

Abstract
Triggering receptor expressed on myeloid cells-2 (TREM-2) is an innate immune receptor that initiates cellular activation upon ligation. In this study, we examined the interaction of TREM-2 with Neisseria gonorrhoeae using murine TREM-2A as it has been reported to recognize bacterial ligands. Using a whole bacteria ELISA, TREM-2A bound to all 6 strains in variable degrees. Far western blots of gonococcal outer membranes revealed TREM-2A binding to lipooligosaccharide (LOS) and Opa protein with predominant binding to LOS. Binding of TREM-2A to LOS was confirmed by ELISA and surface plasmon resonance. O-deacylation of the lipid A significantly reduced binding. Flow cytometry and reporter cell assays showed that gonococci bound to TREM-2A-transfected cells and induced transmembrane signaling. In humans, TREM-2 was constitutively expressed by genitourinary and fallopian tube epithelial cells, both of which are primary targets of gonococcal invasion. Ligation of TREM-2 by LOS induced IL-6 production in HeLa cervical carcinoma cells. To our knowledge, this is the first report of the expression of human TREM-2 by cells deriving from a non-myeloid lineage. We conclude that gonococci can interact with TREM-2 receptors through binding to LOS and Opa protein and initiate cell signaling and cytokine production.
Keywords: innate immunity, TREM-2, gonorrhea, epithelial cell, lipooligosaccharide
Introduction
Neisseria gonorrhoeae remains a major cause of sexually transmitted infections around the world (1). In 10 to 20 percent of women with gonococcal infection, pelvic inflammatory disease develops which includes a variety of inflammatory conditions of the upper reproductive tract including endometritis, parametritis, and salpingitis and often leads to chronic pelvic pain, infertility, and ectopic pregnancy (2). In addition, epidemiologic and clinical studies provide strong evidence that gonococcal infections facilitate the transmission of HIV-1 infection and that control of gonorrhea needs to remain high on the agenda of HIV-1 control programs (3-6).
Evidence is growing that the innate immune system plays a critical role in the initial response to gonococcal infection. Our previous work and that of other investigators has shown that gonococci can initiate proinflammatory cytokine expression through activation of NFκB. In particular, gonococci engage innate immune receptors TLR4 and TLR2 with lipooligosaccharide (LOS) engaging TLR4, and PorB and lipoprotein Lip functioning as agonists for TLR2 (7-9). In urogenital epithelial cell lines, gonococcal infection induces up-regulation of a variety of proinflammatory and inflammatory cytokines, including TNF-α, IL-1β, IL-6, and IL-8 (10-12). In experimental infection, the levels of the same four cytokines have been found to be elevated in both the urine and plasma of men after gonococcal challenge (13). In addition, McGee et al. demonstrated that gonococcal infection of human fallopian tube mucosa resulted in increased production of TNF-α, and that TNF-α concentrations correlated with the sloughing of fallopian tube ciliated cells in organ explant cultures in vitro (14), which closely mimics the progression of gonococcal infection observed in vivo.
Among the important virulence factors involved in the pathogenesis of gonococcal infections, the LOS has been identified as a major component inducing the host proinflammatory cytokine response to the organisms (7). Gonococcal LOS is structurally related to the lipopolysaccharide (LPS) expressed by enteric Gram-negative bacteria, although LOS does not possess the repeating O-polysaccharide regions (15, 16). LOS is composed of a hydrophilic oligosaccharide core which is exposed on the surface, and a hydrophobic lipid A portion containing fatty acid residues that vary in acylation and phosphorylation which anchors the LOS into the bacterial outer membrane (17). In particular, several studies have implicated the lipid A portion as the bioactive component of Neisserial LOS (18, 19).
Triggering receptor expressed on myeloid cells (TREM) is a recently described family of transmembrane glycoproteins that play an important role in initiating innate and adaptive immune responses (20). The two best characterized of these receptors, TREM-l and TREM-2, function through association with a DAP12-mediated pathway for signaling (21, 22). In humans, both the TREM-1 and TREM-2 genes are highly conserved with only a single gene of each receptor having been identified. In mice, TREM-1 is conserved whereas TREM-2 is expressed as two isoforms, TREM-2A and TREM-2B, which differ by three amino acids in the extracellular domain (23). To date, TREM-1 has been found to be expressed by neutrophils and a subset of monocytes and macrophages, whereas the expression of TREM-2 has been reported on immature monocyte-derived dendritic cells, osteoclasts, macrophages, and microglia (20). The ligands for the TREM receptors remain unclear. Although LPS has been shown to be an agonist for TREM-1 activation (24), the direct binding of LPS to TREM-1 has yet to be demonstrated. The recent report of Daws et al. suggests that murine TREM-2 recognizes bacteria through a charge-dependent interaction (25). They found that both soluble and membrane-expressed TREM-2 bound a variety of bacteria including Escherichia coli and Staphylococcus aureus, and that this binding was inhibitable by LPS and other charged carbohydrates of bacterial origin.
In this study, we demonstrate that murine TREM-2 binds to gonococcal outer membrane components LOS and Opa protein which are important virulence determinants of infection, and that binding can initiate transmembrane cell signaling. In addition, we provide evidence that human TREM-2 is constitutively expressed by genitourinary epithelial cells and by the fallopian tube epithelium, both of which are primary targets of gonococcal invasion, and that ligation of TREM-2 by LOS can induce IL-6 production in HeLa cervical carcinoma cells. To our knowledge, this is the first report of the expression of human TREM-2 by cells deriving from a non-myeloid lineage.
Methods
Bacterial strains and LOS
Six strains of N. gonorrhoeae (F62, 1291, GC56, and MS11mk LOS variants C [MkC], A [MkA], and MkC Δ1-5) were studied and have been described previously (7, 26, 27). MkC Δ1-5 is an isogenic mutant of MkC expressing LOS containing a truncated OS α chain (26). LOS was isolated by hot phenol extraction and outer membrane complexes (OMC) containing both LOS and proteins were prepared as described previously (28, 29). For some experiments, LOS was O-deacylated with anhydrous hydrazine as reported previously (30), which removes the O-linked β-hydroxylaurate acyl chains of lipid A while leaving the two N-linked β-hydroxymyristate chains intact. The OS structures of the LOS α chains have been determined previously and are shown in Table 1; none of the strains for which the structures are known express β-chain (26, 27). All strains were maintained for expression of the opacity (Opa) protein by using a dissecting microscope and standard selection criteria, and expression was confirmed by electrophoretic and immunoblotting analyses of whole-cell lysates as described previously (31).
