

Abstract
Following C2 spinal hemisection (C2HS) in adult rats, ipsilateral phrenic motoneuron (PhMN) recovery occurs through a time-dependent activation of latent, crossed-spinal collaterals (i.e., spontaneous crossed phrenic phenomenon; sCPP) from contralateral bulbospinal axons. Ventilation is maintained during quiet breathing after C2HS, but the ability to increase ventilation during a respiratory stimulation (e.g. hypercapnia) is impaired. We hypothesized that long-term expression of the sCPP would correspond to a progressive normalization in ventilatory patterns during respiratory challenge. Breathing was assessed via plethsymography in unanesthetized animals and phrenic motor output was measured in urethane-anesthetized, paralyzed and vagotomized rats. At 2-week post-C2HS, minute ventilation (VE) was maintained during baseline (room air) conditions as expected but was substantially blunted during hypercapnic challenge (68±3% of VE in uninjured, weight-matched rats). However, by 12 weeks the spinal-lesioned rats achieved a hypercapnic VE response that was 85±7% of control (p = 0.017 vs. 2 wks). These rats also exhibited augmented breaths (AB's) or “sighs” more frequently (p<0.05) than controls; however, total AB volume was significantly less than control at 2- and 12-week post-injury (69±4% and 80±5%, p<0.05, respectively). We also noted that phrenic neurograms demonstrated a consistent delay in onset of the ipsilateral vs. contralateral inspiratory phrenic burst at 2-12-week post-injury. Finally, the ipsilateral phrenic response to respiratory challenge (hypoxia) was greater, though not normalized, at 4-12- vs. 2-week post-injury. We conclude that recovery of ventilation deficits occurs over 2-12-week post-C2HS; however, intrinsic neuroplasticity remains insufficient to concurrently restore a normal level of ipsilateral phrenic output.
Keywords: Plasticity, Crossed phrenic phenomenon, Phrenic, Motoneurons
Introduction
The “crossed phrenic phenomenon” (CPP) is a well-documented example of intrinsic motor recovery following spinal cord injury (SCI) and is evoked after removal of descending brainstem projections to phrenic motoneurons (PhMN) via C2 hemisection (C2HS). Substantial ipsilateral phrenic output can be acutely restored by a variety of experimental manipulations that activate latent, crossed-spinal pathways caudal to C2. These midline projections are comprised of axonal collaterals from intact, contralateral bulbospinal fibers (Goshgarian, 2003; Zimmer et al., 2007). The CPP also is expressed naturally (i.e., spontaneous CPP; sCPP) as ipsilateral phrenic inspiratory bursting which resumes weeks (Fuller et al., 2003) to months post-C2HS (Nantwi et al., 1999). This intrinsic recovery of PhMN activity can be manipulated in onset and magnitude thus indicating that targeted phrenic plasticity is a viable therapeutic goal after cervical SCI (Nantwi and Goshgarian, 2001, 2002, 2005; Golder and Mitchell, 2005).
The relative contribution of the sCPP to long-term recovery of breathing behavior itself, however, is uncertain and to what degree this is coupled to PhMN output has not been fully addressed. While it would initially seem that any respiratory-related electrical activity detected in the ipsilateral phrenic nerve or hemi-diaphragm would reflect improved patterns of breathing, this is not necessarily the case. For example, it is unknown what percentage of the total ipsilateral phrenic motor pool is activated by the sCPP. A functional impact on respiratory-related pressures and volumes during breathing may not be detectable if a relatively small fraction of phrenic motoneurons are recruited. Previously, we reported that the sCPP makes no apparent contribution to breathing by 5-week post-injury (Fuller et al., 2006). On the other hand, Golder et al. (2003) demonstrated that C2HS coupled with ipsilateral phrenicotomy (i.e. preventing the sCPP impulses from reaching the diaphragm) reduced the volume of augmented breaths (i.e. “sighs”) under anesthesia (Golder et al., 2003). Augmented breaths are associated with a large increase in respiratory neural drive that results in activation of a much larger percentage of the phrenic motor pool vs. quiet breathing (Sieck and Fournier, 1989, Golder et al., 2005). Thus, the sCPP may be functionally relevant only under conditions of very high respiratory drive (Golder et al., 2003), and assessment of augmented breaths may provide an opportunity to monitor the physiological importance of the sCPP after C2HS.
Respiratory behavior in unanesthetized animals after C2HS has been characterized only up to 5-week post-injury (Fuller et al., 2006). However, increases in ipsilateral phrenic output (i.e. increased sCPP) may occur later (Nantwi et al., 1999). Accordingly, the goal of this study was to determine whether by 12-week post-C2HS, respiratory volumes during conditions of high respiratory drive would show recovery in parallel with increases in IL PhMN output. Thus, our objective was to extend upon prior reports (Goshgarian et al., 1986; Golder et al., 2001a, b, 2003; Fuller et al., 2006) through a detailed analysis of respiratory behavior using a reduced neurophysiological preparation coupled with behavioral analyses in unanesthetized rats. These studies were intended to provide a foundation for assessing future therapeutic manipulations of the sCPP (e.g. Nantwi and Goshgarian, 2001, 2002, 2005; Golder and Mitchell, 2005; Alilain et al., 2007).
Materials and methods
Experiments were conducted using adult male Sprague-Dawley rats obtained from Harlan Inc. (Indianapolis, IN, USA; colony 211A). Plethysmography data were collected in a total of 35 spinal injured rats at 2- (n = 12), 4- (n = 9), 8- (n = 9) and 12-week post-C2HS (n = 6). All rats were injured at ~3 months of age (mean =89±3 days), and thus post-injury measures (i.e. 2-12 weeks) were necessarily obtained at different ages. Since we observed in preliminary plethysmography experiments that body weight has a substantial impact on ventilation over the age ranges used in this study (see Ventilation in C2HS rats relative to weight-matched control rats), three control groups were selected from rats 3-6 months of age that were weight-matched to the C2HS rats at 2-, 4-8-, and 12-week post-injury (see Table 1). These groups are designated as the “2-, 4-8-, and 12-week controls”. The C2HS rats at 4- and 8-week post-injury had similar body weight and ventilation (Table 1, Fig. 4), and accordingly they were grouped and compared to a single control group.
Table 1.
The age and body weight of uninjured, control rats and C2HS rats at 2-, 4-, 8- and 12-week post-injury
| Group | Age (days) | Body weight (g) |
|---|---|---|
| C2HS rats were injured at 3 months of age, and therefore the post-injury measures were obtained at different ages. Breathing patterns in C2HS rats were compared to rats of similar body weight (Fig. 3). | ||
| 2-week weight control | 85±4 | 300±5 |
| 4-8-week weight control | 106±l | 355±2 |
| 12-week weight control | 108±2 | 450±4 |
| 2-week post-C2HS | 105±4 | 293±8 |
| 4-week post-C2HS | 124±2 | 345±14 |
| 8-week post-C2HS | 149±3 | 354±16 |
| 12-week post-C2HS | 155±4 | 452±13 |
Fig. 4.

