

Abstract
Across the boreal forest, fire is the main disturbance factor and driver of ecosystem changes. In this study, we reconstructed a long-term, spatially explicit fire history of a forest-tundra region in northeastern Canada. We hypothesized that current occupation of similar topographic and edaphic sites by tundra and forest was the consequence of cumulative regression with time of forest cover due to compounding fire and climate disturbances. All fires were mapped and dated per 100 year intervals over the last 2000 years using several fire dating techniques. Past fire occurrences and post-fire regeneration at the northern forest limit indicate 70% reduction of forest cover since 1800 yr BP and nearly complete cessation of forest regeneration since 900 yr BP. Regression of forest cover was particularly important between 1500s–1700s and possibly since 900 yr BP. Although fire frequency was very low over the last 100 years, each fire event was followed by drastic removal of spruce cover. Contrary to widespread belief of northward boreal forest expansion due to recent warming, lack of post-fire recovery during the last centuries, in comparison with active tree regeneration more than 1000 years ago, indicates that the current climate does not favour such expansion.
Keywords: boreal forest, forest tundra, subarctic, fire, climate change, fire history
1. Introduction
The northern part of the North American boreal forest corresponds to the forest-tundra ecotone where trees and forests reach their northernmost distribution limits (Rowe 1972; Timoney et al. 1992; Payette et al. 2001). The most extensive forest-tundra areas in North America are located on the Canadian Precambrian Shelf from the Northwest Territories to the Labrador coast. The vegetation of the forest tundra is composed of lichen-shrub tundra communities on hilltops and open forest stands on lower slopes and depressions. This rather common spatial pattern of tundra and forest communities across the ecotone is the result of long-term interactions between fire disturbance and climate (Payette & Gagnon 1985; Asselin & Payette 2006). Similar connections between fire, vegetation and climate are known for other large ecotones like the prairie-forest border in Central North America (Grimm 1983, 1984; Umbanhowar 2004).
The reconstruction of the past dynamics of the boreal forest is based primarily on the stratigraphic analysis of organic cores from lake and peat sediments (Patterson et al. 1987; Ritchie 1987; Wein et al. 1987; MacDonald et al. 1991; Kuhry 1994; Earle et al. 1996; Whitlock & Millspaugh 1996; Tryterud 2003; Whitlock & Bartlein 2004). Most studies focussing on the historical role of fire in vegetation development have used indirect fire evidence based on pollen-slide and thin-section methods applied predominantly to lake sediments (Cwynar 1978; Swain 1978, 1980; Terasmae & Weeks 1979; Green 1982; Patterson et al. 1987; Clark 1988; MacDonald et al. 1991; Clark & Royall 1996; Carcaillet et al. 2001; Asselin & Payette 2005a). These methods give a rather crude assessment of past fire occurrences in time and space and their impact on ecosystems throughout postglacial times. Charcoal fragments from lake sediments are useful to yield a general account of fire presence and influence over time. But the distribution and abundance of microscopic and macroscopic charcoal fragments in a lake core are not reliable enough parameters to identify nearby individual past fire events at both the site and regional scales and, in particular, to calculate fire frequencies as it is sometimes suggested (Carcaillet et al. 2001; Anderson et al. 2006; Stähli et al. 2006).
On the other hand, long-term and spatially explicit fire reconstructions are rare (Niklasson & Granström 2000), difficult to realize in the field given the phasing out of fire evidence with time (Heinselman 1973; Tande 1979; Payette et al. 1989a,b; Johnson & Gutsell 1994; Bergeron et al. 2001, 2004), and generally do not go beyond the last 600–800 years (Engelmark 1984; Niklasson & Granström 2000). It is possible to reconstruct a spatially explicit, long-term fire history of a given area when long-lived tree species are dominant (LaMarche 1969; Engelmark 1984; Lara & Villalba 1993; Swetnam 1993; Niklasson & Granström 2000). Except under exceptional circumstances, like in southwest USA where several trees are living for thousands of years (Swetnam 1993; Lloyd & Graumlich 1997), most long-term fire histories rely on other direct fire evidence like macroscopic charcoal on and in mineral soils (Filion 1984; Payette & Gagnon 1985; Filion et al. 1991; Desponts & Payette 1993; Lavoie & Payette 1996; Carcaillet 2001; Asselin & Payette 2005b; Talon et al. 2005). The reconstruction of long-term fire histories in a given area of a size far greater than the classical lake catchment necessitates time-consuming field research using several complementary methods, including radiocarbon dating of charcoal, tree-ring dating and sometimes allometric scaling of plant traits, to accurately date ancient and recent fire events. Forest-tundra environments are among the best templates to analyse the spatiotemporal development of forest vegetation over several hundred years in an area of sufficient but representative size, at least several tens of square kilometres.
In this study, our primary objective was to reconstruct a long-term, spatially explicit fire history of the northernmost boreal forest ecosystems in northeastern Canada in the context of the dynamics of the global circumboreal forest. Owing to the dual and contrasted occupation of tundra and forest communities in the same forest-tundra area and in the same topographic and edaphic setting, we were also interested (i) to evaluate the climatic impact of fire in time and space on the regeneration of the boreal forest at its northernmost limit and (ii) to determine the inception and expansion of tundra communities in a typical North American forest-tundra landscape. It was hypothesized that the current dual occupation of similar sites by tundra and forest communities was the consequence of the cumulative regression with time of the forest cover due to compounding fire and climate disturbances. To do so, the work necessitated several years of field investigations to map exhaustively and to accurately date the occurrence of all individual fires per 100 year intervals over the last 2000 years. To realize such a detailed account of the long-term behaviour of the northernmost boreal forest, we documented and analysed the fire record of a northern boreal forest-tundra area of 40 km2 within the Rivière Boniface watershed in northeastern Canada (57°45′ N, 76° W; figure 1).
Figure 1.

Location of the study area.
2. Material and methods
The study area harbours the northernmost forest vegetation along the eastern coast of Hudson Bay. Black spruce (Picea mariana [Mill.] B. S. P.) is the dominant tree species in the area with only a few eastern larch (Larix laricina [DuRoi] K. Koch) trees growing in mesic and wet soils (Peňalba & Payette 1997). Spruce-moss stands are the most important forest type in depressions, whereas lichen-spruce woodlands and krummholz (pygmy forests; distorted spruce less than 2.5 m high) are distributed on slopes and exposed summits along with lichen-heath-dwarf birch (Betula glandulosa Michx.) communities (figure 2). Woodlands are restricted to lowlands not exceeding 135 m above sea level (a.s.l.) and krummholz extend to 165 m a.s.l. Krummholz are damaged by snowdrift action although these stands regenerated after fire when conditions were favourable for seedling establishment (Payette et al. 1989a,b; Asselin & Payette 2005b). Woodland and krummholz cover 30% of the study area, whereas lichen-tundra communities occupy the remaining well-drained sites up to the highest altitude, approximately 215 m a.s.l. Several sandy lichen-tundra sites along the river bank are also occupied by sand dunes, largely controlled by fire (Saint-Laurent & Filion 1992). The Boniface area has been occupied in the recent past by small groups of Nunamiut, i.e. Inuit living in interior Ungava peninsula (Vézinet 1980; Payette et al. 2004), as evidenced by summer and winter camps distributed on several headlands along the river.