Table 1.
LOS α Chain Structures of Gonococcal Strains
| LOS | α Chains |
|---|---|
| MkCa | Galβ1-4GlcNAcβ1-3Galβ1-4GlcNAcβ1-3Galβ1-4Glc |
| MkC | GalNAcβ1-3Galβ1-4GlcNAcβ1-3Galβ1-4Glc |
| MkC | Galβ1-4GlcNAcβ1-3Galβ1-4Glc |
| F62b | GalNAcβ1-3Galβ1-4GlcNAcβ1-3Galβ1-4Glc |
| F62 | Galβ1-4GlcNAcβ1-3Galβ1-4Glc |
| 1291 | Galβ1-4GlcNAcβ1-3Galβ1-4Glc |
| MkA | Galβ1-4-Glc |
| MkC Δ1-5c | Galβ1-4-Glc |
| GC56 | NDd |
MkC expresses three major LOS components.
F62 has two main LOS components.
MkC Δ1-5 is an isogenic mutant of variant MkC; MkC and MkA are variants of strain MS11mk.
ND, not determined.
Cell lines
Cervical carcinoma cell lines ME-180 and HeLa, monocytic cell line THP-1, and three HPV16/E6E7 immortalized reproductive tract epithelial cell lines, endocervical End1/E6E7, ectocervical Ect1/E6E7, and vaginal Vk2/E6E7, were obtained from the American Type Culture Collection (Manassas, VA) and have been characterized previously (7, 10, 12). The E6E7 cell lines were maintained in keratinocyte serum-free medium (Invitrogen, Carlsbad, CA) supplemented with 0.05mg/ml bovine pancreatic extract, 0.1 ng/ml recombinant human epidermal growth factor, and 0.44 μg/ml CaCl2. ME-180 and THP-1 cells were cultured in McCoy's 5A medium and RPMI 1640, respectively, containing 10% fetal bovine serum (FBS). HeLa cells were cultured in Eagle's MEM supplemented with 10% FBS, 0.1 mM nonessential amino acids, 0.1 mM HEPES, and 2 mM l-glutamine. The mouse T cell lymphoma cell line BWZ.36 (BWZ) was kindly provided by N. Shastri (University of California, Berkeley, CA). This line does not express endogenous TREM receptors (25). BWZ cells were cultured in RPMI medium containing 5% FBS, 25 μM 2-mercaptoethanol, and 2 mM glutamine.
Production of soluble TREM-2A and TREM-3 and expression on BWZ cells
Murine TREM-2A and TREM-3 were produced as fusion proteins with the human IgG1 Fc domain in 293T cells and purified from conditioned medium using protein G affinity chromatography as described previously (25). For expression of TREM-2A and TREM-3 on BWZ cells, a vector was constructed containing a CD8 leader sequence and FLAG tag, and downstream of this, a CD8 transmembrane domain and the CD3ζ cytoplasmic domain as reported previously (25). The cDNA encoding amino acids 19-171 of TREM-2A and amino acids 20-135 of TREM-3 were cloned between the FLAG tag and CD8 transmembrane domain. BWZ cells were transfected by electroporation and selected with 0.75 mg/ml Zeocin. Expression of TREM-2A and TREM-3 on BWZ cells was confirmed by flow cytometry using affinity-purified rabbit anti-FLAG antibody (Sigma-Aldrich, St. Louis, MO).
Whole Bacteria ELISA
TREM-2A and TREM-3 binding to whole bacteria as well as TREM-2A binding to LOS was investigated using a series of ELISA assays. In addition, the binding of human IgG Fc fragments (Bethyl Laboratories, Montgomery, TX) to whole bacteria was determined as a control for the binding of the IgG Fc portion of the TREM-2A and TREM-3 fusion proteins to bacteria. Microtiter wells were coated overnight with either 75 μl of a suspension of bacteria of absorbance 0.1 at 650 nm or 75 μl of 25 μg/ml LOS. The wells were then blocked with 1% BSA-TBS and incubated with 75 μl of 1 μg/ml of TREM-2A, TREM-3, or IgG Fc fragments. The binding of TREM-2A, TREM-3, and IgG Fc fragments was detected with anti-human IgG Fc antibody conjugated with alkaline phosphatase (Sigma-Aldrich) and p-nitro-phenyl phosphate substrate, and the absorbance was read at 405 nm with a MAXline microplate reader.
Far western and dot blots
For far western analyses, the proteins and LOS in purified gonococcal outer membrane complexes were separated by 12% SDS-PAGE, electroblotted to nitrocellulose, and blocked in 1% BSA-TBS as described previously (32). The membranes were then incubated with 1 μg/ml of TREM-2A or TREM-3 and binding was detected with anti-human IgG Fc antibody conjugated with alkaline phosphatase followed by a substrate solution of 50 mM Tris-HCl (pH 8.0), 0.1% Naphthol AS-MX phosphate disodium salt, and 0.2% fast red TR salt. The identity of LOS was determined with the use of LOS MAb 1B2, which binds to LOS expressing a terminal lacto-N-neotetraose (Galβ1-4GlcNAcβ1-3Galβ1-4Glc) structure (33), and that of Opa proteins was determined with the use of Opa MAb B33, which reacts with all Opa protein variants (31). For dot blots, LOS was spotted onto Hybond nitrocellulose membranes, which were then blocked with 1X DIG Block buffer (Roche, Indianapolis, IN) containing 0.1% Tween and incubated with 1 μg/ml TREM-2A. Binding of TREM-2A to LOS on the membranes was detected as described for the far western blots.
Surface plasmon resonance studies
Surface plasmon resonance analyses were performed on a Biacore 3000 system using CM5 sensor chips (Biacore Inc., Piscataway, NJ), as described previously (34). TREM-2A and TREM-3 were diluted in 10 mM sodium acetate buffer at pH 4.5 to a final concentration of 10 μg/ml and immobilized on the surface of the chip using the amine coupling method. The carboxyl groups on the sensor surfaces were activated with a mixture of 0.2 M N-ethyl-N'-(3-diethylamino-propyl) carbodiimide (EDC) and 0.05 M N-hydroxysuccinimide (NHS), and the protein was coupled under the continuous flow of HBS-EP buffer (10mM Hepes with 0.15 M NaCl, 3.4 mM EDTA and 0.005% surfactant P20, pH 7.4). A flow cell containing an unmodified dextran surface treated with EDC and NHS and not exposed to the coupling protein served as the control flow cell surface for each chip. Residual reactive groups were blocked with 1.0 M ethanolamine hydrochloride at pH 8.5 in both control and test flow cell surfaces. Immobilization levels of 540 and 330 resonance units were achieved for TREM-2A and TREM-3, respectively.