Examples of augmented breath behavior recorded with barometric plethysmography in awake rats. These airflow recordings were obtained in a control, uninjured rat (12-week weight control, see Table 1) and a C2HS rat at 12-weeks post-injury during quiet breathing (baseline). The top trace provides a condensed record of approximately 8 min of breathing with augmented breaths indicated by the arrows. The lower traces provide an expanded time scale to demonstrate the appearance of an individual augmented breath (indicated by the arrows).
Neurophysiology data were obtained from spinal-lesioned rats at 2- (n = 9), 4- (n = 9), 8- (n = 10) and 12-week post-C2HS (n = 7). A group of uninjured rats (n = 10, age 4-6 months) was studied primarily to compare differences in the onset of left vs. right phrenic nerve bursting (e.g. Fig. 9). No consistent differences in phrenic output were observed within the control group of uninjured rats. The Institutional Animal Care and Use Committee at the University of Florida approved all experimental procedures.
Fig. 9.

Examples of the onset difference between the ipsilateral (IL) and contralateral (CL) phrenic inspiratory burst in a control (panel A) and C2HS rat (panel B). Data were recorded during baseline conditions (normoxia) in anesthetized rats. In control rats, the onset of IL and CL bursting was essentially simultaneous. In C2HS rats, the onset of the CL burst (dotted line) preceded the IL burst (dashed line). This particular example (panel B) is from an animal at 8-week post-injury. ∫ indicates moving averaged or “integrated” neurogram.
Spinal cord injury
C2HS injuries were performed under isoflurane anesthesia induced in a closed chamber and maintained (2-3%) via nose cone. Following C2 laminectomy and durotomy, microscissors were used to hemisect the spinal cord on the left side, just caudal to the C2 dorsal roots. A gap (~1 mm) at the incision site was then created by gentle aspiration. The overlying muscles were subsequently sutured with polyglycolic acid suture (vicryl) and the skin was closed with 9 mm stainless steel wound clips. Following surgery, rats were given an analgesic (buprenorphine, 0.03 mg/kg, s.q.) and an anti-inflammatory drug (carprofen, 5.0 mg/kg., s.q.). The post-surgical care protocol included buprenorphine (0.03 mg/kg, s.q.) and carprofen (5.0 mg/kg, s.q.) given at 12-hour intervals for 2 days, and lactated ringers solution (18 ml/day, s.q.), given until adequate volitional drinking resumed.
Spinal cord histology
The cervical spinal cord was examined histologically in all animals. At the conclusion of the neurophysiology experiments, urethane-anesthetized rats (see “Neurophysiology (anesthetized rats)”, below) were euthanized by systemic perfusion with heparinized saline followed by 4% paraformaldehyde. The cervical spinal cord was then removed, cryoprotected and sectioned (50 μm) on a cryostat. Tissue sections were serially mounted on glass slides, stained with luxol fast blue and cresyl violet, and examined with light microscopy. Some bulbospinal projections to ipsilateral phrenic motor neurons travel in close proximity to the spinal ventral median fissure (Lipski et al., 1994). Accordingly, hemisections were accepted as “complete” only with the apparent absence of healthy white matter in the ipsilateral spinal cord at C2 (e.g. see Fig. 1 and also Fig. 1 in Doperalski and Fuller, 2006).
Fig. 1.

A representative histological section depicting a C2HS injury. This section was taken from cervical segment C2 at 8-week post-injury. The tissue was stained with luxol fast blue and cresyl violet. The absence of healthy tissue in the ipsilateral spinal cord suggests an anatomically complete C2HS.
Measurement of ventilation via plethysmography in unanesthetized rats
A commercially available barometric plethysmography system (Buxco Inc., Wilmington, NC, USA) was used to quantify ventilation. The plethysmograph was calibrated by injecting known volumes of air into a Plexiglas recording chamber using a 5 ml syringe. The chamber pressure, temperature and humidity, and rectal temperature of the rat were used in the Drorbaugh and Fenn equation (Drorbaugh and Fenn, 1955) to calculate respiratory volumes including minute ventilation (ml/min/100 g) and tidal volume (ml/100 g). Overall breathing frequency (breaths/min), the duration of inspiration and expiration, and peak airflow rates were calculated from the airflow traces. During the experiments, pressurized gas mixtures flowed through the chamber at a rate of 2 L/min to enable control of inspired gases. Baseline recordings lasted 1-1.5 h and were made while the chamber was flushed with 21% O2 (balance N2)(i.e. eucapnic normoxia). Rats were then exposed to a 10 min period of hypercapnic gas (7% CO2, 21% O2, balance N2). Rectal temperature was measured immediately before rats were placed in the chamber, and again immediately after the hypercapnic exposure. If body temperature had dropped by more than 0.1 °C, the Drorbaugh and Fenn equation was used to recalculate the hypercapnic respiratory volumes using the new temperature.
Neurophysiology (anesthetized rats)
Isoflurane anesthesia was induced in a closed chamber and rats were transferred to a nose cone where they breathed 2.5-3.0% isoflurane. The trachea was then cannulated with PE-240 tubing and rats were mechanically ventilated for the remainder of the experiment. A catheter (PE-50) was inserted into the femoral vein, and rats were converted from isoflurane to urethane anesthesia (1.6 g/kg, i.v.; 0.12 g/ml distilled water). The adequacy of anesthesia was monitored during this period by observing limb withdrawal and palpebral reflexes. A femoral arterial catheter (PE-50) was inserted to enable blood pressure measurements (Statham P-10EZ pressure transducer, CP122 AC/DC strain gage amplifier, Grass Instruments, West Warwick, RI, USA) and withdrawal of blood samples. The vagus nerves were sectioned in the mid-cervical region and the rats were paralyzed with pancuronium bromide (2.5 mg/kg, i.v.). Following paralysis, the adequacy of anesthesia was monitored by observing blood pressure and respiratory responses during application of deep pressure to the paws. A 3:1 solution of lactated Ringer's and sodium bicarbonate was continually infused via the venous catheter (1.5 ml/h) to help maintain arterial blood pressure and acid-base regulation. Arterial partial pressures of O2 (PaO2) and CO2 (PaCO2) as well as pH were periodically determined from 0.2 ml arterial blood samples (i-Stat, Heska, Fort Collins, CO, USA). The end-tidal CO2 partial pressure (PETCO2) was measured throughout the protocol using a rapidly responding mainstream CO2 analyzer positioned a few cm from the tracheostomy tube on the expired line of the ventilator circuit (Capnogard neonatal CO2 monitor, Novametrix Medical Systems, Wallingford, CT, USA). Rectal temperature was maintained at 37.5±1 °C using a rectal thermistor connected to a servo-controlled heating pad (model TC-1000, CWE Inc., Ardmore, PA, USA).
