ausubel, 1994. Isolation of phytoalexin-deficient mutants of Arabidopsis thaliana and characterization of their interactions with bacterial pathogens. Proc. Natl. Acad. Sci. USA 91 8955–8959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gourgues, M., A. Brunet-Simon, M. H. Lebrun and C. Levis, 2004. The tetraspanin BcPls1 is required for appressorium-mediated penetration of Botrytis cinerea into host plant leaves. Mol. Microbiol. 51 619–629. [DOI] [PubMed] [Google Scholar]
- Hammerschmidt, R., 2003. Phytoalexin accumulation: response or defense. Physiol. Mol. Plant Pathol. 62 125–126. [Google Scholar]
- Jirage, D., T. L. Tootle, T. L. Reuber, L. N. Frost, B. J. Feys et al., 1999. Arabidopsis thaliana PAD4 encodes a lipase-like gene that is important for salicylic acid signaling. Proc. Natl. Acad. Sci. USA 96 13583–13588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones, J. D. G., and J. L. Dangl, 2006. The plant immune system. Nature 444 323–329. [DOI] [PubMed] [Google Scholar]
- Jones, P. R., T. Manabe, M. Awazuhara and K. Saito, 2003. A new member of plant CS-lyases: a cystine lyase from Arabidopsis thaliana. J. Biol. Chem. 278 10291–10296. [DOI] [PubMed] [Google Scholar]
- Jorge, V., A. Dowkiw, P. Faivre-Rampant and C. Bastien, 2005. Genetic architecture of qualitative and quantitative Melampsora larici-populina leaf rust resistance in hybrid poplar: genetic mapping and QTL detection. New Phytol. 167 113–127. [DOI] [PubMed] [Google Scholar]
- Keurentjes, J. J. B., J. Y. Fu, I. R. Terpstra, J. M. Garcia, G. van den Ackerveken et al., 2007. Regulatory network construction in Arabidopsis by using genome-wide gene expression quantitative trait loci. Proc. Natl. Acad. Sci. USA 104 1708–1713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim, H. S., and B. W. Diers, 2000. Inheritance of partial resistance to Sclerotinia stem rot in soybean. Crop Sci. 40 55–61. [Google Scholar]
- Kliebenstein, D. J., H. C. Rowe and K. J. Denby, 2005. Secondary metabolites influence Arabidopsis/Botrytis interactions: variation in host production and pathogen sensitivity. Plant J. 44 25–36. [DOI] [PubMed] [Google Scholar]
- Kliebenstein, D. J., M. A. L. West, H. van Leeuwen, O. Loudet, R. W. Doerge et al., 2006. Identification of QTL controlling gene expression networks defined a priori. BMC Bioinformatics 7 308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leon, J., E. Rojo, E. Titarenko and J. J. Sanchez-Serrano, 1998. Jasmonic acid-dependent and -independent wound signal transduction pathways are differentially regulated by Ca2+/calmodulin in Arabidopsis thaliana. Mol. Gen. Genet. 258 412–419. [DOI] [PubMed] [Google Scholar]
- Llorente, F., C. Alonso-Blanco, C. Sanchez-Rodriguez, L. Jorda and A. Molina, 2005. ERECTA receptor-like kinase and heterotrimeric G protein from Arabidopsis are required for resistance to the necrotrophic fungus Plectosphaerella cucumerina. Plant J. 43 165–180. [DOI] [PubMed] [Google Scholar]
- Loudet, O., S. Chaillou, C. Camilleri, D. Bouchez and F. Daniel-Vedele, 2002. Bay-0 × Shahdara recombinant inbred line population: a powerful tool for the genetic dissection of complex traits in Arabidopsis. Theor. Appl. Genet. 104 1173–1184. [DOI] [PubMed] [Google Scholar]
- Loudet, O., S. Chaillou, A. Krapp and F. Daniel-Vedele, 2003. a Quantitative trait loci analysis of water and anion contents in interaction with nitrogen availability in Arabidopsis thaliana. Genetics 163 711–722. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loudet, O., S. Chaillou, P. Merigout, J. Talbotec and F. Daniel-Vedele, 2003. b Quantitative trait loci analysis of nitrogen use efficiency in Arabidopsis. Plant Physiol. 131 345–358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loudet, O., V. Gaudon, A. Trubuil and F. Daniel-Vedele, 2005. Quantitative trait loci controlling root growth and architecture in Arabidopsis thaliana confirmed by heterogeneous inbred family. Theor. Appl. Genet. 110 742–753. [DOI] [PubMed] [Google Scholar]