For kinetic binding determinations, 1291 LOS was diluted in HBS-EP buffer to give a series of increasing concentrations, injected over the test and control flow cell surfaces at 10 μl/min for 1500 s, and then dissociated from TREM-2A and TREM-3 by washing with buffer alone. Chip surfaces were regenerated with 100 sec injections of 0.05% SDS. For some experiments, equimolar concentrations of 1291 LOS and soluble TREM-2A were incubated for 30 min prior to injection over the TREM-2A and control flow cell surfaces.
Data transformation and overlay plots were prepared with BIA evaluation software (version 3.1). Binding signals were corrected for non-specific binding by subtracting the signal of the control flow cell. The affinities of the binding interactions between TREM-2A, TREM-3 and 1291 LOS were calculated by using the 1:1 Langmuir binding model in the BIAevaluation software.
Flow cytometry
For studies of bacterial binding to BWZ cells expressing TREM-2A or TREM-3, gonococci were labeled with CellTrace Far Red DDAO-SE (Invitrogen, Carlsbad, CA) and incubated with cells at a bacteria to cell ratio of 20:1. After removal of unbound bacteria by washing, bacterial binding to cells was detected by analyzing cells for associated fluorescence using a FACScan flow cytometer. For flow cytometric analyses of the membrane expression of human TREM-2 by all cell lines except HeLa, 5×105 cells were incubated for 60 min on ice with 1.5 μg/ml of anti-human TREM-2 MAb (R&D Systems, Minneapolis, MN) in a total volume of 100 μl of PBS containing 0.1% BSA. After 2 washes with PBS/BSA, the cells were incubated for 30 min on ice with 1 μg of FITC-conjugated rat anti-mouse IgG (eBioscience, San Diego, CA) in a total volume of 100 μl of PBS containing 0.1% BSA. The cells were washed twice with PBS/BSA, fixed with 3.7% glutaraldehyde, and analyzed by flow cytometry. In controls, the cells were incubated with an irrelevant mouse IgG2B isotype immunoglobulin (R&D Systems). For HeLa cells, membrane expression of TREM-2 was detected as above using affinity-purified goat anti-human TREM-2 polyclonal IgG and phycoerythrin-conjugated donkey anti-goat IgG (R&D Systems).
Reporter cell assay
The BWZ cell line was derived from BW5147 T cells and contained a lacZ reporter gene as described previously (35). The cells were transfected to surface-express TREM-2A or TREM-3 linked to a CD3ζ cytoplasmic domain. Signaling through CD3ζ activated four NFAT promoter elements which induced expression of the lacZ reporter gene. For the assay, nontransfected BWZ cells or BWZ cells transfected with either TREM-2A or TREM-3 were seeded in 96-well plates at 1 × 105 cells/well in media supplemented with 10 ng/ml phorbol 12-myristate 13-acetate (PMA). For stimulation with gonococci, reporter cells were incubated with gonococci at a bacteria to cell ratio of either 1:1 or 100:1. For LOS, TREM-2A-expressing reporter cells were treated with LOS concentrations of either 1 μg/ml or 100 μg/ml. Plates were then incubated for 16 h at 37°C in 5% CO2. Cells were washed once in PBS and lacZ activity was determined by incubating the cells with 150 μM chlorophenol red-β-D-galactopyranoside in PBS supplemented with 100 mM 2-mercaptoethanol, 9 mM MgCl2, and 0.125% Nonidet P-40. After sufficient color development, absorbance was measured at 595 nm and corrected for background absorbance at 650 nm. Following subtraction of the absorbance of wells treated with PMA alone, values were calculated as the fold increase in lacZ activity in TREM-2A- or TREM-3-expressing cells treated with either gonococci or LOS as compared with nontransfected cells treated similarly.
Immunohistochemistry
Fallopian tube tissue was obtained from women undergoing elective hysterectomies as described previously (36). The fallopian tubes were cleaned of adventitial tissue, cut into 2 mm2 square pieces and incubated overnight in MEM/F-12 supplemented with 10% FBS. The tissues were placed on a 10% formalin soaked sponge, fixed in formalin and processed for paraffin embedding and sectioning. The tissue was sectioned to 3 μm, deparaffinized in SafeClear II and rehydrated in 100, 95, and 70% EtOH followed by a 2 minute wash in distilled H2O. The slides containing tissue were incubated in methanol containing 3% H2O2 for 30 minutes. Following fixation, the slides were rinsed in PBS (pH 7.4), placed in sodium citrate buffer (10 mM) containing 0.05% Tween 20, pH 6.0 and heated in a microwave oven for 3 minutes at low power setting. The solution was allowed to cool for 1 minute and then microwaved for an additional 3 minutes. The solution containing the slides was allowed to cool for 15 min to return to ambient temperature. The slides were rinsed twice in distilled water and then washed twice in PBS for 5 minutes each. TREM-2 staining was carried out using the Cell and Tissue HRP-DAB staining system according to the recommendation of the manufacturer (R&D Systems). Sections were incubated with 20 μg/ml of affinity-purified goat anti-human TREM-2 polyclonal IgG (R&D Systems) for 60 minutes at room temperature, rinsed, and then incubated with the biotinylated secondary antibody for 40 min. After rinsing, sections were incubated with streptavidin-conjugated horseradish peroxidase for 30 min followed by substrate development with diaminobenzidine. Stained sections were counterstained with hematoxylin for 75 seconds, mounted with Permount, coversliped, and allowed to dry overnight prior to observation. Negative control slides were incubated with non-immune goat IgG (R&D Systems). This research was approved by the Springfield Committee for Research Involving Human Subjects.
IL-6 ELISA
HeLa cells were seeded in 24-well plates at a concentration of 1 × 105 cells/well and incubated to confluency. Cells were treated with LOS from strain 1291 at concentrations of 10 and 100 ng/ml for 12 hours at 37°C. Control wells were treated with culture media only. For some experiments, cells were treated with 100 μl of affinity-purified goat anti-human TREM-2 polyclonal IgG (R&D Systems) at a concentration of 2.5 μg/ml for 30 min at 4°C prior to addition of the LOS. Supernatants from the LOS-treated and untreated cells were assayed for IL-6 using an IL-6 ELISA kit as recommended by the manufacturer (eBioscience, San Diego, CA).