A ventral approach was used to isolate both phrenic nerves within the caudal neck region, medial to the brachial plexus. The IL nerve was always the left phrenic nerve, and the CL nerve was always the right phrenic nerve. Electrical activity was recorded using silver wire electrodes with a monopolar configuration, amplified (1000×) and filtered (band pass=300-10,000 Hz, notch=60 Hz) using a differential A/C amplifier (Model 1700, A-M Systems, Carlsborg, WA, USA). The amplified signal was full-wave rectified and moving averaged (time constant 100 ms; model MA-1000; CWE Inc., Ardmore, PA, USA). Data were digitized using a CED Power 1401 data acquisition interface and recorded on a PC using Spike2 software (Cambridge Electronic Design Limited, Cambridge, England).
After an adequate plane of anesthesia was established, the PETCO2 was maintained at 40±2 mmHg for 45-60 min. The end-tidal CO2 apneic threshold for inspiratory activity was then determined by gradually increasing the ventilator pump rate until inspiratory bursting ceased in both phrenic nerves. The ensuing apnea was maintained for 2-3 min, and the ventilator rate was then gradually decreased until inspiratory activity reappeared. The PETCO2 associated with the onset of inspiratory bursting was noted, and the ventilator rate was adjusted to maintain PETCO2 2 mmHg above this value throughout the experiment. The PETCO2 measurements, however, were merely a guide to help maintain isocapnia, and CO2 levels were determined exclusively by arterial blood analyses. An arterial blood sample was drawn a few minutes prior to hypoxia as a baseline measurement. Rats were then exposed to a three-minute bout of hypoxia. At the conclusion of the experiment, rats were euthanized via systemic perfusion (see “Spinal cord histology”, above).
Data analyses
The peak height of the moving averaged phrenic neurograms was analyzed using Spike2 software (CED, Cambridge UK). Phrenic activity was averaged over 30 s periods prior to the first hypoxic episode (baseline), at the onset of and during the final minute of hypoxia. Baseline phrenic output was expressed relative to maximum output and also relative to output in the contralateral nerve (Fuller et al., 2003; Nantwi and Goshgarian, 2002). Changes in neurogram amplitude during hypoxia were expressed relative to the value observed during baseline (i.e. Δ% baseline). The relative onset difference between the ipsilateral and contralateral phrenic inspiratory bursts (e.g. Fig. 8) was calculated from the moving averaged signals using Spike2 software.
Fig. 8.
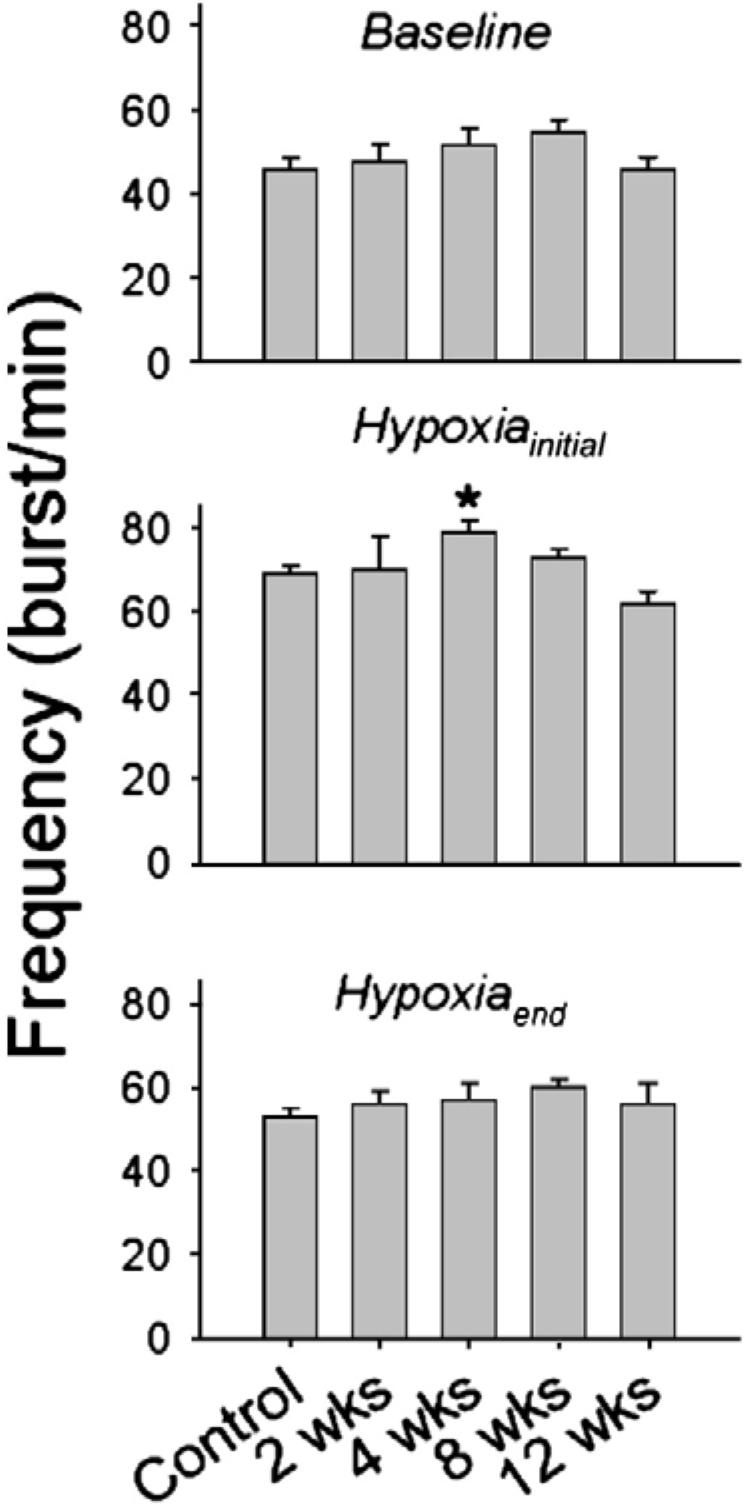
Inspiratory phrenic burst frequency during quiet breathing (baseline) and during the initial 30 s (Hypoxiainitial) and final minute of respiratory stimulation with hypoxia (Hypoxiaend). Data are presented in control (i.e. uninjured) rats, and rats at 2-, 4-, 8- and 12-week post-C2HS. *, significantly different than 12-week rats.