Figure 2.

Distribution of present dominant vegetation types according to altitude in the Rivière Boniface area. Lichen-spruce woodlands in black, lichen-spruce krummholz in medium grey and lichen-heath tundras in light grey.
The systematic analysis of fire evidence included the exhaustive sampling of all the well-drained sites within the 40 km2 study area, i.e. approximately 300 contiguous, well-drained hill sites ranging between 115 and 215 m a.s.l. In this study, a site (or a fire site) was defined as an individual hill (or rarely an alluvial terrace) delineated by wetland vegetation occupying confined depressions, small or large valleys, or mires (peatlands). The hills are Precambrian outcrops covered by thin glacial and fluvial-marine sediments where podzolic soils develop in gravelly loams. Surface analysis for the reconstruction of fire activity included the dating of all fires at each site and evaluation of spruce cover (cover classes of <1%, 1–10%, 10–20%, 20–30%, etc.). Lichen-spruce woodland and lichen-spruce krummholz are two dominant vegetation types having more than 10% spruce cover, whereas lichen-tundra is a spatially dominant vegetation type having less than 10% spruce cover. An examination of all the fire evidence was done at each site using a stratigraphical approach. The latter included a survey of the surface and examination of the uppermost mineral soil for sampling and dating of each fire in order to produce a stand-origin map (Heinselman 1973; Johnson & Gutsell 1994). Two contrasted situations based on the distribution of woodlands, krummholz and tundras were encountered during sampling of fire evidence, which necessitated the selection of several dating techniques. Whereas recent fires are identified rather easily from fire scars and charred wood at the soil surface, old post-fire stands show evidence of fire from charcoal located on, within, or beneath the modern organic soil horizon. An evaluation of the former abundance of black spruce in lichen-tundra sites, based on distribution, number and growth form of charred standing or fallen dead stems was done in order to deduce the presence of woodland or krummholz stands at the moment of the last fire event. This evaluation was used to measure forest survival over time and correlative success of post-fire tree regeneration.
(a) Fire dating
The following methods were used to date the last fire and sometimes the previous fire occurring at a site: dating of fire scars, dating of post-fire trees, dating of pre-fire trees, radiocarbon dating of charcoal and clonometry.
Fire scars on black spruce stems were used to date recent fires. Fire scars were rare because subarctic spruces usually display dense foliage at the stem base that is susceptible to burning completely during fire. Dating of relatively old fires (back to 450 years ago) was possible using fire scars on living and standing dead stems. Fire-scarred trees were cross-dated based on an accurately dated sequence of light rings developed in this area (Filion et al. 1986). Exceptionally old-aged fire scars are found only on long-lived Gymnosperms (Swetnam 1993).
Dating of post-fire trees from tree-ring counts is a common practice in sites where the time elapsed since fire is shorter than the lifespan of regenerated tree species (Heinselman 1973). This technique proved useless most of the time because the large majority of post-fire spruces were growing as stunted shrubs. Instead of forming normal single-stemmed trees, most post-fire black spruces in the Rivière Boniface area are growing as stunted individuals in the form of multi-stemmed clones (Bégin & Filion 1999; Laberge et al. 2001), spreading in all directions when soil topography is uniform, thus forming circular, elliptic or elongated features. Every post-fire spruce clone increases in size with time since the last fire date, but the centre of the clone opens with time due to death of the oldest stems (Laberge et al. 2001). In a few cases, it has been possible to approximately date the last fire occurring at a site based on dating of the oldest stems at the centre of clones (Simard 1996; Laberge et al. 2001). To approximate the fire date, we developed a technique called clonometry, based on size (perimeter) of post-fire clones used as a surrogate for true age as commonly used in lichenometry (Innes 1985). An allometric relationship was developed between clone size and time-since-fire in sites where fire was dated by fire scars and dead pre-fire trees (recent fires) and charcoal (old fires). Clones used in the model were all genuine genotypes identified with genetic markers (Laberge et al. 2000). We extended our original dataset to include the perimeter of 809 clones from 105 different sites. A model of clone growth with time was constructed and based on the relation between time-since-fire dates of tree-ring dated fires (recent) and 14C-dated fires (old) and the perimeter of the two largest clones in each site where genetic markers were used to identify genuine clones (figure 3). On a millennial time scale, the size of clones gives a reliable approximation of time-since-fire. The size structure of all post-fire clones per site was constructed for each of the 105 sites. Perimeter distribution of post-fire clones (figure 4) is similar to diameter distribution of post-fire trees used in standard forest inventories (Oliver & Larson 1996). The number of clones per size class gives a portrait of recruitment success of black spruce with time-since-fire. The model of clone growth has been used for testing the validity of radiocarbon charcoal dates for the determination of time-since-fire dates and delineation of old fires in a given area. For example, it is highly probable that clones with a perimeter of greater than 60 m are not growing in sites disturbed by recent or relatively recent fires. In contrast, the probability is low for finding only small clones with a circumference less than 20 m occupying a given old-fire site, for example more than 500 years old.
Figure 3.

Growth model of black spruce clones (two clones per site identified with genetic markers and clone morphology) according to time-since-fire based on tree ring and 14C dating of fire events.
Figure 4.

Size structure of (a) 50, (b) 250, (c) ca 450 and (d) ca 1000 years old post-fire stands based on distribution of number of spruce clones per 10 m perimeter classes.
The date of mortality of pre-fire trees was used to evaluate the age of the last fire to have occurred in 130 stands where charred trees were lying on the ground (figure 5). More than 500 burned stems were cross-dated using the light-ring chronology constructed by Filion et al. (1986). Tree-ring dating of pre-fire stems was also checked with Cofecha (Holmes 1983). The majority of stems dated corresponded to trees killed by fire. This was particularly the case for a number of well-preserved charred spruce stems found in small lakes and ponds. Depending on the time elapsed since the last fire, a large number of pre-fire spruce stems were at a varying stage of decomposition and thus have lost an unknown number of annual rings. For trees alive at the time of fire, the last dated tree ring on each stem gives a maximum age of fire. Also, an unknown proportion of burned stems were trees already dead when fire occurred. Pooling all the cross-dated pre-fire stems at a particular site gives a better approximation of maximum time-since-fire in the form of a bell-shaped or skewed-to-left distribution of the number of residual, cross-dated pre-fire stems going back in time from the actual date of fire (figure 5). Plotting the respective distribution of all cross-dated pre-fire stems for all the sites burned by a given fire event and for all the fire events produces a temporal sequence of successive bell-shaped distributions corresponding to all the fire events (figure 6).
Figure 5.

Mortality structure of pre-fire trees based on date of the outermost tree ring of each spruce stem lying on the ground. Age determination of fire occurrence in 50 year class is based on the youngest charred stems sampled at each site. (a–d) Different sites burned at ca 1550–1600, 1700–1750, 1800–1850 and 1900–1950, respectively, with the temporal distribution of the number of tree-ring cross-dated spruce stems.
Figure 6.

