

1. Introduction
Madagascar's imperilled biota are now experiencing the effects of a new threat—climate change (Raxworthy et al. 2008). With more than 90% endemism among plants, mammals, reptiles and amphibians, the stakes are high. The pristine landscapes that allowed this exceptional biodiversity to survive past climate changes are largely gone. Deforestation has claimed approximately 90% of the island's natural forest (Ingram & Dawson 2005; Harper et al. 2007) and what remains is highly fragmented, providing a poor template for large-scale species range shifts. The impacts of current and future climate change may therefore be much different than past impacts, with profound implications for biodiversity.
We review evidence of past response to climate change, models of future change and projected biological response, developing insights to formulate adaptation actions for reducing extinction in Madagascar's biota. We then explore the cost of implementing actions and examine new income opportunities developing through efforts to mitigate climate change.
2. Past climate change and biological response
The diversity of Madagascar's climates, their variability over time and the contrast between the eastern and western slopes have been invoked to explain patterns of endemism (Perrier de la Bâthie 1921; Dewar & Richard 2007). Climate change during past glaciations suggests mechanisms shaping distributions of some modern taxa (Wilmé et al. 2006). Major climate changes accompanied global glaciations, with drier/cooler and wetter/hotter periods and rapid transitions (Wells 2003). The fauna has been shaped by climatic unpredictability (Dewar & Richard 2007) and riverine forests have served as pathways or refugia for many forest taxa (Ganzhorn & Sorg 1996).
Upland refugia have been proposed in areas retaining relict vertebrate and plant populations (Rakotondrainibe 2000; Andreone et al. 2005). Rivers with high-elevation headwaters have served as palaeo-refuges, maintaining riparian forest where species retreated during the driest periods. As conditions became wetter, riparian habitats extended into tributaries with lower headwaters, opening dispersal corridors. Retreat–dispersion watersheds (figure 1, A–J) with higher headwaters are separated by others originating at lower elevations (figure 1, watersheds 1–12) that were isolated during dry periods; contraction and loss of riparian habitats rendered the latter more isolated, leading to high endemism.
Figure 1.

Watershed headwaters in Madagascar. High-elevation headwaters (A–J) are associated with range migrations in response to past climate change; low-elevation headwaters (1–12) are associated with high endemism due to isolation resulting from limited range migrations.
The distributions of many animal groups can be explained by the retreat–dispersion hypothesis (Wilmé et al. 2006); preliminary analyses indicate that the flora also conforms to this scenario. Conserving riverine corridors and intervening isolated forest blocks are thus critical to maintaining resilience in the face of future climate change.
3. Projected future climate change and response
Pronounced climate change is expected in this century. Projections indicate mean temperature increases of 1.1–2.6°C throughout the island (figure 2a) with greatest warming in the south and least along the coast and in the north (Tadross et al. 2008). These spatial characteristics have important biological implications because the south is already the driest region and the highly fragmented eastern forest is vulnerable to drying.
Figure 2.

(a) Maximum and minimum temperature increase scenarios for Madagascar. (i) Minimum and (ii) maximum increase derived from a suite of climate change projections. (b) Median change in total monthly rainfall (mm per month) for (i) January, (ii) April, (iii) July and (iv) September from six downscaled global models (for the period centred on 2055). Regions where three models give positive and three negative changes are left blank, as are those with increases less than 10 mm per month, which will be offset by increases in potential evapotranspiration.
Projections for median rainfall, from statistical downscaling of six general circulation models (Hewitson & Crane 2006), are presented in figure 2b and discussed by Tadross et al. (2008). Rainfall should increase in summer (January–April), and winter (July–September) will be drier along the southeast coast by 2050 but wetter elsewhere.
Dynamic global vegetation model (BIOME and MAPPS) projections indicate Madagascar will lose 11–27% of its current habitat due to climate change if range migrations are possible (perfect dispersal) and 17–50% if not (no dispersal; Malcolm et al. 2006). They also indicate biome migration rates of 144–532 m yr−1, significantly higher than average post-glacial rates (100–200 m yr−1).