Plethysmography data were analyzed in 1-min bins. For the baseline condition, data represent the average of 10 consecutive 1-min bins just prior to hypercapnia. For hypercapnia, we report the average of the last 5 min of exposure. Respiratory volume data (i.e. tidal volume, VT, and minute ventilation, VE) were expressed per 100 ml of body weight. Spontaneous augmented breaths or “sighs” were analyzed by integrating the plethysmography airflow record using Spike2 software. The integrated airflow trace was used to assess the amplitude and frequency of augmented breaths over the final 30 min of the baseline period. Augmented breaths were identified by a distinct two-phase pattern (Cherniack et al., 1981; Golder et al., 2005)with an initial phase that resembled the preceding breath followed by a second phase characterized by a steeper rate of rise in the integrated airflow trace (see Fig. 5).
Fig. 5.

The impact of C2HS on augmented breath (AB) frequency (breaths per 30 min, top panel) and amplitude (ml/breath/100 g, bottom panel). The appearance of ABs (“sighs”) was analyzed in detail at 2- and 12-week post-C2HS. At 12-week post-injury, ABs occurred at a greater frequency as compared to weight-matched control rats. The relative difference between C2HS and control rats (i.e. C2HS data as “% control”) is listed above each plot along with the p-value. *, C2HS data point is different than corresponding control data point; #, 12-week control is different than the 2-week control.
Statistical comparisons across time points (i.e. control vs. 2-12-week post-injury) were done using analysis of variance (ANOVA) followed by the Student Neuman Keuls (SNK) post hoc test (Sigma-Stat version 1.0; Jandel Scientific, St. Louis, MO, USA). Whenever possible, a two-way ANOVA with repeated measures was used to assess differences across conditions (e.g. baseline vs. hypoxia at different times post-injury). In some cases this was not possible due to differences in data analyses (e.g. % baseline vs. % maximum), and in these cases a one-way ANOVA was used. Differences were considered statistically significant when the p-value was <0.05. All data are presented as the mean±SEM.
Results
A representative histological example demonstrating the completeness of a C2HS lesion is provided in Fig. 1. The body weight and age of control and C2HS rats are presented in Table 1.
Ventilation in C2HS rats relative to weight-matched control rats
Representative examples of airflow recorded during barometric plethysmography experiments are provided in Fig. 2. Preliminary plethysmography experiments in spinal intact rats indicated that breathing patterns could be affected by body weight, even if respiratory volumes are normalized to body mass (Choi et al., 2005; Fuller et al., 2006). In particular, we compared the pattern of breathing in two groups of age-matched, uninjured male Sprague-Dawley rats of differing body weights as follows: group 1, age=103±1 days, weight=354±3 g, n=7; group 2, age=104±1 days, weight=450±2 g, n=9. During quiet breathing, inspiratory frequency was greater in the heavier rats ( p<0.001). During both quiet breathing and hypercapnic challenge, tidal volume was lower in the heavier rats (p<0.001). Accordingly, ventilation data obtained in C2HS rats were compared to rats of similar body weight and within 1-2 months of age (Table 1).
Fig. 2.

Examples of respiratory behavior recorded with barometric plethysmography in awake rats. These airflow recordings were obtained in a C2HS rat at 2-weeks post-injury and an appropriate control rat (see Table 1).
During baseline breathing (i.e. 21% O2, balance N2), minute ventilation was not affected by C2HS (Fig. 3). However, the pattern of breathing was predictably altered with elevated frequency and reduced tidal volume (Fig. 3). Baseline inspiratory tidal volumes were closer to values recorded in weight-matched control rats (hereafter simply stated as control rats or controls) at both 4-8- and 12-week vs. 2-week post-injury ( p<0.05, Fig. 3). For instance, relative to control rats, baseline tidal volume was reduced by 34±2, 25±2 and 23±2% at 2-, 4-8-, and 12-week post-injury. Baseline frequency also was significantly closer to values observed in control rats at 12-week (+23±10% of control) vs. 2-week post-C2HS (+52±10%, p=0.032). At 2-week post-C2HS, rats showed a considerable reduction in hypercapnic tidal volume and minute ventilation (Fig. 3) as previously reported (Fuller et al., 2006). However, relative to controls, the hypercapnic ventilation deficit was actually less in 12- (-15±7%) vs. 2-week post-C2HS rats (-32±3%; 12 vs. 2 weeks: p=0.020).
Fig. 3.

Comparison of ventilation in C2HS rats with weight-matched control rats. Inspiratory frequency (breaths/min), tidal volume (ml/breath/100 g), and minute ventilation (ml/min/100 g) were recorded during quiet breathing (baseline) and hypercapnia. The progressive increase in body weight post-C2HS (Table 1) necessitated the use of multiple control groups for a comparison of ventilation. Thus, each group of C2HS rats (2-week, 4-8-week, and 12-week post-C2HS) is compared to a separate group of weight-matched control rats (see text for details). In each panel, baseline data are presented on the left, and hypercapnic data on the right. The relative difference between C2HS and control rats (i.e. C2HS data as “% control”) is listed above each condition (baseline or hypercapnia) along with the p-value.
Augmented breaths after C2HS
An example of augmented breath behavior in spinal intact and C2HS rats is provided in Fig. 4. Augmented breath frequency averaged 11±1, 14±1, 14±1, and 15±3 breaths per 30 min at 2-, 4-, 8- and 12-week post-C2HS, respectively (p=0.45). Augmented breath frequency at 2-week post-C2HS was similar to that seen in controls (p=0.480), but by 12 weeks augmented breaths occurred at a greater rate than in corresponding controls (p=0.009, Fig. 5). The total volume of the augmented breath (e.g. phase 1+phase 2, see Materials and methods and Fig. 4) was significantly less than observed in appropriate control rats (p<0.05, Fig. 5). Similar to the tidal volume response, the volume of the augmented breath declined over 2-12-week post-injury. Thus, augmented breath volumes were 2.3±0.1, 2.6±0.2, 2.6±0.1 and 1.9±0.1 ml/100 g at 2-, 4-, 8- and 12-week post-C2HS, respectively (p=0.009). Relative to control values, however, augmented breath volumes actually tended to be greater at 12- vs. 2-week post-injury. That is, the relative difference in augmented breath volumes (i.e. vs. controls, Fig. 5) showed a trend towards being reduced in 12-week (-20±6% of control) vs. 2-week post-C2HS rats (-31±4% of control, p=0.15).
Phrenic output
Phrenic neurograms were obtained from the same C2HS animals used for the plethysmography experiments presented above. Arterial blood gases and pressure recorded during the terminal neurophysiology experiments are presented in Table 2. Examples of ipsilateral (IL) and contralateral (CL) phrenic neurograms recorded in control and C2HS rats are provided in Figs. 6 and 9.
Table 2.