Dating of past fires based on distribution of the date of mortality of all cross-dated pre-fire trees as indicated by the outermost tree ring formed on each stem (n=500). All the depressions separating the bell-shaped distribution of pre-fire trees (highlighted by dotted curves) correspond to fire events (arrows indicate scar-dated fires). The graph illustrates the temporal sequence of successive bell-shaped distributions corresponding to all the fire events.
Time-since-fire of all post-fire woodlands and krummholz and several lichen-tundra sites was determined based on 203 radiocarbon-dated charcoal (or wood) fragments located beneath the modern organic topsoil (appendix A). Charcoal samples and other organic samples were cleaned of rootlets and other extraneous plant materials before processing for radiocarbon dating at the Centre d'études nordiques 14C Laboratory (Université Laval, Québec City, PQ, Canada; UL), IsoTrace Laboratory (University of Toronto, Toronto, ON, Canada; TO) and Beta Analytic, Incorporated (Miami, FL, USA; BETA). All radiocarbon dates were calibrated from a probability distribution based on Stuiver et al.'s (1998) program and Calib Rev 5.0.2. Macroscopic charcoal (≥0.5 mm) in a soil is direct evidence for local fire influence (Ohlson & Tryterud 2000). All charcoal samples were pieces of individual spruce branches and stems which were probably dead when fire occurred, indicating a charcoal age older than the age of the fire proper. Charcoal pieces were sampled generally at the summit of each hill site. When possible, time-since-fire dates determined from charcoal dates were validated by charred wood from the same sites, either submerged in nearby small lakes or buried or surfacing in mires and having the same radiometric age. Contiguous hill sites with similar charcoal dates (±1 s.d.) and clone-size distribution suggested the occurrence of the same fire event.
(b) Fire statistics
The surface covered by individual fires was reported on 1 : 40 000 air photographs and measured with MapInfo Professional (v. 7.5, MapInfo Corporation 2005). Fire frequency (i.e. the number of fires per year) was calculated for the last 100 (twentieth century), 200 (nineteenth and twentieth centuries together) and 300 years (eighteenth, nineteenth and twentieth centuries together), respectively. The calculation of these fire frequencies was done because it has been possible to identify, map and measure the surface of all individual fires that occurred since AD 1700. The fire rotation period (Heinselman 1973), or fire cycle (Van Wagner 1978), also has been calculated for the last 100, 200 and 300 years, respectively. It is defined as the number of years necessary to burn at least one time an area equal to the entire study area. The fire rotation period has been calculated based on fire date and fire-area data. The calculation of the fire rotation period for the last 100, 200 and 300 years was possible because the contour of each individual fire was delineated clearly with no obvious overlapping.
3. Results and discussion
(a) Fire-free sites
According to charcoal found at the surface or beneath the organic topsoil, 90% of the surface of all the study sites burned at least one time recently or over the last millennia. The remaining 10% is an exceptionally fire-free area composed of old-growth, lichen-Empetrum nigrum woodlands and krummholz (figure 7a,b), probably much older than 3000 yr according to 14C dates of the organic matter at the base of modern organic horizons (Payette & Morneau 1993). Each fire-free stand was visited several times and the organic horizons at the soil surface were examined in detail for evidence of charcoal fragments. The absence of charcoal or any other sign of fire was confirmed independently by several observers, which suggests that fire never occurred at the sites since initial forest or krummholz establishment or that fire occurrence was too old to be detected with current field techniques at the soil surface. The oldest charcoal date at the soil surface recorded in the Rivière Boniface area is from charcoal partly buried in mineral soil (3200 yr BP according to Payette & Morneau 1993). Similar 14C dates (3190 and 3490 yr BP) were obtained only from charcoal buried in sand dunes along the Rivière Boniface (Saint-Laurent & Filion 1992).
Figure 7.

(a) Old-growth lichen-spruce woodland more than 2000 years old. Woodland at equilibrium with all-sized and all-aged dead and living spruce trees. (b) Old-growth lichen-spruce krummholz more than 2000 years old with all-sized and all-aged spruce shrubs. Dead tree stems on the ground were growing in the 1400s and 1500s when growth conditions were more favourable than at present (Payette et al. 1989). (c) Clonal black spruce, ca 100 year old, growing in a lichen-heath tundra site. (d) Clonal black spruce, ca 450 year old, growing in a lichen-heath tundra site. Mosaic of post-fire old-growth woodlands and lichen-heath tundras in the background. (e) Pre-fire spruce stems on the ground associated with fire 450 years old. Cross-dating of pre-fire trees was used to date past fire events when fire scar and post-fire tree evidence was lacking. (f) Lichen-heath tundras caused by deforesting fire 450 years ago adjacent to fire-free lichen-spruce woodlands.
(b) Spatiotemporal distribution of burned stands at the landscape scale
Dating and delineation of each fire recorded in the remaining 90% of the surface of the studied sites have been a continuous ecological challenge owing to the long period of time considered in this study and the nature and complexity of old-growth subarctic vegetation. Recent fires are generally easy to date and circumscribe in the landscape, more particularly in the forest tundra where fire frequency, size and overlapping are greatly reduced in comparison with the same fire metrics in the zone of the continuous boreal forest (Payette et al. 1989a,b; Johnson 1992; Arseneault 2001). Older fires are less easy to circumscribe owing to the incidence of more recent fires that have obliterated their tracks. In a region like the Rivière Boniface, where fire activity is greatly reduced owing to humid and cold conditions associated with the climatic influence of Hudson Bay, it is possible, however, to go back in time without losing too much paleoecological information on the local and regional fire history. The use of complementary dating techniques described in §2 along with the systematic comparison of clone-size distributions between all post-fire lichen-tundra sites (figure 7c,d) helped to discriminate individual fires, at least back to the last 1000 years and also fires older than 1000 radiocarbon years typically recorded in all the woodlands and krummholz of the study area. The interpretation of radiocarbon dates of charcoal material to infer fire dates requires knowledge on the long-term dynamics of old-growth lichen-spruce woodlands. Three important features were considered in this respect before determining the age of a post-fire stand based on radiocarbon-dated charcoal: the origin of the wood material dated (figure 7e), the distribution of charcoal at the soil surface, and the distribution and abundance of black spruce along soil toposequences (figure 7f).
The origin of the wood material dated is closely associated with the nature and age of the nursery stand. Charcoal is produced generally from dead, dry wood (twigs, branches or stems) still attached to a living tree or shrub or lying on the soil surface. Because a radiocarbon date of wood charcoal indicates the time when the wood was living, not the time when fire occurred, it is necessary to know when the dated wood died before the fire conflagration. In sites with frequent fires, radiocarbon dates of charcoal approximate actual fire dates, as in dry boreal environments where fire frequency is relatively high (Filion 1984). This is not necessarily the case in sites with less frequent fires. The problem of age determination of fire based on wood charcoal has been mentioned earlier (Payette & Lajeunesse 1980) and described in more detail recently (Gavin 2001). Previous studies in the Rivière Boniface area have described the structure and old age of spruce woodlands and krummholz (Payette et al. 1985, 1989a,b; Payette & Morneau 1993), composed of both living and dead spruce stems and branches several hundred years old. Slowly decomposing dead wood remains on the ground for decades and centuries, sometimes with extreme residence times of ca 600 years (Payette & Morneau 1993) and, as a result, may yield potentially anomalous, much older fire dates when burned. Thus, caution must be exercised when interpreting charcoal dates from forest-tundra environments where old-growth trees and stands are common. For a given fire, charcoal dates in table 1 (appendix A) are often much older than the age of the associated burn because they originated probably from trees that died several years before the fire event. This interpretation is based on the fact that all extant spruce stands at the time fire occurred were old-growth woodlands and krummholz composed of old living and dead spruce stems. Most fires that occurred during the last 900 years burned old-growth spruce stands, as shown in a particular case by Arseneault & Payette (1992).