- Ma, S., and H. J. Bohnert, 2008. Gene networks in Arabidopsis thaliana for metabolic and environmental functions. Mol. Biosyst. 4 199–204. [DOI] [PubMed] [Google Scholar]
- Maxwell, J. J., M. A. Brick, P. F. Byrne, H. F. Schwartz, X. Shan et al., 2007. Quantitative trait loci linked to white mold resistance in common bean. Crop Sci. 47 2285–2294. [Google Scholar]
- Melchinger, A. E., H. F. Utz and C. C. Schön, 1998. Quantitative trait locus (QTL) mapping using different testers and independent population samples in maize reveals low power of QTL detection and large bias in estimates of QTL effects. Genetics 149 383–403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mengiste, T., X. Chen, J. Salmeron and R. Dietrich, 2003. The BOTRYTIS SUSCEPTIBLE1 gene encodes an R2R3MYB transcription factor protein that is required for biotic and abiotic stress responses in Arabidopsis. Plant Cell 15 2551–2565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Micic, Z., V. Hahn, E. Bauer, C. C. Schon, S. J. Knapp et al., 2004. QTL mapping of Sclerotinia midstalk-rot resistance in sunflower. Theor. Appl. Genet. 109 1474–1484. [DOI] [PubMed] [Google Scholar]
- Mihaljevic, R., H. F. Utz and A. E. Melchinger, 2005. No evidence for epistasis in hybrid and per se performance of elite European flint maize inbreds from generation means and QTL analyses. Crop Sci. 45 2605–2613. [Google Scholar]
- Mikkelsen, M. D., B. L. Petersen, E. Glawischnig, A. B. Jensen, E. Andreasson et al., 2003. Modulation of CYP79 genes and glucosinolate profiles in Arabidopsis by defense signaling pathways. Plant Physiol. 131 298–308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nafisi, M., S. Goregaoker, C. J. Botanga, E. Glawischnig, C. E. Olsen et al., 2007. Arabidopsis cytochrome P450 monooxygenase 71A13 catalyzes the conversion of indole-3-acetaldoxime in camalexin synthesis. Plant Cell 19 2039–2052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niks, R. E., and D. Rubiales, 2002. Potentially durable resistance mechanisms in plants to specialised fungal pathogens. Euphytica 124 201–216. [Google Scholar]
- Nordborg, M., T. T. Hu, Y. Ishino, J. Jhaveri, C. Toomajian et al., 2005. The pattern of polymorphism in Arabidopsis thaliana. PLoS Biol. 3 e196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parisy, V., B. Poinssot, L. Owsianowski, A. Buchala, J. Glazebrook et al., 2007. Identification of PAD2 as a gamma-glutamylcysteine synthetase highlights the importance of glutathione in disease resistance of Arabidopsis. Plant J. 49 159–172. [DOI] [PubMed] [Google Scholar]
- Park, S. O., D. P. Coyne, J. R. Steadman and P. W. Skroch, 2001. Mapping of QTL for resistance to white mold disease in common bean. Crop Sci. 41 1253–1262. [Google Scholar]
- Perchepied, L., T. Kroj, M. Tronchet, O. Loudet and D. Roby, 2006. Natural variation in partial resistance to Pseudomonas syringae is controlled by two major QTLs in Arabidopsis thaliana. PLoS One 1 e123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prins, T. W., P. Tudzynski, A. von Tiedemann, B. Tudzynski, A. ten Have et al., 2000. Infection strategies of Botrytis cinerea and related necrotrophic pathogens, pp. 32–64 in Fungal Pathology, edited by J. Kronstad. Kluwer Academic Publishers, Dordrecht, The Netherlands.