Arterial blood gases, pH and mean arterial pressure (MAP) recorded in anesthetized rats during phrenic recording experiments
| Parameter | Group | Baseline | Hypoxia |
|---|---|---|---|
| PaCO2 (mmHg) | Control | 40±l | 39±1 |
| 2 weeks | 37±1 | a33±l | |
| 4 weeks | 36±1 | a32±2 | |
| 8 weeks | 37±1 | b37±l | |
| 12 weeks | 36±2 | 34±2 | |
| PaO2 (mmHg) | Control | 116±8 | 31±1 |
| 2 weeks | a139±9 | 34±1 | |
| 4 weeks | 122±9 | 31±2 | |
| 8 weeks | 122±5 | 33±1 | |
| 12 weeks | 125±6 | 33±1 | |
| pH | Control | 7.32±0.01 | 7.32±0.01 |
| 2 weeks | 7.39±0.01 | 7.38±0.02 | |
| 4 weeks | 7.35±0.01 | 7.35±0.01 | |
| 8 weeks | 7.39±0.02 | 7.39±0.02 | |
| 12 weeks | 7.41 ±0.01 | a7.42±0.02 | |
| MAP (mmHg) | Control | 129±6 | 99±10 |
| 2 weeks | 102±10 | 82±12 | |
| 4 weeks | 101±7 | c49±8 | |
| 8 weeks | 110±4 | 89±6 | |
| 12 weeks | 103±5 | 77±7 |
Different than corresponding value in the control group.
Different than corresponding value in 4-week post-C2HS
Different than all other groups.
Fig. 6.

Examples of phrenic motor output recorded in anesthetized rats. Panels A-C present ipsilateral (IL) and contralateral (CL) phrenic neurograms recorded in control (panel A) and C2HS rats at 2- (panel B) and 8- (panel C) week post-injury during quiet breathing (baseline) and respiratory stimulation with hypoxia. Note that the IL and CL phrenic neurograms are similar in the control rat, but IL burst amplitude is dramatically reduced (vs. CL) at both 2- and 8-week post-injury. All scaling are the same between panels A and C; the calibration bars depict volts. ∫ indicates moving averaged or “integrated” neurogram.
During normoxic baseline conditions, clear IL phrenic bursting (i.e. the sCPP) was observed in 75% of rats at 2-week post-injury, and 100% of rats at 4-12-week post-injury. The absolute (e.g. mV, p<0.001) and relative amplitude (e.g. % peak activity or % contralateral activity, p<0.05) of the IL phrenic burst was reduced in all C2HS rats but IL burst amplitudes did not differ across 2-12 weeks. That is, there was no evidence for progressive recovery of IL bursting during baseline conditions. The amplitudes of the baseline CL phrenic bursts were not different than recordings obtained in the phrenic nerve of control, uninjured rats, and were similar at all post-injury times. Baseline PaCO2, pH and mean arterial pressures (MAPs) were not significantly different across groups; however, PaO2 values were slightly elevated at 2-week post-injury (Table 2).
During hypoxia, all C2HS rats displayed clear IL phrenic bursting, and there was evidence for an increase in the magnitude of the hypoxic response over time post-injury. Specifically, the increase in phrenic bursting evoked by hypoxia (i.e. % baseline) recorded at 4-12-week post-injury was substantially greater than observed in control rats (μ0.05, Fig. 7). However, at 2-week post-injury the hypoxic response of the IL phrenic nerve was not statistically greater than control. Phrenic burst amplitude in the CL nerve during hypoxia was similar across all groups (data not shown). Rats studied at 2- and 4-week post-injury were slightly hypocapnic during hypoxia as compared to the baseline PaCO2 value (Table 2). The 12-week post-injury rats had an elevated arterial pH during hypoxia. All rats became hypotensive during hypoxia (Table 2) as has been reported previously (Fuller et al., 2003). The relative degree of hypoxic hypotension was greatest at 4-week post-injury (p0.05 vs. other groups, Table 2).
Fig. 7.
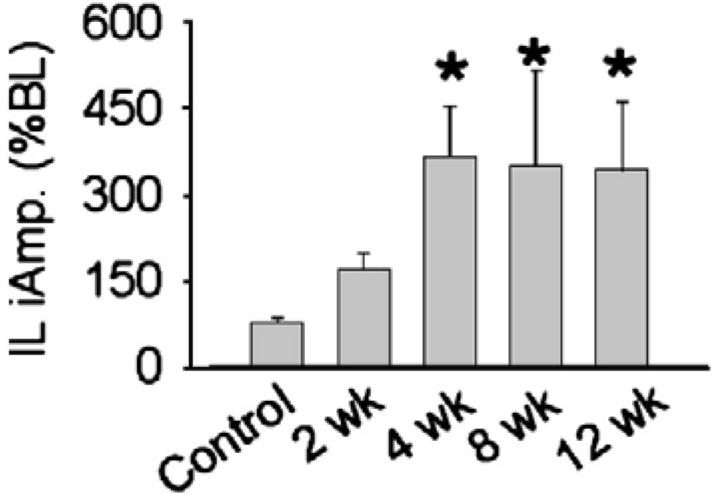
Ipsilateral (IL) phrenic burst amplitude during respiratory stimulation with hypoxia in anesthetized rats. Data are presented from control (i.e. uninjured) rats, and rats at 2-, 4-, 8- and 12-week post-C2HS. Data are expressed relative to baseline bursting (% BL). *, significantly different than control.
Baseline inspiratory phrenic burst frequency was not significantly affected by C2HS (Fig. 8). Hypoxia caused an increase in phrenic burst frequency in all rats (p0.001, Fig. 8), and the only significant difference between experimental groups occurred in the 4-week post-C2HS group. These rats showed a greater phrenic burst frequency during the onset of hypoxia compared to the 12-week rats (p=0.005, Fig. 8). All groups showed a “roll off” in burst frequency during hypoxia, and the values recorded during the final minute of hypoxia were similar across 2-12-week post-injury (Fig. 8).
During analysis of the phrenic neurogram recordings we also noted an interesting change in the pattern of bilateral PhMN bursting activity in C2HS rats. While the onset of IL and CL phrenic inspiratory bursts was essentially simultaneous in control rats during both baseline and hypoxia (Fig. 9), the initiation of IL bursting occurred after the CL bursts in all C2HS rats (p0.001 vs. control; Figs. 9 and 10). Under baseline conditions, the relative onset difference was similar across 2-12-week post-C2HS (Fig. 10). The IL vs. CL phrenic burst onset difference was reduced during hypoxia in C2HS rats, Fig. 10).
Fig. 10.