Table 1.
Fire occurrence and burned areas per 100 year intervals in the Rivière Boniface area based on tree-ring data and radiocarbon-dated charcoal and wood. (Each 14C date from 1100 cal. yr BP onwards (to 200–100 BC) corresponds to a dated fire event. 14C dates in box from 1100 cal. yr BP to present correspond to a dated fire event. All the other 14C dates correspond to previous fires.)
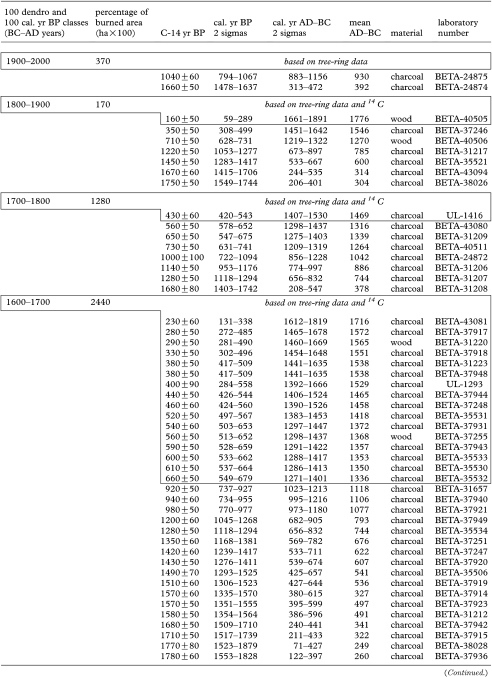
|
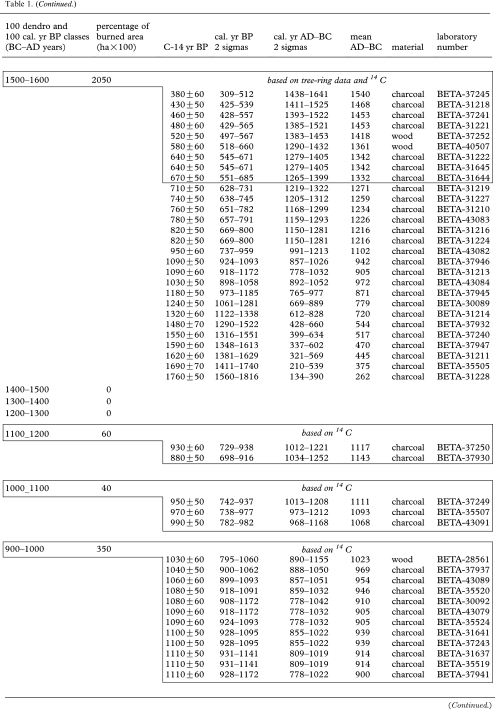
|
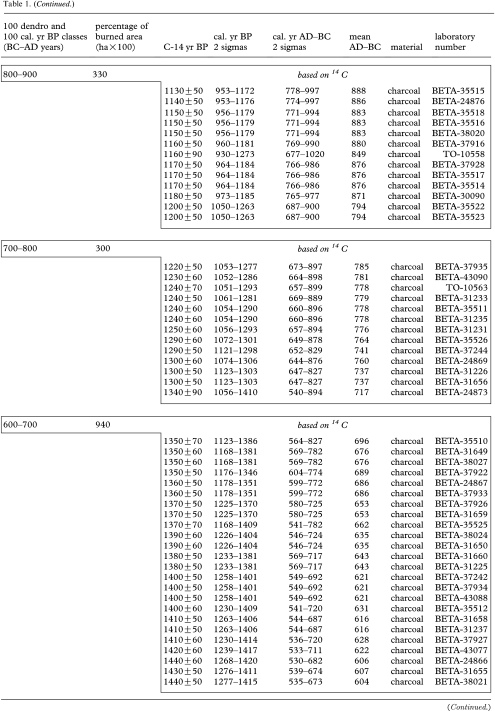
|
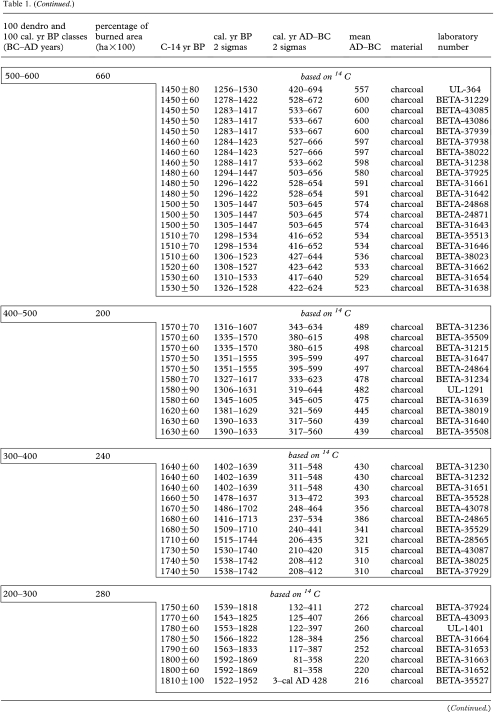
|
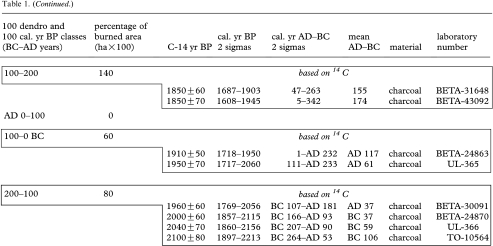
|
Charcoal at the soil surface of a forest stand is distributed more or less discontinuously according to the distribution of charcoal-prone materials when fire occurred. When successive fires override the same site, then the distribution of charcoal materials reflects the distribution of trees burned at different times (figure 8). Charcoal produced by different post-fire regenerations of trees is thus distributed more widely (and sometimes piled or mixed) at the soil surface. Because charcoal decomposes less rapidly than other organic materials, it will remain for a much longer period at the soil surface, in most cases for thousands of years (Payette & Morneau 1993). Given the fact that charcoal sampling at a site is a routine process of trial and error, every piece of charcoal (i.e. a given black spruce stem, branch or cone fragment) sampled at one point corresponds probably to the location of a tree burned during the last fire or previous fires. However, charcoal produced during a given fire will be probably consumed partly or completely by the next fire, a natural process that reduces greatly the probability of survival of ancient charcoal in sites frequently disturbed by fires. Charcoal-to-ash transformation due to complete combustion of wood material during successive fires probably reduces to a large extent the residence time of charcoal at the soil surface. Again, caution should be used here when sampling for charcoal pieces and interpreting charcoal dates. Thus, it is not readily apparent that a given radiocarbon date of charcoal corresponds to the approximate date of the last fire occurring at the site. Complementary information, like the distribution of clone size, and further enquiry on soil and plant characteristics of the fire site is often necessary to determine the date of the last fire event.