Models for 74 endemic plant species indicate significant changes in suitable climate space by 2080 in nearly all cases (Schatz et al. 2008, table 1). Substantial agreement was found among six projections made using three climate models and IPCC (A2a and B2a) scenarios (see electronic supplementary material). Eighteen per cent of species were projected to expand in range and 45% to contract, even under the optimistic full dispersal assumption. Thirty seven per cent had mixed projections. Expansion/contraction is not the only response; as elsewhere, projections include range shifts along altitudinal and moisture gradients. Full assessment of threat from climate change must consider potential range size change combined with ability to migrate sufficiently. In southwestern Madagascar, where projected temperature changes are greatest, many species are predicted to undergo range expansion, but future distributions overlap very little with current ones, suggesting migration will be essential for species to persist, a process requiring maintenance of maximum connectivity between areas of suitable habitat. Extinction risk appears to be greater among habitat specialists, and narrow endemics are consistently predicted to fare badly under future scenarios. Low-elevation humid littoral forest endemics show dramatic range reduction—six out of the seven species modelled suffer 98% or greater loss under at least one model/scenario—a situation compounded by expected sea-level rise that will threaten all 380 littoral forest endemics (Consiglio et al. 2006).
Table 1.
Summary of projected range changes under full dispersal assumption for 74 endemic plant species using climate change scenarios for 2080 (Schatz et al. 2008).
| vegetation/bioclimate/geography | expansion (≥4 models) | contraction (≥4 models) | mixed | ‘worst case scenario’a |
|---|---|---|---|---|
| eastern humid littoral forest | 1 | 6 | 0 | 6 |
| low- to mid-elevation humid to sub-humid forest | 3 | 15 | 4 | 6 |
| mid-elevation sub-humid forest | 0 | 6 | 0 | 1 |
| central high plateau sub-humid to montane tapia woodland | 0 | 3 | 0 | 3 |
| western dry forest | 2 | 1 | 0 | 0 |
| northern dry to sub-humid forest | 3 | 6 | 0 | 1 |
| southwestern sub-arid forest/thicket | 11 | 1 | 2 | 0 |
| western dry forest and southwestern sub-arid forest/thicket | 4 | 0 | 1 | 0 |
| southeastern sub-humid to sub-arid transitional forest | 0 | 1 | 0 | 1 |
| widespread/broad tolerance | 1 | 3 | 0 | 0 |
| totals | 25 | 42 | 7 | 18 |
At least one model projects ≥95% contraction.
Empirical evidence for climate change impacts already exists. Winters were drier in Ranomafana National Park from 1986–2005 relative to 1960–1985 and fruit production and lemur survivorship decreased (Wright 2007). Repeat observations in 1993 and 2003 of amphibians and reptiles suggest that montane endemics, restricted to elevations within 600 m of the highest summits, are vulnerable to loss due to warming (Raxworthy et al. 2008, in press).
4. Adaptation actions and costs
Madagascar's terrestrial biodiversity is concentrated in forests (including several woody vegetation types), making headwater dispersal and reduced fragmentation and degradation of forests obvious focal points for adaptation actions to reduce climate change impacts. Accordingly, the following three overarching sets of action may be proposed:
restoration and protection of riverine corridor forests important for migration,
maintenance and restoration of connectivity among fragmented forests, especially in regions with high genetic divergence between populations across major riverine corridors, and
management of all remaining natural forest to maximize the potential for species migration in response to climate change.
Riverine corridors have been important as migration pathways in the past and probably will be in the future. Although the current status of these forests is poorly documented, many will probably require restoration to facilitate biological responses to climate change.
Areas between riverine corridors, where isolation has led to local endemism, are particularly vulnerable to species loss during climate change due to their current levels of fragmentation. Native forest restoration in these areas can help restore connectivity and reduce extinction risk due to climate change. Since most of the Madagascar's forests are heavily fragmented, cost-effective restoration must target areas of the highest biological importance.
Because Madagascar has already lost most of its natural forest, what remains will be critical for species range shifts in response to climate change. Madagascar is tripling its protected area network, and approximately 30% of natural forest outside the network is needed to provide forest products and income. Resolving conflicts between maintaining natural forest and meeting human needs is best accomplished through natural forest use substitution, including growing plantations on degraded land to provide alternative sources of wood.
Implementing an effective programme of adaptation actions in Madagascar will be costly. Growing plantations equivalent to one-quarter of the natural forest outside protection would cost approximately US$ 0.4 billion (at an estimated $360 ha−1). These plantations would provide wood and forest product substitutes for communities, relieving pressure on natural forests. Restoring an equivalent area of Madagascar's fragmented forest lands to maintain connectivity would cost approximately US$ 0.8 billion (at an estimated $850 ha−1). Costs of protection and restoration of riverine corridors are poorly known, but would add to this cost, as would the incremental costs from climate change impacts on human communities to avoid conflicts over land use.