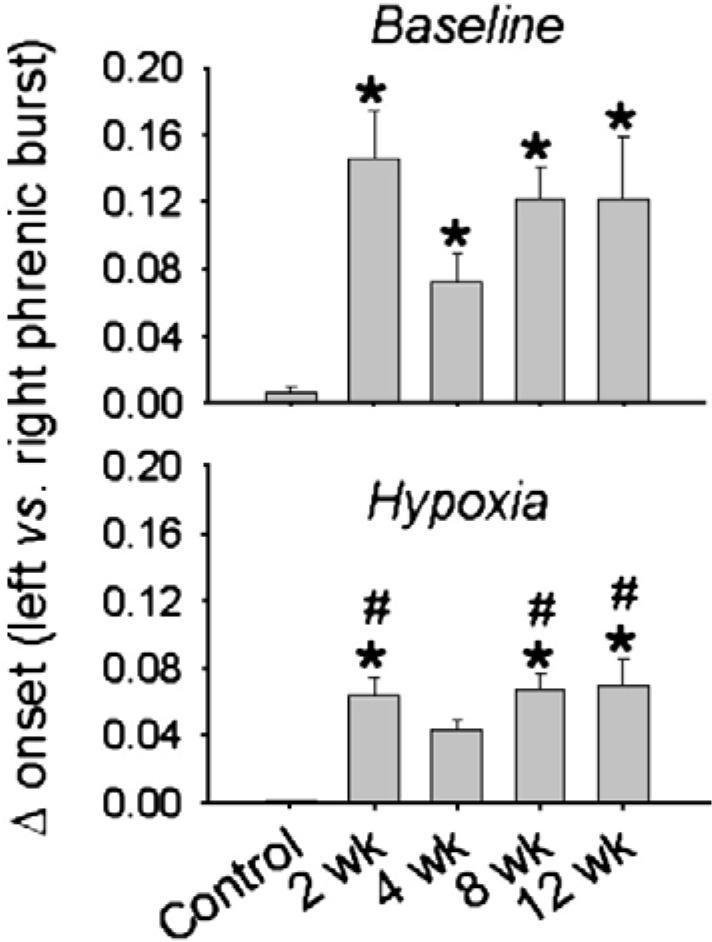
Mean onset difference (s) between the ipsilateral (IL) and contralateral (CL) phrenic inspiratory burst during quiet breathing (baseline) and respiratory stimulation with hypoxia. Data are presented in control (i.e. uninjured) rats, and rats at 2-, 4-, 8- and 12-week post-C2HS. An onset difference was observed in all C2HS rats, and the onset difference was significantly less during respiratory stimulation with hypoxia. *, significantly different than control (uninjured); #, hypoxic data are different than corresponding baseline data point.
Discussion
To date, the long-term impact of C2HS and the sCPP on breathing behavior in awake animals have not been fully evaluated. It was previously reported that the sCPP had little effect on ventilation at 5-week post-injury (Fuller et al., 2006). However, others have reported progressive improvements in diaphragm EMG activity over longer post-C2HS intervals (Nantwi et al., 1999). In the present study we assessed recovery in the context of respiratory challenge since previous (Fuller et al., 2006) and current data show that ventilation appears relatively normal after C2HS under baseline (i.e., normoxic) conditions. In this hemisection model, respiratory deficits become more evident during chemical challenge (Fuller et al., 2006). Conditions of high respiratory drive (i.e., induced by hypoxia or hypercapnia) are also known to activate the sCPP (Goshgarian, 2003; Golder et al., 2003).
Our present findings indicate that by 12-week post-injury ventilation in C2HS rats under hypercapnic stress begins to approximate values observed in uninjured, control animals. The sCPP, therefore, appears to play a candidate role in contributing to improved breathing under conditions of elevated respiratory drive as previously observed in anesthetized and vagotomized rats (Golder et al., 2003). This improvement, however, only becomes evident when injured and control rats of similar body weight are compared. Our data also suggest that endogenous neuroplasticity in the phrenic motor system after C2HS remains insufficient to promote full respiratory recovery as evidenced by fact that tidal volume remains below control values at 12-week post-injury.
Progressive changes in ventilation after C2 hemisection
An underlying logic of this investigation was that if the sCPP strengthens over weeks to months post-injury, there should be corresponding progressive changes in breathing patterns as reflected by plethysmographic measures. Specifically, the rapid shallow compensatory pattern of breathing observed after C2HS (Fuller et al., 2006) should progressively lessen with increasing ipsilateral diaphragm performance. As envisioned, when ventilation was assessed during a hypercapnic breathing challenge, the pattern of breathing was closer to control values at 12 compared to 2-week post-injury (Fig. 3). We also examined the appearance of spontaneous augmented breaths or “sighs” after C2HS. Augmented breaths help to prevent lung atelectasis, and require an increase in respiratory drive (Sieck and Fournier 1989; Golder et al., 2001a). Similar to a prior report (Golder et al., 2001a), we noted that augmented breaths occur more frequently after C2HS, and this adaptation does not appear immediately after the injury. There was a suggestion of a progressive recovery in augmented breath volume (Fig. 5), but in contrast to the hypercapnic tidal volume data (Fig. 3), the relative differences between control and C2HS rats are not statistically significant between 2- and 12-week post-injury. However, Golder et al. (2003) previously reported that anesthetized rats with a dual injury of C2HS and IL phrenicotomy (i.e. preventing crossed phrenic impulses from reaching the diaphragm) have a reduced volume of augmented breaths compared to C2HS alone indicating that the sCPP can contribute to augmented breath volume.
Re-emergence of ipsilateral phrenic motoneuron activity
In the current study, we observed sCPP activity by phrenic neurogram recordings at 2-week post-injury in the majority (75%) of rats. This early onset is consistent with other reports (Fuller et al., 2003, 2006; Golder and Mitchell, 2005). However, other studies have noted a much longer delay between C2HS and the re-appearance of inspiratory IL PhMN activity. For example, Nantwi et al. (1999) and Golder et al. (2001b) reported that sCPP is not observed until 1-2 month post-C2HS, whereas Vinit et al. (2006) observed that the sCPP was absent in the majority of rats they studied even at 3 month post-C2HS. Gender is the most immediately recognizable variable that could account for this disparity since protracted appearance of the sCPP was consistently observed in female rats (e.g., Nantwi et al., 1999; Golder et al., 2001). In contrast, our current and prior studies were done using male rats. Other conditions, however, may be involved in regulating the recovery of IL PhMN activity since we have recently noted that the onset of the sCPP is similar (i.e., 2-week post-C2HS) in male and female rats using the same neurophysiological conditions described here (Doperalski et al., 2007). However, to our knowledge the impact of gender on the robustness of the sCPP has not been carefully assessed at intervals longer than 2-week post-C2HS.