Figure 8.

(a–d) Spatiotemporal distribution of charcoal associated with successive forest fires. Charcoal on the ground reflects the distribution of different post-fire regenerations of trees and charcoal survival associated with successive fires.
A comparative analysis of the distribution and abundance of black spruce trees and stratigraphy of organic soil horizons at the landscape scale was used to identify the chronology of fire conflagrations and correlative post-fire tree regeneration success. The following patterns of soil–plant–fire interactions distributed along soil toposequences were deduced from our survey of fire evidence and vegetation development and correspond to the distribution of dominant forest and tundra communities at the landscape scale (figure 9). (i) Fire-free sites are those sites without charcoal macrofossils beneath, in, and on the current organic horizon (corresponding to 10% of the surface of the study area) and with stands composed of a dense cover (generally >40%) of black spruce trees and shrubs—i.e. sites free of fire disturbance during a long period, but of unknown duration, of the Holocene. These sites are surrounded by wetlands colonized by dense spruce vegetation, possibly untouched by fire during a long period of time of the Holocene. (ii) Sites burned only once are those sites with patches of charcoal-free organic horizons in a matrix of organic horizons burned completely and replaced by a charcoal layer overlaid by the modern organic horizon (<5% of the sites of the study area). This corresponds to stands composed of a fragmented cover (<10%) of black spruce trees and shrubs—i.e. sites burned only once during a long period, but of unknown duration, of the Holocene. Given the fact that these stands were formerly growing on soils devoid of charcoal, one can infer that they were deforested in a one-shot fire event. These sites are surrounded by wetlands colonized by dense spruce vegetation, possibly untouched by fire during a long period of the Holocene. (iii) Sites with charcoal macrofossils beneath the current organic horizon (corresponding to more than 20% of the sites of the study area) are stands composed of a relatively dense cover (generally between 10 and 40%) of black spruce trees and shrubs—i.e. sites burned probably more than twice since initial forest establishment with successful tree regeneration because they are currently forming woodlands and krummholz. These sites are surrounded by wetlands colonized by dense spruce vegetation touched by fire. (iv) Sites with charcoal macrofossils beneath, in, and above the current organic horizon (corresponding to the large majority of the remaining sites of the study area) are treeless stands composed of a much scattered cover (generally <1%) of black spruce shrubs—i.e. sites burned probably more than twice since initial forest establishment, without successful tree regeneration and currently forming lichen-shrub tundra. These sites are expanding and are often surrounded by treeless wetlands owing to progressive penetration of fire in nearby wetlands, facilitated by previous fire conflagrations. Given these a posteriori pattern-and-process combinations, it has been possible to reconstruct the chronology of fire occurrences and correlative tree regeneration of all the study sites over the last 2000 years.
Figure 9.

Vegetation patterns along soil toposequences illustrating forest-fire interactions through time. (a) Dense lichen woodlands in a fire-free landscape and corresponding to 10% of the study sites. (b) Less dense lichen woodlands in a landscape with fire occurring only once during the Holocene. (c) Lichen woodlands in a landscape with successive fires. (d) Lichen-heath tundra resulting from deforestation of stands associated with successive fires. Charcoal fragments are indicated in black at the soil surface.
(c) Fire statistics
The number of sites burned per 100 year interval varied greatly over the last 2000 years (appendix A). Cumulated time-since-fire dates calculated from the 500 cross-dated pre-fire stems correspond to major depressions in the curve of the date of mortality of burned stems among the 130 stands burned over the last 450 years (figures 6 and 7e). Five major fire events were recorded in the Rivière Boniface region from the 1500s to present: 1550–1590s (particularly in 1568, see also Arseneault & Payette 1997), 1640–1650s, 1750 (see also Arseneault & Payette 1992), 1850s and 1900–1920s. From the eighteenth century to present, 49 individual burned sites were recorded corresponding to six scar-dated and tree-ring cross-dated fires, i.e. 1750, ca 1850, ca 1900, 1906, ca 1924 and 1943. Also, an exceptionally small fire occurred ca 1650, as an individual tree was struck by lighting. This small number of individual fire years indicates a very low fire frequency of 0.04, 0.03 and 0.02 fires yr−1 over the last 100 (twentieth century), 200 (nineteenth and twentieth centuries) and 300 years (eighteenth, nineteenth and twentieth centuries), respectively. The number of individual sites burned over the last 100 (twentieth century), 200 (nineteenth and twentieth centuries) and 300 years (eighteenth, nineteenth and twentieth centuries) was 7, 10 and 32, respectively. On the other hand, the number of sites burned during the 1500s and the 1600s was 38 and 41, respectively, and corresponded to relatively large fire areas. It is probable that these fire tracks obliterated former fire areas between the 1200s and the 1500s, thus explaining the lack of fire records for this 300 year period (appendix A: table 1).
The fire rotation period calculated per 100 year interval also varied greatly, ca 3000, 4000 and 1800 years over the last 100, 200 and 300 years, respectively. These figures indicate that vegetation tends to develop old-growth stands able to survive for centuries, if not millennia. The duration of the fire rotation period calculated for the last 100, 200 and 300 years appears realistic given the fact that 10% of the surface of the studied sites are still occupied by spruce stands more than 2000 years old.
(d) Chronology of fires and tree regeneration
Plotting all the fire dates calculated for 90% of the surface of the study area gives a portrait of fire occurrence per 100 year interval over the last 2000 years. The time-since-fire distribution (Johnson & Gutsell 1994) shows mixed fire frequencies over the last two millennia (figure 10), corresponding to five fire periods. Period I corresponds to low fire activity over the last 250 years (from AD 1750 to present). Comparatively higher fire activity was registered during period II, 250–450 years ago (from AD 1550 to AD 1750), and period V, 1200–2000 years ago (from 1200 to 2000 cal. yr BP). Fire activity similar to that of period I occurred during period IV, 900–1200 years ago (from 900 to 1200 cal. yr BP). Period III, 450–900 years ago (between 900 cal. yr BP and AD 1550), was probably a quiescent fire period. However, it is highly probable that fire occurred during this period, but the evidence was obliterated by the relatively large fires of period II.
Figure 10.

Cumulative distribution (in log) of burnt areas in relation to time-since-fire subdivided into five periods of contrasted fire activity.