Statistical analysis
Groups of data were analyzed by the Tukey test for multiple pairwise comparisons. Values of p<0.05 were considered significant for all comparisons.
Results
ELISA analyses of TREM-2A binding
Whole bacteria ELISA analyses were performed to determine whether gonococci bound soluble recombinant TREM-2A and whether strains varied in their recognition and binding. As shown in Figure 1, TREM-2A bound to all six strains tested but to variable degrees (p = 0.01). In contrast, the binding of soluble recombinant TREM-3, a related member of the TREM family (37), to strains 1291 and F62 was significantly less than that of TREM-2A (p < 0.001) and was comparable to the binding of IgG Fc fragments used as a control for the binding of the IgG Fc portion of the TREM-2A and TREM-3 fusion proteins. The length of the OS α chain did not affect binding as MkC and MkC Δ1-5 bound similar amounts of TREM-2 despite MkC Δ1-5 expressing a truncated version of the OS α chain of MkC (Table 1).
Figure 1.

TREM-2A binds to various strains of N. gonorrhoeae. Whole-cell ELISA analyses were performed by coating microtiter wells with gonococci and incubating the wells with TREM-2A (TR-2A), TREM-3 (TR-3), or IgG Fc fragments. The bars represent the mean of triplicate data points and the error bars are ±SD. The results are representative of four independent experiments.
Far western blot analyses of TREM-2A binding
We next investigated the site of TREM-2A binding on the gonococcal membrane using far western blots to probe the binding to dissociated LOS and protein components of OMC purified from the highest and lowest binding strains 1291 and F62, respectively. The blots revealed that TREM-2A bound to two components of the gonococcal outer membrane, LOS and a protein with an approximate mass of 30 kDa, with >75% of the binding to LOS as determined by densitometry (Figure 2). The conclusion that LOS served as the primary TREM-2A binding site derived from the low molecular mass of the reactive band as well as from the binding of LOS MAb 1B2 to a band of comparable SDS-PAGE migration for both strains. As the size of the 30 kDa protein was appropriate for a gonococcal Opa protein, we used Opa MAb B33 to provide evidence for its identity as Opa. As with the LOS MAb, the Opa MAb bound to a band of comparable SDS-PAGE migration to that bound by TREM-2A, suggesting that the Opa protein is secondarily bound by TREM-2A. Interestingly, Opa MAb B33 revealed that strain F62 expresses two Opa variants, only one of which bound TREM-2A. As a control, the binding of TREM-3 was tested similarly, with no binding to any outer membrane components detectable.
Figure 2.
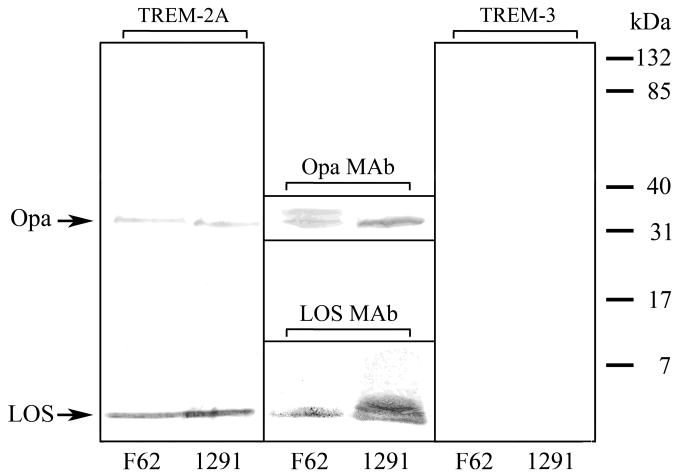
TREM-2A binds gonococcal outer membrane components LOS and Opa protein as shown by far western blots. The SDS-PAGE-separated LOS and proteins from OMC purified from gonococcal strains F62 and 1291 were transferred to nitrocellulose and reacted with TREM-2A (left panel) or TREM-3 (right panel). Both strains bound TREM-2A to outer membrane components LOS and Opa protein, however >75% of binding was to LOS as determined by densitometry. No binding of TREM-3 to outer membrane components was detected. The identity of LOS was determined with the use of LOS MAb 1B2 (lower center panel), which binds to the lacto-N-neotetraose terminal structure of the LOS expressed by strains F62 and 1291, and that of Opa proteins was determined with the use of Opa MAb B33 (upper center panel), which reacts with all Opa protein variants. For the LOS, intensity differences in LOS MAb staining are the result of differences between the two strains in the amount of LOS expressed terminating in lacto-N-neotetraose as shown in Table 1. For the Opa proteins, F62 expressed two variants, only one of which bound TREM-2A. The results are representative of three independent experiments.
TREM-2A binding to LOS
As the predominance of TREM-2A binding to gonococcal outer membrane components was to LOS, we further investigated this finding using LOS ELISA assays. As shown in Figure 3, we found that TREM-2A bound to purified LOS, thus confirming the results of the far western blots. The relative level of binding was variable and paralleled the degree to which TREM-2A bound to whole bacteria. Further evidence that OS structure was independent of the amount of binding was seen with the significant difference in the binding of TREM-2A to 1291 and F62 LOS (p < 0.001), both of which express a lacto-N-neotetraose OS α chain (Table 1). This suggested that the structure of the lipoidal moiety may play a role in binding. To assess this possibility, LOS was O-deacylated with anhydrous hydrazine which removes the O-linked β-hydroxylaurate acyl chains of lipid A and the binding of TREM-2A was determined by ELISA. We found that O-deacylated LOS bound significantly less TREM-2A than untreated LOS (p < 0.001 for all three LOS comparing untreated versus O-deacylated) (Figure 3). As it is possible that O-deacylation affected the binding of LOS to microtiter wells, we confirmed the ELISA results using a dot blot assay. As shown in Figure 3, dot blot analyses revealed that O-deacylated LOS bound substantially less TREM-2A than untreated LOS.
Figure 3.