Consistent with our plethysmographic findings, phrenic neurogram recordings indicated a progressive increase in the ability to augment IL PhMN output during a respiratory challenge (i.e., hypoxia) after C2HS (Fig. 7). These results conform to the hypothesis that endogenous “self-repair” mechanisms associated with the sCPP can impact breathing in the awake, freely behaving rat after chronic C2HS. It should be mentioned, however, that our hypoxia data contrast with prior reports (Golder and Mitchell, 2005; Fuller et al., 2006), but this appears to be due to different approaches employed. For example, Golder and Mitchell (2005) found that the IL phrenic hypoxic response did not change 2-8-week post-C2HS. In that study, the hypoxic response was expressed relative to IL phrenic activity during a subsequent hypercapnic challenge. Unfortunately, similar indexing of the hypoxic response to hypercapnia was not possible in the present investigation thus eliminating an opportunity to make direct comparisons. In another study, we (Fuller et al., 2006) observed that the IL phrenic response to hypercapnia was actually lower at 4- vs. 2-week post-injury in a similar neurophysiological preparation; however, hypoxic responses were not assessed. Thus, the observed recovery of the IL phrenic bursting after C2HS may be substantially different during respiratory stimulation with hypoxia vs. hypercapnia in the vagotomized and ventilated rat preparation.
The sCPP substrate
Neuroanatomical tracing studies suggest that following C2HS, IL PhMNs are recruited via activation of contralateral bulbospinal neurons whose axons cross the spinal midline caudal to C2 (reviewed in Goshgarian, 2003). Current understanding of spinal synaptic circuitry associated with the sCPP derives from ultrastructural studies describing changes in axodendritic synaptology within the IL PhMN pool. For example, an increase in the number of dendro-dendritic appositions and synaptically active zones is observed in the IL phrenic motor nucleus a few hours post-C2 hemisection (Goshgarian et al., 1989; Goshgarian, 2003). There is also one report suggesting that the overall soma size of IL phrenic motoneurons decreases after C2HS (preliminary evidence cited in Mantilla and Sieck, 2003). Recent preliminary experiments raise the possibility that other cells (e.g. spinal interneurons) may be anatomically positioned to modulate phrenic motoneuron activity at the spinal level (Sandhu et al., 2006; Lane et al., 2007). This may be relevant to an interesting feature demonstrated by the present neurogram recordings which indicated that recovered IL PhMN activity was not entirely in phase with contralateral PhMN inspiratory activity. The onset of bursting was delayed by approximately 100 ms, whereas in normal animals, the initiation of right and left inspiratory discharges was more synchronous (e.g. Fig. 10). This difference in firing onset was most apparent during baseline conditions; a much smaller difference was observed during hypoxic challenge.
Data reported in El-Bohy and Goshgarian (1999) suggest that the crossed pathway responsible for return of IL PhMN function may predominantly project to larger, later-firing motoneurons. The present findings may thus reflect that under normoxic conditions, smaller early-firing cells may not be recruited as effectively as they could be when respiratory drive is enhanced, such as during hypoxic episodes. Other variables also may account for this post-injury change which could introduce additional dimensions to neuroplasticity dynamics in this system. Detailed cross-correlation analyses are in progress to further delineate this novel feature of recovered IL PhMN activity.
Implications
Approximately one half of spinal cord injuries occur in the cervical region (Winslow and Rozovsky, 2003) which with rare exception produce some degree of respiratory insufficiency and related complications — the severity of which correlates with the specific cervical spinal level of the injury (Winslow and Rozovsky, 2003). Although clinically relevant in cases of asymmetric (i.e., lateralized) high cervical contusion injuries, C2HS primarily provides a proof-of-principle model for respiratory resiliency in the phrenic motor system. Our findings suggest that endogenous “self-repair” mechanisms associated with and/or triggered by normal tidal breathing are sufficient to enhance respiratory recovery. However, other features of respiratory function remain poorly restored, and a systematic program of rehabilitation, pharmacological intervention, and/or combined regenerative/axonal sprouting strategies may be required to optimize plasticity and respiratory motor recovery. The fact that IL PhMN output can be significantly enhanced by a variety of surgical (e.g. cervical dorsal rhizotomy; Goshgarian, 1981; Fuller et al., 2002), chemical (e.g. hypoxia, Fuller et al., 2003; Golder and Mitchell, 2005) and pharmacological (Nantwi and Goshgarian, 2001, 2002, 2005; Alilain et al., 2007) manipulations suggests that these cells are capable of greater recruitment.
Acknowledgments
This work was funded by the Christopher Reeve Paralysis Foundation and the American Paraplegia Society (DDF) and NIH, NINDS, NS054025 (PJR). We thank Ms. Sandy Walker and Mr. Kevin Siegel for their technical assistance.
References
- Alilain WJ, Horn KP, Dick TE, Silver JE. Neuroscience Meeting Planner. Society for Neuroscience; San Diego, CA: 2007. Chondroitinase ABC treatment following C2 hemisection results in dramatically enhanced ipsilateral hemi-diaphragmatic recovery. Online. [Google Scholar]
- Cherniack NS, von Euler C, Glogowska M, Homma I. Characteristics and rate of occurrence of spontaneous and provoked augmented breaths. Acta Physiol. Scand. 1981;111:349–360. doi: 10.1111/j.1748-1716.1981.tb06747.x. [DOI] [PubMed] [Google Scholar]
- Doperalski NJ, Fuller DD. Long-term facilitation of ipsilateral but not contralateral phrenic output after cervical spinal cord hemisection. Exp. Neurol. 2006;200(1):74–81. doi: 10.1016/j.expneurol.2006.01.035. [DOI] [PubMed] [Google Scholar]
- Doperalski NJ, Reier PJ, Fuller DD. Neuroscience Meeting Planner. Society for Neuroscience; 2007. Sex differences in respiratory recovery following high cervical spinal hemisection in rats. Available online at http://www.sfn.org/am2007/ [Google Scholar]
- Drorbaugh JE, Fenn WO. A barometric method for measuring ventilation in newborn infants. Pediatrics. 1955;16:81–87. [PubMed] [Google Scholar]
- El-Bohy AA, Goshgarian HG. The use of single phrenic axon recordings to assess diaphragm recovery after cervical spinal cord injury. Exp. Neurol. 1999;156(1):172–179. doi: 10.1006/exnr.1999.7013. [DOI] [PubMed] [Google Scholar]