Forest survival or distribution of time-since-fire spruce regeneration, based on the current spruce tree and shrub cover of each fire-dated site, is a direct response to climate forcing during the Late Holocene. Forest survival (figure 11) indicates a marked pattern of varying spruce abundance with a sharp cut-off at AD 1550–1600 (ca 400–500 dendro years). Sustained tree regeneration (>10% cover) was recorded only in sites with charcoal dated to more than 900 cal. yr BP. Without exception, all post-fire woodlands and krummholz are old-growth stands regenerated between 900 and 2200 cal. yr BP. At each successive fire over the last 2200 cal. yr BP, however, spruce cover decreased steadily from an average of 35% at 2200 cal. yr BP to 20% at 900 cal. yr BP and 0.5% during the last 450 dendro years (R2=0.85, p<0.001). Furthermore, all fire-free woodlands and krummholz have a dense spruce cover often greater than 40%, which suggests that each fire event at the tree line has a negative effect on spruce regeneration. More than 63% of the area burned over the last 2200 cal. yr BP burned again during the last 450 years. In particular, 58% of the surface of all the fire-prone sites burned in less than 200 years from AD 1550 to AD 1750. All the fires were followed by a sharp reduction of tree regeneration and consecutive deforestation. Owing to fire overlapping, it is probable that massive spruce deforestation started earlier than during period II, i.e. after 900 cal. yr BP. The inception of lichen-tundra vegetation due to post-fire deforestation started slowly at ca 1800 cal. yr BP. All lichen-tundra sites studied were formerly forested according to abundance of spruce charcoal at the soil surface. Only 15% of the study area was covered by lichen-tundra vegetation 500 years ago (or between 500 and 900 years ago, given the probable incidence of overlapping fires). Overall, the drastic reduction of the forest cover was followed by a lowering of approximately 30 m of the regional forest line—the difference between maximum position in altitude of krummholz and woodland—and approximately 50 m of the regional tree line (i.e. the difference between maximum altitude of spruce macrofossils in tundra sites and position of the current tree line). Finally, no field information was found on the possible influence of Nunamiut hunters on fire activity.
Figure 11.

Forest survival over the last two millennia associated with varying success of post-fire spruce regeneration. Left arrow indicates initial deforestation and inception of the boreal forest-tundra ecotone and right arrow corresponds to start-up of rapid deforestation after AD 1550. Arrow positioned at 900 cal. yr BP corresponds to possible start-up of rapid deforestation, assuming that fire activity from 900 to 600 cal. yr BP was masked by fire overlapping after AD 1550.
The pattern of fire activity depicted over the last two millennia appears to be nested in a suite of favourable dry conditions conducive to fire ignition across Central and Eastern Canada (Filion 1984; Filion et al. 1991; Laird et al. 1996) and the Northern Hemisphere (Ferrett et al. 2005). The distribution and cover of lichen-tundra and lichen-spruce stands in the northern boreal forest region across Eastern Canada are very similar to the measured spatiotemporal patterns in this study. As was the case in our study, the magnitude and timing of post-fire deforestation throughout the northern boreal biome were closely linked to the climatic deterioration that prevailed during the Little Ice Age (Overpeck et al. 1997), i.e. after the Medieval Warmth (ca 900–1200 years ago) and before recent warming commencing at the end of the nineteenth century. The lack of post-fire spruce regeneration from 900 cal. yr BP to present is a direct indication that climatic conditions deteriorated after the Medieval Warmth. According to the distribution of fire dates and tree regeneration, the climate at the tree line was drier and warmer before 900 cal. yr BP. Wetland and lake development in the Rivière Boniface area also showed the same spatiotemporal trend with contrasted tree regeneration and lake-level fluctuations over the last 2000 years (Payette & Delwaide 2004; Asselin & Payette 2006).
4. Conclusion
According to fire and tree regeneration data from our study, the northern part of the forest tundra in Eastern Canada has been heavily deforested over the last millennium. The main direct cause is climate deterioration coinciding with the phasing-out of the Medieval Warmth and incidence of the Little Ice Age. Drier and warmer conditions prevailed before 900 cal. yr BP when forest regenerated in most sites. Given the fact that the latitudinal position of successful post-fire regeneration of lichen-spruce woodlands is situated approximately 1.5° south of the Boniface area (Payette et al. 1989a,b), as a rule of thumb it is probable that a drop of at least 1°C in mean annual temperature occurred after 900 cal. yr BP. At present, the northernmost woodlands and krummholz are less susceptible to burning and being destroyed by fire owing to the still cool and humid climatic conditions prevailing in the northern part of the forest tundra, where fire frequency is very low, and the lack of extensive forest fuels. Recovery of the boreal forest after a long period of deforestation will require sustained warming which, indeed, has currently been taking place since the mid-1990s in Eastern subarctic Canada.
Acknowledgments
Research funding was supported by the Natural Sciences and Engineering Research Council of Canada (NSERC) and the Fonds québécois de la recherche sur la nature et les technologies (FQRNT, Québec). We thank Simon Bélanger, Édith Bonneau, Marie-Claude Bouchard, Mario Charrette, Claude Morneau, Claude Lavoie, Sylvain Lamoureux and several other dedicated students and colleagues for their field and laboratory assistance. We extend our thanks to Prof. Ed Johnson for the kind invitation to participate in the ESA's boreal conference in Montréal and to contribute to this special issue on the boreal forest.
Appendix A.
Footnotes
One contribution of 12 to a Theme Issue ‘The boreal forest and global change’.
References
- Anderson R.S, Hallett D.J, Berg E, Jass R.B, Toney J.L, de Fontaine C.S, De Volder A. Holocene development of boreal forests and fire regimes on the Kenai Lowlands of Alaska. Holocene. 2006;16:791–803. doi: 10.1191/0959683606hl901ft. [DOI] [Google Scholar]
- Arseneault D. Impact of fire behavior on postfire forest development in a homogeneous forest landscape. Can. J. For. Res. 2001;31:1367–1374. doi: 10.1139/cjfr-31-8-1367. [DOI] [Google Scholar]
- Arseneault D, Payette S. A postfire shift from lichen-spruce to lichen-tundra vegetation at tree line. Ecology. 1992;73:1067–1081. doi: 10.2307/1940181. [DOI] [Google Scholar]