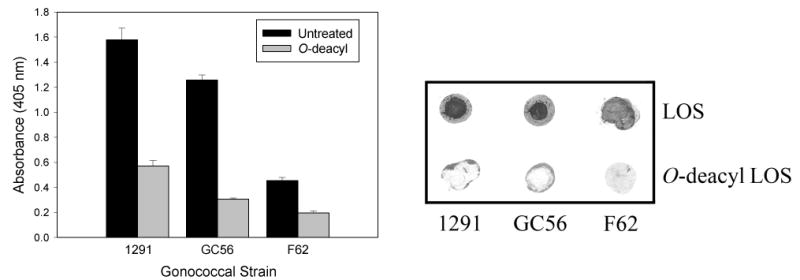
TREM-2A binds to intact gonococcal LOS but not to O-deacylated LOS. Left Panel: ELISA analyses of the binding of TREM-2A to intact and O-deacylated LOS. Wells were coated with LOS, blocked and reacted with TREM-2A. TREM-2A bound to the intact LOS of the strains tested but to variable degrees. No correlation was observed between the length of the OS α chains and TREM-2A binding based on the structures in Table 1. O-deacylated LOS bound substantially less TREM-2A than untreated LOS. Statistical comparisons: p < 0.001 for all three LOS comparing untreated versus O-deacylated. The bars represent the mean of triplicate data points and the error bars are ±SD. The data are representative of three independent experiments. Right Panel: Dot blot analyses of the binding of TREM-2A to LOS and O-deacylated LOS. LOS was spotted onto Hybond nitrocellulose membranes, which were blocked and incubated with TREM-2A. The significant decrease in binding of TREM-2A to O-deacylated LOS suggested a critical role for lipid A in the binding of TREM-2A to LOS. The data are representative of three independent experiments.
Surface plasmon resonance studies
To measure the affinity of TREM-2A for LOS, surface plasmon resonance studies were performed using 1291 LOS to which TREM-2A bound to the greatest degree in ELISA analyses. TREM-2A was immobilized on the surface of the chip, and the affinity of TREM-2A for LOS was determined following the injection of increasing concentrations of LOS. As shown in Figure 4, LOS showed specific and dose-responsive binding to TREM-2A. In contrast, no specific binding of LOS to TREM-3 was detected as exemplified by the horizontal sensorgram generated following injection of the highest concentration of LOS (62 μM) over the TREM-3-immobilized surface. The specificity of the recognition of TREM-2A by LOS was confirmed by incubating the highest concentration of LOS with an equimolar concentration of soluble TREM-2A prior to injection of the LOS. Such treatment effectively eliminated the binding of the LOS to TREM-2A as judged by the minimal increase in the slope of the association portion of the sensorgram following injection. To calculate the dissociation constant value (KD) for the observed interaction between TREM-2A and LOS, the one-to-one Langmuir model was used. The choice of this model was supported by a χ2 value of 2.88, indicating that the experimental curves fit well to the theoretical ideal curves of one-to-one binding interactions. Using this model, the KD of 1291 LOS for TREM-2A was calculated to be 3.5 × 10-7 M.
Figure 4.

Surface plasmon resonance kinetic analyses of the binding of 1291 LOS to TREM-2A performed on a Biacore 3000 system using CM5 sensor chips. Increasing concentrations of LOS were injected at 10 μl/min into a flow cell containing TREM-2A-coated and control surfaces. For TREM-3, the highest concentration of LOS (62 μM) was injected over the TREM-3-coated and control surfaces. All binding signals were corrected for non-specific binding by subtracting the signal of the control flow cell. For the sensorgram labeled as LOS and TREM-2A, equimolar concentrations of LOS and soluble TREM-2A were incubated for 30 min prior to injection over the TREM-2A and control surfaces.
Binding of gonococci to TREM-2A expressed on cells
We used BWZ cells transfected to express TREM-2A to determine whether gonococci could bind to TREM-2A expressed on the surface of cells. To this end, we incubated fluorescent-labeled gonococci with TREM-2A-expressing BWZ cells and analyzed bacterial binding to cells using flow cytometry. As shown in Figure 5, gonococci bound to the TREM-2A transfected cells to a greater degree than to nontransfected cells. As a control for the specificity of gonococcal binding to TREM-2A on BWZ cells, we also assessed the binding of gonococci to BWZ cells expressing TREM-3 and found that gonococci bound to a greater degree to TREM-2A-expressing cells than to cells expressing TREM-3 (Figure 5).
Figure 5.
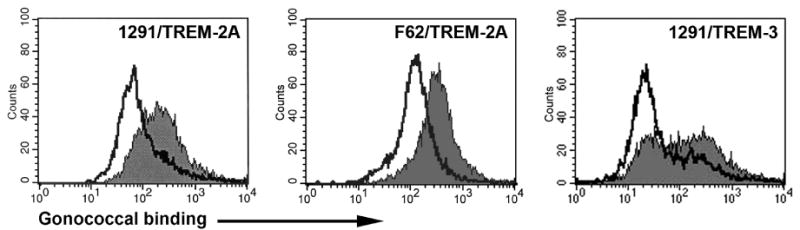
Gonococci bind to TREM-2A-transfected cells to a greater degree than to TREM-3-transfected or nontransfected cells. Gonococcal strains 1291 and F62 were fluorescently labeled and incubated with TREM-2A-expressing, TREM-3-expressing or nonexpressing BWZ cells. Flow cytometric analyses were performed to assess the degree of binding to each cell type. In the panels labeled TREM-2A, the filled histograms represent TREM-2A-expressing cells and the open histograms are nonexpressing BWZ cells. In the panel labeled TREM-3, the filled histogram is TREM-2A-expressing cells and the open histogram is TREM-3-expressing cells. The data are representative of three independent experiments.
TREM-2A reporter assay analyses
We used a BWZ reporter cell line assay to establish whether the binding of gonococci or LOS to TREM-2A on the surface of cells could initiate a transmembrane signaling event. Cells were incubated with gonococci at a bacteria to cell ratio of either 1:1 or 100:1, and for LOS, cells were treated with concentrations of either 1 μg/ml or 100 μg/ml. We found that engagement of TREM-2A by gonococci as well as by LOS induced transmembrane signaling as evidenced by a twofold or greater increase in lacZ activity in TREM-2A-expressing cells compared with nontransfected cells (Figure 6; p < 0.05 for all comparisons). In contrast, BWZ reporter cells expressing TREM-3 showed significantly less lacZ activity than TREM-2A-expressing cells when treated with strains 1291 or F62 gonococci (p < 0.001).
Figure 6.