- Fuller DD, Johnson SM, Johnson RA, Mitchell GS. Chronic cervical spinal sensory denervation reveals ineffective spinal pathways to phrenic motoneurons in the rat. Neurosci. Lett. 2002;323(1):25–28. doi: 10.1016/s0304-3940(02)00121-0. [DOI] [PubMed] [Google Scholar]
- Fuller DD, Johnson SM, Olson EB, Jr., Mitchell GS. Synaptic pathways to phrenic motoneurons are enhanced by chronic intermittent hypoxia after cervical spinal cord injury. J. Neurosci. 2003;23(7):2993–3000. doi: 10.1523/JNEUROSCI.23-07-02993.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fuller DD, Golder FJ, Olson EB, Jr., Mitchell GS. Recovery of phrenic activity and ventilation following cervical spinal hemisection in rats. J. Appl. Physiol. 2006;100(3):800–806. doi: 10.1152/japplphysiol.00960.2005. [DOI] [PubMed] [Google Scholar]
- Golder FJ, Mitchell GS. Spinal synaptic enhancement with acute intermittent hypoxia improves respiratory function after chronic cervical spinal cord injury. J. Neurosci. 2005;25(11):2925–2932. doi: 10.1523/JNEUROSCI.0148-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Golder FJ, Reier PJ, Davenport PW, Bolser DC. Cervical spinal cord injury alters the pattern of breathing in anesthetized rats. J. Appl. Physiol. 2001a;91(6):2451–2458. doi: 10.1152/jappl.2001.91.6.2451. [DOI] [PubMed] [Google Scholar]
- Golder FJ, Reier PJ, Bolser DC. Altered respiratory motor drive after spinal cord injury: supraspinal and bilateral effects of a unilateral lesion. J. Neurosci. 2001b;21:8680–8689. doi: 10.1523/JNEUROSCI.21-21-08680.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Golder FJ, Fuller DD, Davenport P, Johnson RD, Reier PJ, Bolser DC. Respiratory motor recovery after unilateral spinal cord injury: eliminating crossed phrenic activity decreases tidal volume and increases contralateral respiratory motor output. J. Neurosci. 2003;23:2494–2501. doi: 10.1523/JNEUROSCI.23-06-02494.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Golder FJ, Davenport PW, Johnson RD, Reier PJ, Bolser DC. Augmented breath phase volume and timing relationships in the anesthetized rat. Neurosci. Lett. 2005;373(2):89–93. doi: 10.1016/j.neulet.2004.09.063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goshgarian HG. The role of cervical afferent nerve fiber inhibition of the crossed phrenic phenomenon. Exp. Neurol. 1981;72:211–225. doi: 10.1016/0014-4886(81)90139-4. [DOI] [PubMed] [Google Scholar]
- Goshgarian HG. The crossed phrenic phenomenon: a model for plasticity in the respiratory pathways following spinal cord injury. J. Appl. Physiol. 2003;94:795–810. doi: 10.1152/japplphysiol.00847.2002. [DOI] [PubMed] [Google Scholar]
- Goshgarian HG, Moron MF, Prcevski P. Effect of cervical spinal cord hemisection and hemidiaphragm paralysis on arterial blood gases, pH, and respiratory rate in the adult rat. Exp. Neurol. 1986;93(2):440–445. doi: 10.1016/0014-4886(86)90206-2. [DOI] [PubMed] [Google Scholar]
- Goshgarian HG, Yu XJ, Rafols JA. Neuronal and glial changes in the rat phrenic nucleus occurring within hours after spinal cord injury. J. Comp. Neurol. 1989;284(4):519–533. doi: 10.1002/cne.902840404. [DOI] [PubMed] [Google Scholar]
- Lane MA, White TE, Coutts MA, Sandhu MS, Siegel KA, O'Steen BE, Fuller DD, Reier PJ. Neuroscience Meeting Planner. Society for Neuroscience; San Diego, CA: 2007. Plasticity in respiratory circuitry following partial spinal cord injury (SCI) in the adult rat. 2007. Online. [Google Scholar]
- Lipski J, Zhang X, Kruszewska B, Kanjhan R. Morphological study of long axonal projections of ventral medullary inspiratory neurons in the rat. Brain Res. 1994;640:171–184. doi: 10.1016/0006-8993(94)91871-6. [DOI] [PubMed] [Google Scholar]
- Mantilla CB, Sieck GC. Mechanisms underlying motor unit plasticity in the respiratory system. J. Appl. Physiol. 2003;94:1230–1241. doi: 10.1152/japplphysiol.01120.2002. [DOI] [PubMed] [Google Scholar]
- Nantwi KD, Goshgarian HG. Alkylxanthine-induced recovery of respiratory function following cervical spinal cord injury in adult rats. Exp. Neurol. 2001;168(1):123–134. doi: 10.1006/exnr.2000.7581. Mar. [DOI] [PubMed] [Google Scholar]
- Nantwi KD, Goshgarian HG. Actions of specific adenosine receptor A1 and A2 agonists and antagonists in recovery of phrenic motor output following upper cervical spinal cord injury in adult rats. Clin. Exp. Pharmacol. Physiol. 2002;29:915–923. doi: 10.1046/j.1440-1681.2002.03750.x. [DOI] [PubMed] [Google Scholar]
- Nantwi KD, Goshgarian HG. Adenosinergic mechanisms underlying recovery of diaphragm motor function following upper cervical spinal cord injury: potential therapeutic implications. Neurol. Res. 2005;27(2):195–205. doi: 10.1179/016164105X21977. Mar. [DOI] [PubMed] [Google Scholar]
- Nantwi K, El-bohy AA, Schrimsher GW, Reier PJ, Goshgarian HG. Spontaneous recovery in a paralyzed hemidiaphragm following upper cervical spinal cord injury in adult rats. Neurorehabil. Neural Repair. 1999;13:225–234. [Google Scholar]
- Sandhu MS, Doperalski NJ, Bolser DC, Reier PJ, Fuller DD. Neuroscience Meeting Planner. Society for Neuroscience; Atlanta, GA: 2006. Correlation of ipsilateral (IL) versus contralateral (CL) phrenic inspiratory waveforms after chronic C2 cervical hemisection (C2HS) in rats. 2006. Online. [Google Scholar]
- Sieck GC, Fournier M. Diaphragm motor unit recruitment during ventilatory and nonventilatory behaviors. J. Appl. Physiol. 1989;66(6):2539–2545. doi: 10.1152/jappl.1989.66.6.2539. [DOI] [PubMed] [Google Scholar]
- Choi H, Liao WL, Newton KM, Onario RC, King AM, Desilets FC, Woodard EJ, Eichler ME, Frontera WR, Sabharwal S, Teng YD. Respiratory abnormalities resulting from midcervical spinal cord injury and their reversal by serotonin 1A agonists in conscious rats. J. Neurosci. 2005;25(18):4550–4559. doi: 10.1523/JNEUROSCI.5135-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vinit S, Gauthier P, Stamegna JC, Kastner A. High cervical lateral spinal cord injury results in long-term ipsilateral hemidiaphragm paralysis. J. Neurotrauma. 2006;23(7):1137–1146. doi: 10.1089/neu.2006.23.1137. [DOI] [PubMed] [Google Scholar]
- Winslow C, Rozovsky J. Effect of spinal cord injury on the respiratory system. Am. J. Phys. Med. Rehabil. 2003;82(10):803–814. doi: 10.1097/01.PHM.0000078184.08835.01. [DOI] [PubMed] [Google Scholar]
- Zimmer MB, Nantwi K, Goshgarian HG. Effect of spinal cord injury on the respiratory system: basic research and current clinical treatment options. J. Spinal Cord Med. 2007;30(4):319–330. doi: 10.1080/10790268.2007.11753947. [DOI] [PMC free article] [PubMed] [Google Scholar]