- Arseneault D, Payette S. Landscape change following deforestation at the arctic tree line in Québec, Canada. Ecology. 1997;78:693–706. [Google Scholar]
- Asselin H, Payette S. Detecting local-scale fire episodes on pollen slides. Rev. Palaeobot. Palynol. 2005a;137:31–40. doi: 10.1016/j.revpalbo.2005.08.002. [DOI] [Google Scholar]
- Asselin H, Payette S. Late Holocene deforestation of a tree line site: estimation of pre-fire vegetation composition and black spruce cover using soil charcoal. Ecography. 2005b;28:801–805. doi: 10.1111/j.2005.0906-7590.04216.x. [DOI] [Google Scholar]
- Asselin H, Payette S. Origin and long-term dynamics of a subarctic tree line. Ecoscience. 2006;13:135–142. doi: 10.2980/i1195-6860-13-2-135.1. [DOI] [Google Scholar]
- Bégin C, Filion L. Black spruce (Picea mariana) architecture. Can. J. Bot. 1999;77:664–672. doi: 10.1139/cjb-77-5-664. [DOI] [Google Scholar]
- Bergeron Y, Gauthier S, Kafka V, Lefort P, Lesieur D. Natural fire frequency for the Eastern Canadian boreal forest: consequences for sustainable forestry. Can. J. For. Res. 2001;31:384–391. doi: 10.1139/cjfr-31-3-384. [DOI] [Google Scholar]
- Bergeron Y, Gauthier S, Flannigan M, Kafka V. Fire regimes at the transition between mixedwood and coniferous boreal forest in northwestern Québec. Ecology. 2004;85:1916–1932. doi: 10.1890/02-0716. [DOI] [Google Scholar]
- Carcaillet C. Are Holocene wood–charcoal fragments stratified in alpine and subalpine soils? Evidence from the European Alps based on AMS 14C dates. Holocene. 2001;11:231–242. doi: 10.1191/095968301674071040. [DOI] [Google Scholar]
- Carcaillet C, Bergeron Y, Richard P.J.H, Fréchette B, Gauthier S, Prairie Y.T. Change of fire frequency in the Eastern Canadian boreal forests during the Holocene: does vegetation composition or climate trigger the fire regime? J. Ecol. 2001;89:930–946. doi: 10.1111/j.1365-2745.2001.00614.x. [DOI] [Google Scholar]
- Clark J.S. Charcoal-stratigraphic analysis on petrographic thin sections: recent fire history in northwest Minnesota. Quatern. Res. 1988;30:81–91. doi: 10.1016/0033-5894(88)90089-0. [DOI] [Google Scholar]
- Clark J.S, Royall P.D. Local and regional sediment charcoal evidence for fire regimes in presettlement north-eastern North America. J. Ecol. 1996;84:365–382. doi: 10.2307/2261199. [DOI] [Google Scholar]
- Cwynar L.C. Recent history of fire and vegetation from laminated sediment of Greenleaf Lake, Algonquin Park, Ontario. Can. J. Bot. 1978;56:10–21. [Google Scholar]
- Desponts M, Payette S. The Holocene dynamics of jack pine at its northern range limit in Québec. J. Ecol. 1993;81:719–727. doi: 10.2307/2261669. [DOI] [Google Scholar]
- Earle C.J, Brubaker L.B, Anderson P.M. Charcoal in northcentral Alaskan lake sediments: relationships to fire and Late-Quaternary vegetation history. Rev. Palaeobot. Palynol. 1996;92:83–95. doi: 10.1016/0034-6667(95)00095-X. [DOI] [Google Scholar]
- Engelmark O. Forest fires in the Muddus National Park (northern Sweden) during the past 600 years. Can. J. Bot. 1984;62:893–898. [Google Scholar]
- Ferrett D.F, et al. Unexpected changes to the global methane budget over the past 2000 years. Science. 2005;309:1714–1717. doi: 10.1126/science.1115193. [DOI] [PubMed] [Google Scholar]
- Filion L. A relationship between dunes, fire and climate in the Holocene deposits of Québec. Nature. 1984;309:543–546. doi: 10.1038/309543a0. [DOI] [Google Scholar]
- Filion L, Payette S, Gauthier L, Boutin Y. Light rings in subarctic conifers as a dendrochronological tool. Quatern. Res. 1986;26:272–279. doi: 10.1016/0033-5894(86)90111-0. [DOI] [Google Scholar]
- Filion L, Saint-Laurent D, Desponts M, Payette S. The Late Holocene record of aeolian and fire activity in northern Québec, Canada. Holocene. 1991;1:201–208. [Google Scholar]
- Gavin D.G. Estimation of inbuilt age in radiocarbon ages of soil charcoal for fire history studies. Radiocarbon. 2001;43:27–44. [Google Scholar]
- Green D.G. Fire and stability in the postglacial forests of southwest Nova Scotia. J. Biogeogr. 1982;9:29–40. doi: 10.2307/2844728. [DOI] [Google Scholar]
- Grimm E.C. Chronology and dynamics of vegetation change in the prairie-woodland region of southern Minnesota, USA. New Phytol. 1983;93:311–350. doi: 10.1111/j.1469-8137.1983.tb03434.x. [DOI] [Google Scholar]
- Grimm E.C. Fire and other factors controlling the vegetation of the Big Woods region of Minnesota. Ecol. Monogr. 1984;54:291–311. doi: 10.2307/1942499. [DOI] [Google Scholar]
- Heinselman M.L. Fire in the virgin forests of the Boundary Waters Canoe Area, Minnesota. Quatern. Res. 1973;3:329–382. doi: 10.1016/0033-5894(73)90003-3. [DOI] [Google Scholar]
- Holmes R.W. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring B. 1983;43:69–78. [Google Scholar]
- Innes J.L. Lichenometry. Prog. Phys. Geogr. 1985;9:187–254. [Google Scholar]
- Johnson E.A. Cambridge University Press; Cambridge, UK: 1992. Fire and vegetation dynamics: studies from the North American boreal forest. [Google Scholar]
- Johnson E.A, Gutsell S.L. Fire frequency models, methods and interpretations. Adv. Ecol. Res. 1994;25:239–287. [Google Scholar]
- Kuhry P. The role of fire in the development of Sphagnum-dominated peatlands in western boreal Canada. J. Ecol. 1994;82:899–910. doi: 10.2307/2261453. [DOI] [Google Scholar]
- Laberge M.-J, Payette S, Bousquet J. Life span and biomass allocation of stunted black spruce clones in the subarctic environment. J. Ecol. 2000;88:584–593. doi: 10.1046/j.1365-2745.2000.00471.x. [DOI] [Google Scholar]
- Laberge M.-J, Payette S, Pitre N. Development of stunted black spruce (Picea mariana) clones in the subarctic environment: a dendro-architectural analysis. Ecoscience. 2001;8:489–498. [Google Scholar]
- Laird K.R, Fritz S.C, Maasch K.A, Cummning B.F. Greater drought intensity and frequency before AD 1200 in the Northern Great Plains, USA. Nature. 1996;384:552–554. doi: 10.1038/384552a0. [DOI] [Google Scholar]
- LaMarche V.C., Jr Environment in relation to age of bristlecone pines. Ecology. 1969;50:53–59. doi: 10.2307/1934662. [DOI] [Google Scholar]
- Lara A, Villalba R. A 3620-year temperature record from Fitzroya cupressoides tree rings in southern South America. Science. 1993;260:1104–1106. doi: 10.1126/science.260.5111.1104. [DOI] [PubMed] [Google Scholar]
- Lavoie C, Payette S. The long-term stability of the boreal forest limit in subarctic Quebec. Ecology. 1996;77:1226–1233. doi: 10.2307/2265591. [DOI] [Google Scholar]
- Lloyd A.H, Graumlich L.J. A 3,500 year record of changes in the structure and distribution of forests at treeline in the Sierra Nevada, California, U.S.A. Ecology. 1997;78:1199–1210. [Google Scholar]