Gonococci and purified LOS engage membrane-expressed TREM-2A and induce transmembrane signaling. BWZ reporter cell constructs expressing either TREM-2A (TR-2A) or TREM-3 (TR-3) were used to measure LacZ activity as an indicator of signal transduction following incubation of cells with gonococci or LOS. For stimulation with gonococci, reporter cells were incubated with a bacteria to cell ratio of either 100:1 or 1:1. For LOS, TREM-2A-expressing reporter cells were treated with LOS concentrations of either 100 μg/ml or 1 μg/ml. The LOS and all strains of gonococci induced LacZ activity but to variable degrees. Significantly greater signaling was induced in cells expressing TREM-2A than in those expressing TREM-3. Statistical comparisons: p < 0.05 for all comparisons of lacZ activity induced by gonococci and LOS in TREM-2A-expressing cells versus nontransfected cells; p < 0.001 for the comparisons of lacZ activity induced by 1291 and F62 in TREM-2A-expressing cells versus TREM-3-expressing cells. The bars represent the mean of triplicate data points and the error bars are ±SD. The results are representative of four independent experiments.
TREM-2 expression by human reproductive tract epithelial cells
We next determined whether human TREM-2 is constitutively expressed by reproductive tract epithelial cells that are known to be invaded by N. gonorrhoeae (12, 38, 39). Constitutive expression of TREM-2 by cultured human ectocervical, endocervical, vaginal, ME-180, and HeLa cells was detected by flow cytometry (Figure 7). As a positive control, THP-1 cells were found to express TREM-2 as reported previously (40). Immunohistochemical staining also showed constitutive expression of TREM-2 by fallopian tubes as seen in the photomicrographs in Figure 8. Expression of TREM-2 was patchy in distribution and was localized to the epithelium with scant cells staining for the protein in the stroma. Greater expression was observed on the apical side of the epithelial cells and expression was detected predominantly on ciliated cells.
Figure 7.
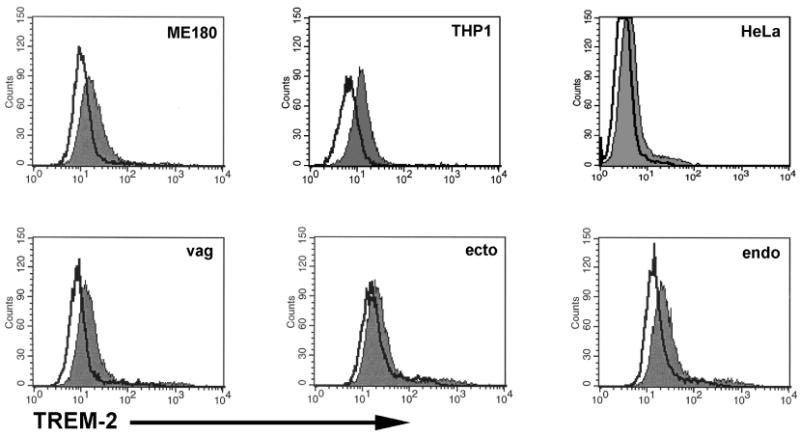
Human reproductive tract epithelial cells constitutively express TREM-2. Expression of TREM-2 by cultured human ectocervical, endocervical, vaginal, ME-180, and HeLa cells was detected by flow cytometric analyses. THP-1 cells known to express TREM-2 were stained as a positive control. Filled histograms represent cells stained with an anti-human TREM-2 antibody and open histograms represent cells stained with an irrelevant isotype matched antibody. The results are representative of four independent experiments.
Figure 8.
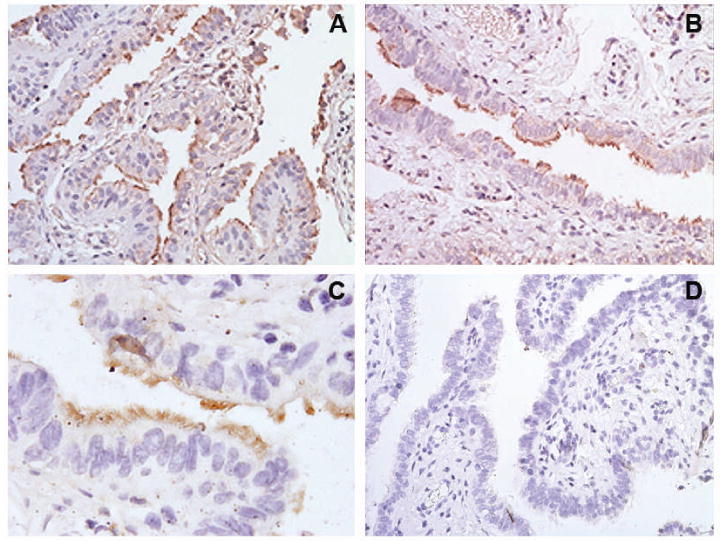
Human fallopian tube epithelial cells constitutively express TREM-2. Photomicrographs of human fallopian tube organ cultures stained immunohistochemically for the expression of TREM-2. Panels A-C: Sections from three different female subjects stained for TREM-2 expression. Each section shows the localization of TREM-2 in the apical side of the brown-stained epithelium lining the lumen with little stromal cell staining for the protein. Staining was patchy in distribution with groups of stained cells located adjacent to groups of unstained cells, and expression was detected predominantly on ciliated epithelial cells; original magnification, ×200 (panels A and B), ×600 (panel C). Panel D: Negative control section incubated with non-immune goat IgG and the secondary detection antibody. No staining of any cell type was observed in control sections; original magnification, ×200.
IL-6 induction by HeLa cells through LOS ligation of TREM-2
To assess the biological importance of TREM-2 ligation on reproductive tract epithelial cells, we investigated whether the engagement of TREM-2 by LOS resulted in the production of proinflammatory cytokine IL-6. HeLa cells were chosen as the test cell line as they do not express MD2, which is necessary for the response of TLR4 to LOS (7). Thus, induction of IL-6 in response to LOS treatment would be independent of signaling through the TLR4 receptor complex. When treated with strain 1291 LOS for 12 hours, HeLa cells expressed a significant dose-dependent increase in IL-6 compared with the constitutive level of expression by untreated cells (Figure 9). To confirm the role of TREM-2 ligation in the stimulation of IL-6 production in response to LOS, cells were treated with affinity-purified goat anti-human TREM-2 polyclonal IgG prior to exposure of the cells to LOS to block the interaction between TREM-2 and LOS. As shown in Figure 9, IL-6 production by HeLa cells was significantly reduced as the result of treatment of the cells with anti-TREM-2 antibody (p < 0.05 for both LOS concentrations comparing antibody treated versus untreated). This suggests that IL-6 production by HeLa cells was triggered as a functional consequence of TREM-2 ligation by LOS.