- Qutob, D., B. Kemmerling, F. Brunner, I. Kufner, S. Engelhardt et al., 2006. Phytotoxicity and innate immune responses induced by Nep1-like proteins. Plant Cell 18 3721–3744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ren, D., Y. Liu, K.-Y. Yang, L. Han, G. Mao et al., 2008. A fungal-responsive MAPK cascade regulates phytoalexin biosynthesis in Arabidopsis. Proc. Natl. Acad. Sci. USA 105 5638–5643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robert-Seilaniantz, A., L. Navarro, R. Bari and J. D. Jones, 2007. Pathological hormone imbalances. Curr. Opin. Plant Biol. 10 372–379. [DOI] [PubMed] [Google Scholar]
- Rowe, H. C., and D. J. Kliebenstein, 2007. Elevated genetic variation within virulence-associated Botrytis cinerea polygalacturonase loci. Mol. Plant-Microbe Interact. 20 1126–1137. [DOI] [PubMed] [Google Scholar]
- Rowe, H. C., B. G. Hansen, B. A. Halkier and D. J. Kliebenstein, 2008. Biochemical networks and epistasis shape the Arabidopsis thaliana metabolome. Plant Cell 20 1199–1216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sellam, A., B. Iacomi-Vasilescu, P. Hudhomme and P. Simoneau, 2007. In vitro antifungal activity of brassinin, camalexin and two isothiocyanates against the crucifer pathogens Alternaria brassicicola and Alternaria brassicae. Plant Pathol. 56 296–301. [Google Scholar]
- Sharbel, T. F., B. Haubold and T. Mitchell-Olds, 2000. Genetic isolation by distance in Arabidopsis thaliana: biogeography and postglacial colonization of Europe. Mol. Ecol. 9 2109–2118. [DOI] [PubMed] [Google Scholar]
- Shindo, C., G. Bernasconi and C. S. Hardtke, 2007. Natural genetic variation in Arabidopsis: tools, traits and prospects for evolutionary ecology. Ann. Bot. 99 1043–1054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stein, M., J. Dittgen, C. Sanchez-Rodriguez, B. H. Hou, A. Molina et al., 2006. Arabidopsis PEN3/PDR8, an ATP binding cassette transporter, contributes to nonhost resistance to inappropriate pathogens that enter by direct penetration. Plant Cell 18 731–746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stich, B., J. M. Yu, A. E. Melchinger, H. P. Piepho, H. F. Utz et al., 2007. Power to detect higher-order epistatic interactions in a metabolic pathway using a new mapping strategy. Genetics 176 563–570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swartzberg, D., B. Kirshner, D. Rav-David, Y. Elad and D. Granot, 2008. Botrytis cinerea induces senescence and is inhibited by autoregulated expression of the IPT gene. Eur. J. Plant Pathol. 120 289–297. [Google Scholar]
- Symonds, V. V., A. V. Godoy, T. Alconada, J. F. Botto, T. E. Juenger et al., 2005. Mapping quantitative trait loci in multiple populations of Arabidopsis thaliana identifies natural allelic variation for trichome density. Genetics 169 1649–1658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Talukder, Z. I., D. Tharreau and A. H. Price, 2004. Quantitative trait loci analysis suggests that partial resistance to rice blast is mostly determined by race-specific interactions. New Phytol. 162 197–209. [Google Scholar]