To place these costs in context, estimates of climate change-related income (da Fonseca et al. 2007) for Madagascar from post-Kyoto adoption of reduction in emissions from deforestation and degradation (REDD) protocols are in the range of US$ 72–144 million annually. A recent estimate of current conservation investment in protected areas projected an annual investment of US$ 9 million (FTHM Finance 2003). While there may be some cross-subsidy of adaptation from these income streams, adaptation costs are clearly large relative to current conservation funding.
5. Discussion
Recommendations to ensure conservation of Madagascar's biodiversity as climate changes must be based on understanding past and monitoring current responses, projecting future change scenarios, modelling biological response and assessing imperatives of the human–biodiversity interface. Biological considerations suggest that conservation of riverine forest corridors, and management (including restoration) of native forest for connectivity to maximize species response capacity in the face of climate change, are essential elements of an adaptation strategy. While woodlots and other managed production systems cannot replace natural forest, they can simultaneously satisfy human needs and promote biodiversity adaptation.
The potential effectiveness of the actions proposed here depends on two biological assumptions: mechanisms important in past biodiversity response will operate in the future, and species dispersal/colonization capabilities are sufficient to track human-induced climate change. Although climates may become warmer by 2050 than in any past interglacial, response characteristics are expected to persist; upslope range shifts will be essential for survival of lowland species and will depend on maintaining connectivity of forest habitats. Range shift ability of species will remain unchanged and will increasingly constrain response capacity if climate change continues unabated late into this century. This underscores the urgency of international action to combat climate change.
Other assumptions involve socio-political factors, especially the continued commitment of Madagascar's government and the international community to conserving the island's biological heritage and the essential ecosystem goods and services it provides. Major investment, much greater than the sum of current conservation investments and potential income from REDD and payments from ecosystem services (e.g. hydrological services), will be required to sustain current initiatives and implement the adaptation agenda needed to ensure that Madagascar's globally important biodiversity can withstand the impacts of climate change.