Figure 9.

Ligation of TREM-2 by LOS induces IL-6 production in HeLa cells. HeLa cells were treated with LOS from strain 1291 at concentrations of 10 and 100 ng/ml for 12 hours. Control wells were treated with culture media only. For some experiments, cells were treated with affinity-purified goat anti-human TREM-2 polyclonal IgG prior to addition of the LOS. Supernatants from the LOS-treated and untreated cells in the presence and absence of anti-human TREM-2 were assayed for IL-6 by ELISA. The bars represent the mean of quadruplicate data points and the error bars are ±SEM. The results are representative of three independent experiments. Statistical comparisons: *, p > 0.1; **, p < 0.05; ***, p < 0.05.
Discussion
Evidence is growing to support the concept that the interaction of gonococci with innate immune receptors plays a critical role in the initial response to gonococcal infection. The results of this study demonstrate that gonococci can interact with the TREM-2 member of the TREM family of receptors and initiate transmembrane cell signaling and cytokine production. In contrast, a related member of the TREM family, TREM-3, showed minimal interactions with gonococci. Both soluble and cell surface-expressed TREM-2A bound to N. gonorrhoeae through interactions with LOS and Opa proteins with the predominant binding to LOS. Binding of TREM-2A to whole bacteria paralleled its binding to LOS purified from them, suggesting that binding to LOS accounts for the majority of recognition of these bacteria by TREM-2A. Surface plasmon resonance analyses revealed that the affinity of TREM-2A for the highest binding 1291 LOS was comparable to that of E. coli LPS for LPS-binding protein (41) and for the LPS-scavenging high-density lipoprotein (42).
The ligands for TREM-2 are not well-characterized. A study by Dawes et al. identified ligands for murine TREM-2 on both bacteria and mammalian cells and concluded that TREM-2 recognizes anionic ligands such as E. coli LPS (25). The results of our study extend those findings and provide a more detailed analysis of the structural features of the anionic gonococcal LOS molecule important for recognition by TREM-2. We found that lipid A acylation and not OS α chain structure is the critical determinant of TREM-2A binding to LOS. This was evidenced by the finding that O-deacylation of LOS significantly reduced the binding of TREM-2A to the three strains tested, indicating that the lipoidal moiety of the LOS dictates TREM-2A binding. Heterogeneity in the structure of gonococcal lipid A in terms of differences in the number of acyl groups (hexaacyl versus pentaacyl) as well as in the numbers of phosphate and phosphoethanolamine residues contained within the lipid A molecules expressed by each strain likely underlie the variability in the binding of TREM-2A to LOS (15, 17). In contrast, we observed a significant difference in the binding of TREM-2A to 1291 and F62 LOS, both of which express a structurally identical OS α chain terminating in lacto-N-neotetraose (27). If TREM-2A binding was dependent on the OS structure, one would anticipate similar levels of binding of TREM-2A to the LOS from these two strains, which was not observed. In addition, the binding of TREM-2A to MkC and to mutational variant MkC Δ1-5 expressing a truncated OS was similar, lending further support to the conclusion that OS structure does not play a significant role in TREM-2A binding. This result is analogous to our previous finding that structural differences in the OS α chain did not affect cell signaling mediated by TLR4 (7), indicating that both types of pattern recognition receptors target the lipid A moiety of LOS.
Recently, a putative soluble variant of human TREM-2 lacking a transmembrane domain was reported (43). This protein could act as a soluble receptor for LOS and Opa on bacterial membranes if produced by cells in the local environment of gonococcal infection. The effect of soluble TREM-2 binding to these molecules remains to be investigated, but we can speculate that it might modulate their interaction with human cells as both LOS and Opa function as adhesins for several epithelial cell receptors (7, 44-47). In addition, the binding of soluble TREM-2 to LOS and Opa may influence the binding and function of soluble proteins galectin-3 and mannose-binding lectin known to target LOS and Opa, respectively (27, 32).
In our examination of TREM-2 expression by cell types relevant to the pathogenesis of gonorrhea, we found that human TREM-2 is constitutively expressed by genitourinary epithelial cells and by the fallopian tube epithelium, both of which are primary targets of gonococcal invasion (12, 38, 39). To our knowledge, this is the first report of the expression of human TREM-2 by cells deriving from a non-myeloid lineage. Previously, cells of myeloid origin such as dendritic cells were the only cell types to express TREM-2 that were known to interact with gonococci (48, 49). For the immortalized cell lines, however, it should be noted that they contain DNA sequences from human papillomaviruses which could theoretically influence TREM-2 expression (50, 51), and as such they may not accurately represent the expression of TREM-2 on primary reproductive tract epithelial cells lacking papillomavirus DNA.
In the fallopian tube epithelium, expression of TREM-2 was patchy in distribution and was detected predominantly on ciliated cells. This expression pattern contrasts with that reported for the LOS-binding lectin galectin-3 which is limited in expression to nonciliated epithelial cells of the fallopian tube (27). The significance of a heterogeneous distribution in the constitutive expression of TREM-2 in the fallopian tube of an uninfected female is yet to be determined. As only a subset of epithelial cells in the fallopian tube expressed TREM-2, the data suggest that regional differences in TREM-2 expression may influence local responses to gonococcal infection.