- MacDonald G.M, Larsen C.P.S, Szeicz J.M, Moser K.A. The reconstruction of boreal forest fire history from lake sediments: a comparison of charcoal, pollen, sedimentological, and geochemical indices. Quatern. Sci. Rev. 1991;10:53–72. doi: 10.1016/0277-3791(91)90030-X. [DOI] [Google Scholar]
- Niklasson M, Granström A. Numbers and sizes of fires: long-term spatially explicit fire history in a Swedish boreal landscape. Ecology. 2000;81:1484–1499. [Google Scholar]
- Ohlson M, Tryterud E. Interpretation of the charcoal record in forest soils: forest fires and their production and deposition of macroscopic charcoal. Holocene. 2000;10:519–525. doi: 10.1191/095968300667442551. [DOI] [Google Scholar]
- Oliver C.D, Larson B.C. Wiley; New York, NY: 1996. Forest stand dynamics. [Google Scholar]
- Overpeck J, et al. Arctic environmental change of the last four centuries. Science. 1997;278:1251–1256. doi: 10.1126/science.278.5341.1251. [DOI] [Google Scholar]
- Patterson W.A, III, Edwards K.J, MacGuire D.J. Microscopic charcoal as a fossil indicator of fire. Quatern. Sci. Rev. 1987;6:3–23. doi: 10.1016/0277-3791(87)90012-6. [DOI] [Google Scholar]
- Payette S, Delwaide A. Dynamics of subarctic wetland forests over the past 1500 years. Ecol. Monogr. 2004;74:373–391. doi: 10.1890/03-4033. [DOI] [Google Scholar]
- Payette S, Gagnon R. Late Holocene deforestation and tree regeneration in the forest-tundra of Quebec. Nature. 1985;313:570–572. doi: 10.1038/313570a0. [DOI] [Google Scholar]
- Payette S, Lajeunesse R. Les combes à neige de la rivière aux Feuilles (Nouveau-Québec): indicateurs paléoclimatiques holocènes. Géogr. Phys. Quatern. 1980;34:209–220. [Google Scholar]
- Payette S, Morneau C. Holocene relict woodlands at the Eastern Canadian treeline. Quatern. Res. 1993;39:84–89. doi: 10.1006/qres.1993.1010. [DOI] [Google Scholar]
- Payette S, Filion L, Gauthier L, Boutin Y. Secular climate change in old-growth tree-line vegetation of northern Québec. Nature. 1985;315:135–138. doi: 10.1038/315135a0. [DOI] [Google Scholar]
- Payette S, Filion L, Delwaide A, Bégin C. Reconstruction of tree-line vegetation response to long-term climate change. Nature. 1989a;341:429–432. doi: 10.1038/341429a0. [DOI] [Google Scholar]
- Payette S, Morneau C, Sirois L, Desponts M. Recent fire history of the northern Québec biomes. Ecology. 1989b;70:656–673. doi: 10.2307/1940217. [DOI] [Google Scholar]
- Payette S, Fortin M.-J, Gamache I. The subarctic forest-tundra: the structure of a biome in a changing climate. BioScience. 2001;51:709–718. doi: 10.1641/0006-3568(2001)051[0709:TSFTTS]2.0.CO;2. [DOI] [Google Scholar]
- Payette S, Boudreau S, Morneau C, Pitre N. Long-term interactions between migratory caribou, wildfires and Nunavik hunters inferred from tree rings. Ambio. 2004;33:482–486. doi: 10.1639/0044-7447(2004)033[0482:LIBMCW]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- Peňalba C, Payette S. Late-Holocene expansion of eastern larch (Larix laricina [Du Roi] K. Koch) in northwestern Québec. Quatern. Res. 1997;48:114–121. doi: 10.1006/qres.1997.1906. [DOI] [Google Scholar]
- Ritchie J.C. Cambridge University Press; Cambridge, UK: 1987. Postglacial Vegetation of Canada. [Google Scholar]
- Rowe J.S. Canada Forestry Service Publication 1300. Information Canada; Ottawa, Canada: 1972. Forest Regions of Canada. [Google Scholar]
- Saint-Laurent D, Filion L. Interprétation paléoécologique des dunes à la limite des arbres, secteur nord-est de la mer d'Hudson, Québec. Géogr. Phys. Quatern. 1992;46:209–220. [Google Scholar]
- Simard, M. 1996 Développement spatio-temporel d'un clone d'épinette noire à la limite des arbres. BSc thesis, Université Laval, Québec.
- Stähli M, Finsinger W, Tinner W, Allgöwer B. Wildfire history and fire ecology of the Swiss National Park (Central Alps): new evidence from charcoal, pollen and plant macrofossils. Holocene. 2006;16:805–817. [Google Scholar]
- Stuiver M, et al. INTCAL 98: radiocarbon age calibration 24 000–0 cal BP. Radiocarbon. 1998;40:1041–1083. [Google Scholar]
- Swain A.M. Environmental changes during the past 2000 years in north-central Wisconsin: analysis of pollen, charcoal, and seeds from varved lake sediments. Quatern. Res. 1978;10:55–68. doi: 10.1016/0033-5894(78)90013-3. [DOI] [Google Scholar]
- Swain A.M. Landscape patterns and forest history in the Boundary Waters Canoe area, Minnesota: a pollen study from Hug Lake. Ecology. 1980;61:747–754. doi: 10.2307/1936744. [DOI] [Google Scholar]
- Swetnam T.W. Fire history and climate change in giant sequoia groves. Science. 1993;262:885–889. doi: 10.1126/science.262.5135.885. [DOI] [PubMed] [Google Scholar]
- Talon B, Payette S, Filion L, Delwaide A. Reconstruction of the long-term fire history of an old-growth deciduous forest in southern Québec, Canada, from charred wood in mineral soils. Quatern. Res. 2005;64:36–43. doi: 10.1016/j.yqres.2005.03.003. [DOI] [Google Scholar]
- Tande G.F. Fire history and vegetation pattern of coniferous forests in Jasper National Park, Alberta. Can. J. Bot. 1979;57:1912–1931. doi: 10.1139/b79-241. [DOI] [Google Scholar]
- Terasmae J, Weeks N.C. Natural fires as an index of paleoclimate. Can. Field Nat. 1979;93:116–125. [Google Scholar]
- Timoney K.P, LaRoi G.H, Zoltai S.C, Robinson A.L. The high subarctic forest-tundra of northwestern Canada: position, width, and vegetation gradients in relation to climate. Arctic. 1992;45:1–9. [Google Scholar]
- Tryterud E. Forest fire history in Norway: from fire-disturbed pine forests to fire-free spruce forests. Ecography. 2003;26:161–170. doi: 10.1034/j.1600-0587.2003.02942.x. [DOI] [Google Scholar]
- Umbanhowar C.E., Jr Interaction of fire, climate and vegetation change at a large landscape scale in the Big Woods of Minnesota, USA. Holocene. 2004;14:661–676. doi: 10.1191/0959683604hl745rp. [DOI] [Google Scholar]
- Van Wagner C.E. Age class distribution and the forest fire cycle. Can. J. For. Res. 1978;8:220–227. [Google Scholar]
- Vézinet M. Ministère des Affaires culturelles, Gouvernement du Québec; Québec, Canada: 1980. Les Nunamiut, Inuit au Coeur des Terres. [Google Scholar]
- Wein R.W, Burzinski M.P, Sreenivasa B.A, Tolonen K. Bog profile evidence of fire and vegetation dynamics since 3000 years BP in the Acadian forest. Can. J. Bot. 1987;65:1180–1186. [Google Scholar]
- Whitlock C, Bartlein P.J. Holocene fire activity as a record of past environmental change. In: Gillespie A, Porter S.C, Atwater B.F, editors. The Quaternary period in the United States. vol. 1. Elsevier; Amsterdam, The Netherlands: 2004. pp. 479–489. [Google Scholar]
- Whitlock C, Millspaugh S.H. Testing the assumptions of fire-history studies: an examination of modern charcoal accumulation in Yellowstone National Park, USA. Holocene. 1996;6:7–15. [Google Scholar]