- Thaler, J. S., B. Owen and V. J. Higgins, 2004. The role of the jasmonate response in plant susceptibility to diverse pathogens with a range of lifestyles. Plant Physiol. 135 530–538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomma, B. P. H. J., K. Eggermont, I. A. M. A. Penninckx, B. Mauch-Mani, R. Vogelsang et al., 1998. Separate jasmonate-dependent and salicylate-dependent defense-response pathways in Arabidopsis are essential for resistance to distinct microbial pathogens. Proc. Natl. Acad. Sci. USA 95 15107–15111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomma, B. P. H. J., I. Nelissen, K. Eggermont and W. F. Broekaert, 1999. Deficiency in phytoalexin production causes enhanced susceptibility of Arabidopsis thaliana to the fungus Alternaria brassicicola. Plant J. 19 163–171. [DOI] [PubMed] [Google Scholar]
- Tonsor, S. J., C. Alonso-Blanco and M. Koornneef, 2005. Gene function beyond the single trait: natural variation, gene effects, and evolutionary ecology in Arabidopsis thaliana. Plant Cell Environ. 28 2–20. [Google Scholar]
- Vales, M. I., C. C. Schon, F. Capettini, X. M. Chen, A. E. Corey et al., 2005. Effect of population size on the estimation of QTL: a test using resistance to barley stripe rust. Theor. Appl. Genet. 111 1260–1270. [DOI] [PubMed] [Google Scholar]
- van der Vlugt-Bergmans, C. J. B., C. A. M. Wagemakers and J. A. L. van Kan, 1997. Cloning and expression of the cutinase A gene of Botrytis cinerea. Mol. Plant-Microbe Interact. 10 21–29. [DOI] [PubMed] [Google Scholar]
- van Kan, J. A. L., 2006. Licensed to kill: the lifestyle of a necrotrophic plant pathogen. Trends Plant Sci. 11 247–253. [DOI] [PubMed] [Google Scholar]
- van Kan, J. A. L., J. W. vant Klooster, C. A. Wagemakers, D. C. Dees and C. J. van der Vlugt-Bergmans, 1997. Cutinase A of Botrytis cinerea is expressed, but not essential, during penetration of gerbera and tomato. Mol. Plant-Microbe Interact. 10 30–38. [DOI] [PubMed] [Google Scholar]
- Wentzell, A. M., H. C. Rowe, B. G. Hansen, C. Ticconi, B. A. Halkier et al., 2007. Linking metabolic QTL with network and cis-eQTL controlling biosynthetic pathways. PloS Genet. 3 1687–1701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- West, M. A. L., K. Kim, D. J. Kliebenstein, H. van Leeuwen, R. W. Michelmore et al., 2007. Global eQTL mapping reveals the complex genetic architecture of transcript-level variation in Arabidopsis. Genetics 175 1441–1450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williamson, B., B. Tudzynsk, P. Tudzynski and J. A. L. van Kan, 2007. Botrytis cinerea: the cause of grey mould disease. Mol. Plant Pathol. 8 561–580. [DOI] [PubMed] [Google Scholar]
- Young, N. D., 1996. QTL mapping and quantitative disease resistance in plants. Annu. Rev. Phytopathol. 34 479–501. [DOI] [PubMed] [Google Scholar]
- Zeng, Z.-B., C.-H. Kao and C. J. Basten, 1999. Estimating the genetic architecture of quantitative traits. Genet. Res. 74 279–289. [DOI] [PubMed] [Google Scholar]
- Zhao, J. W., and J. L. Meng, 2003. Genetic analysis of loci associated with partial resistance to Sclerotinia sclerotiorum in rapeseed (Brassica napus L.). Theor. Appl. Genet. 106 759–764. [DOI] [PubMed] [Google Scholar]
- Zhou, N., T. L. Tootle and J. Glazebrook, 1999. Arabidopsis PAD3, a gene required for camalexin biosynthesis, encodes a putative cytochrome P450 monooxygenase. Plant Cell 11 2419–2428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zimmerli, L., J.-P. Metraux and B. Mauch-Mani, 2001. β-aminobutyric acid-induced protection of Arabidopsis against the necrotrophic fungus Botrytis cinerea. Plant Physiol. 126 517–523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zook, M., and R. Hammerschmidt, 1997. Origin of the thiazole ring of camalexin, a phytoalexin from Arabidopsis thaliana. Plant Physiol. 113 463–468. [DOI] [PMC free article] [PubMed] [Google Scholar]