The likely impacts of changing climate in Madagascar dictate a second set of responses, not developed here. Human adaptation to climate change will also take place (migration, altered agricultural practices), compounding human hardship and creating additional threats to biodiversity that must in turn be factored into conservation strategies. Where management to maintain biological response capacity conflicts with community use of resources, alternatives will have to be found. Madagascar's widespread poverty makes it imperative to integrate human well-being into conservation planning, and efforts to deal with climate change are no exception.
Footnotes
One contribution of 12 to a Special Feature on ‘Global change and biodiversity: future challenges’.
Supplementary Material
(i) Past climate change and biological response. (ii) Species distribution modeling for projecting biological response. (iii) Calculating restoration and plantation costs. (iv) Maximum REDD income. Appendix 1
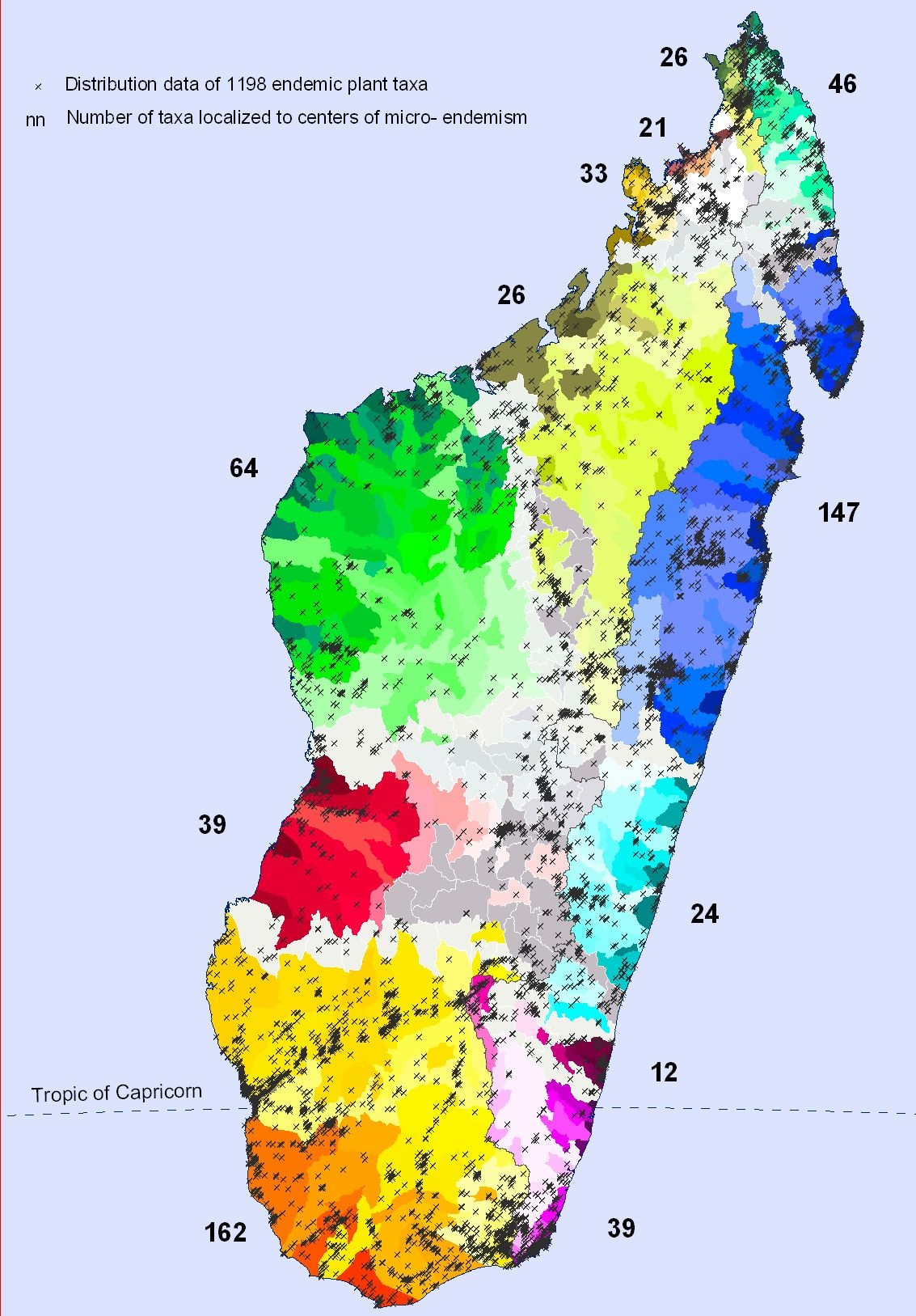
Distribution of 1198 plant taxa endemic to Madagascar from a sample used by APAPC project (Raharimampionona et al. 2005) including 15 133 georeferenced records. Numbers are numbers of taxa localized to the 12 centers of micro-endemism defined by Wilmé et al. (2006) and Wilmé & Callmander (2006)
{kind=link}
References
- Andreone F, Cadle J.E, Cox N, Glaw F, Nussbaum R.A, Raxworthy C.J, Stuart S.N, Vallan D, Vences M. Species review of amphibian extinction risks in Madagascar: conclusions from the global amphibian assessment. Conserv. Biol. 2005;19:1790–1802. doi:10.1111/j.1523-1739.2005.00249.x [Google Scholar]
- Consiglio T, Schatz G.E, McPherson G, Lowry P.P, II, Rabenantoandro J, Rogers Z.S, Rabevohitra R, Rabehevitra D. Deforestation and plant diversity of Madagascar's littoral forests. Conserv. Biol. 2006;20:1523–1739. doi: 10.1111/j.1523-1739.2006.00562.x. doi:10.1111/j.1523-1739.2006.00562.x [DOI] [PubMed] [Google Scholar]
- da Fonseca G.A.B, Rodriguez C.M, Midgley G, Busch J, Hannah L, Mittermeier R.A. No forest left behind. PLoS Biol. 2007;5:1645–1646. doi: 10.1371/journal.pbio.0050216. doi:10.1371/journal.pbio.0050216 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dewar R.E, Richard A.F. Evolution in the hypervariable environment of Madagascar. Proc. Natl Acad. Sci. USA. 2007;104:13 723–13 727. doi: 10.1073/pnas.0704346104. doi:10.1073/pnas.0704346104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- FTHM Finance 2003 Modélisation et projections financières d'ANGAP. A report of the Association Nationale pour la Gestion des Aire Protegees, Madagascar.
- Ganzhorn J.U, Sorg J.P. Ecology and economy of a tropical dry forest in Madagascar. Primate Rep., Deutsches Primatenzentrum, Göttingen (Special issue) 1996;46:1–382. [Google Scholar]
- Harper G, Steininger M, Tucker C, Juhn D, Hawkins F. Fifty years of deforestation and forest fragmentation in Madagascar. Environ. Conserv. 2007;34:325–333. [Google Scholar]
- Hewitson B.C, Crane R.G. Consensus between GCM climate change projections with empirical downscaling: precipitation downscaling over South Africa. Int. J. Climatol. 2006;26:1315–1337. doi:10.1002/joc.1314 [Google Scholar]
- Ingram J.C, Dawson T.P. Climate change impacts and vegetation response on the island of Madagascar. Phil. Trans. R. Soc. A. 2005;363:55–59. doi: 10.1098/rsta.2004.1476. doi:10.1098/rsta.2004.1476 [DOI] [PubMed] [Google Scholar]
- Malcolm J.R, Liu C, Neilson R.P, Hansen L.A, Hannah L. Global warming and extinctions of endemic species from biodiversity hotspots. Conserv. Biol. 2006;20:538–548. doi: 10.1111/j.1523-1739.2006.00364.x. doi:10.1111/j.1523-1739.2006.00364.x [DOI] [PubMed] [Google Scholar]
- Perrier de la Bâthie H. La végétation malgache. Ann. Mus. Col. Marseille, sér. 3. 1921;9:1–273. [Google Scholar]
- Rakotondrainibe, F. 2000 La flore ptéridologique d'une forêt relictuelle sur les hauts Plateaux de Madagascar (la Réserve Spéciale d'Ambohitantely). In Monographie de la forêt d'Ambohitantely (eds J. Ratsirarson & S. M. Goodman). Recherches pour le Développement, Sér. Sci. Biol.16, 37–51.
- Raxworthy, C. J. In press. Global warming and extinction risks for amphibians in Madagascar: a preliminary assessment of potential upslope displacement. Conservation Strategy for the Amphibians of Madagascar. Monogr. Museo Regionale Sci. Nat. Torino41
- Raxworthy C.J, Pearson R.G, Rabibisoa N, Rakotondrazafy A.M, Ramanamanjato J.-B, Raselimanana A.P, Wu S, Nussbaum R.A, Stone D.A. Extinction vulnerability of tropical montane endemism from warming and upslope displacement: a preliminary appraisal for the highest massif in Madagascar. Global Change Biol. 2008;14:1–18. doi:10.1111/j.1365-2486.2008.1596.x [Google Scholar]
- Schatz, G., Cameron, A. & Raminosoa, T. 2008 Modeling of endemic plant species of Madagascar under climate change. Assessing the Impact of Climate Change on Madagascar's Livelihoods and Biodiversity. Conference. Antananarivo, Madagascar, 28 January 2008
- Tadross, M., Randriamarolaza, L., Rabefitia, Z. & Zheng, K. Y. 2008 Climate change in Madagascar; recent past and future, pp. 18. Washington, DC: World Bank.
- Wells, N. A. 2003 Some hypotheses on the Mesozoic and Cenozoic paleoenvironmental history of Madagascar. In The Natural History of Madagascar (eds S. Goodman & J. Banstead), pp. 16–34. Chicago, IL: University of Chicago Press.
- Wilmé L, Goodman S.M, Ganzhorn J.U. Biogeographic evolution of Madagascar's microendemic biota. Science. 2006;312:1063–1065. doi: 10.1126/science.1122806. doi:10.1126/science.1122806 [DOI] [PubMed] [Google Scholar]
- Wright P. Considering climate change effects in lemur ecology and conservation. In: Gould L, Sauther M, editors. Lemurs, ecology and adaptation. Springer; New York, NY: 2007. pp. 385–401. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(i) Past climate change and biological response. (ii) Species distribution modeling for projecting biological response. (iii) Calculating restoration and plantation costs. (iv) Maximum REDD income. Appendix 1
Distribution of 1198 plant taxa endemic to Madagascar from a sample used by APAPC project (Raharimampionona et al. 2005) including 15 133 georeferenced records. Numbers are numbers of taxa localized to the 12 centers of micro-endemism defined by Wilmé et al. (2006) and Wilmé & Callmander (2006)