We used a BWZ reporter cell line assay to determine whether the binding of gonococci to TREM-2A on the surface of cells could initiate a transmembrane signaling event. The assay utilized BWZ cells containing a lacZ reporter gene which were transfected to surface-express TREM-2 linked to a CD3ζ cytoplasmic domain. Signaling through CD3ζ activated NFAT promoter elements which induced expression of the lacZ reporter gene. We found that engagement of TREM-2A by whole bacteria and LOS induced transmembrane signaling through the CD3ζ cytoplasmic domain resulting in lacZ gene activity. In contrast, cells transfected to surface-express TREM-3 showed minimal lacZ gene activity. Although we did not determine directly whether the binding of bacteria or LOS to TREM-2A on reporter cells signaled through DAP12, we did obtain evidence that the ligation by LOS of human TREM-2 expressed on HeLa cells resulted in the production of proinflammatory cytokine IL-6. This suggests that the ligation of TREM-2 by gonococci mediates signal transduction through a functional molecular complex of DAP12 and TREM-2 which may play a role in the innate immune response of human reproductive tract epithelial cells to gonococcal infection. However, unlike TREM-1 which amplifies the inflammatory response to bacteria through the production of proinflammatory cytokines and chemokines when engaged (21, 24), ligation of TREM-2 results in upregulation of certain receptors such as CCR7, CD40, and CD86 on dendritic cells (48), but inhibition of TLR receptor responses such as TNF production in macrophages (52). A similar inhibitory response mediated by TREM-2 expressed by myeloid cell types in the human genitourinary tract may contribute to the limited cytokine responses seen in uncomplicated cervical gonococcal infection (53). Further characterization of the importance of the DAP12/TREM-2-mediated signal transduction pathway in both myeloid and non-myeloid cell types in response to gonococcal infection will provide new insights into the pathogenesis of gonorrhea.
In conclusion, this study provides evidence that TREM-2 can function as a pattern recognition receptor in the induction of the innate immune response to gonococci. Because of its constitutive expression on epithelial cells that are the target of gonococcal invasion, and its affinity for gonococcal outer membrane components LOS and Opa required for efficient invasion, TREM-2 is likely to play some role in the pathogenesis of gonococcal infection.
Acknowledgments
We thank Dr. William Seaman of the VA Medical Center in San Francisco for supplying soluble TREM-2A/IgG1-Fc and TREM-3/IgG1-Fc fusion proteins, as well as TREM-2A- and TREM-3-expressing BWZ cells. We thank Ona Barauskas for her excellent technical assistance. This work was supported by Public Health Service grant AI063927 (GAJ) from the National Institute of Allergy and Infectious Diseases and by the Research Service of the U.S. Department of Veterans Affairs (GAJ). This is paper number 101 from the Center for Immunochemistry.
References
- 1.Centers for Disease Control and Prevention. Sexually Transmitted Disease Surveillance. U S Department of Health and Human Services, Public Health Service. 2005 [Google Scholar]
- 2.Paavonen J. Pelvic inflammatory disease. From diagnosis to prevention. Dermatol Clin. 1998;16:747–756. doi: 10.1016/s0733-8635(05)70041-3. [DOI] [PubMed] [Google Scholar]
- 3.Cohen MS, et al. Reduction of concentration of HIV-1 in semen after treatment of urethritis: implications for prevention of sexual transmission of HIV-1. Lancet. 1997;349:1868–1873. doi: 10.1016/s0140-6736(97)02190-9. [DOI] [PubMed] [Google Scholar]
- 4.Chesson HW, Pinkerton SD. Sexually transmitted diseases and the increased risk for HIV transmission: implications for cost-effectiveness analyses of sexually transmitted disease prevention interventions. J Acquir Immune Defic Syndr. 2000;24:48–56. doi: 10.1097/00126334-200005010-00009. [DOI] [PubMed] [Google Scholar]
- 5.Mabey D. Interactions between HIV infection and other sexually transmitted diseases. Trop Med Int Health. 2000;5:A32–A36. doi: 10.1046/j.1365-3156.2000.00595.x. [DOI] [PubMed] [Google Scholar]
- 6.Farley TA, Cohen DA, Wu SY, Besch CL. The value of screening for sexually transmitted diseases in an HIV clinic. J Acquir Immune Defic Syndr. 2003;33:642–648. doi: 10.1097/00126334-200308150-00014. [DOI] [PubMed] [Google Scholar]
- 7.Pridmore AC, et al. Activation of toll-like receptor 2 (TLR2) and TLR4/MD2 by Neisseria is independent of capsule and lipooligosaccharide (LOS) sialylation but varies widely among LOS from different strains. Infect Immun. 2003;71:3901–3908. doi: 10.1128/IAI.71.7.3901-3908.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Massari P, et al. Cutting edge: Immune stimulation by neisserial porins is toll-like receptor 2 and MyD88 dependent. J Immunol. 2002;168:1533–1537. doi: 10.4049/jimmunol.168.4.1533. [DOI] [PubMed] [Google Scholar]
- 9.Fisette PL, Ram S, Andersen JM, Guo W, Ingalls RR. The Lip lipoprotein from Neisseria gonorrhoeae stimulates cytokine release and NF-κB activation in epithelial cells in a Toll-like receptor 2-dependent manner. J Biol Chem. 2003;278:46252–46260. doi: 10.1074/jbc.M306587200. [DOI] [PubMed] [Google Scholar]
- 10.Fichorova RN, Desai PJ, Gibson FC, III, Genco CA. Distinct proinflammatory host responses to Neisseria gonorrhoeae infection in immortalized human cervical and vaginal epithelial cells. Infect Immun. 2001;69:5840–5848. doi: 10.1128/IAI.69.9.5840-5848.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Naumann M, Wessler S, Bartsch C, Wieland B, Meyer TF. Neisseria gonorrhoeae epithelial cell interaction leads to the activation of the transcription factors nuclear factor κB and activator protein 1 and the induction of inflammatory cytokines. J Exp Med. 1997;186:247–258. doi: 10.1084/jem.186.2.247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Jarvis GA, Li J, Swanson KV. Invasion of human mucosal epithelial cells by Neisseria gonorrhoeae upregulates expression of intercellular adhesion molecule 1 (ICAM-1) Infect Immun. 1999;67:1149–1156. doi: 10.1128/iai.67.3.1149-1156.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ramsey KH, et al. Inflammatory cytokines produced in response to experimental human gonorrhea. J Infect Dis. 1995;172:186–191. doi: 10.1093/infdis/172.1.186. [DOI] [PubMed] [Google Scholar]
- 14.McGee ZA, et al. Gonococcal infection of human fallopian tube mucosa in organ culture: relationship of mucosal tissue TNF-α concentration to sloughing of ciliated cells. Sex Transm Dis. 1999;26:160–165. doi: 10.1097/00007435-199903000-00007. [DOI] [PubMed] [Google Scholar]
- 15.Kahler CM, Stephens DS. Genetic basis for biosynthesis, structure, and function of meningococcal lipooligosaccharide (endotoxin) Crit Rev Microbiol. 1998;24:281–334. doi: 10.1080/10408419891294216. [DOI] [PubMed] [Google Scholar]
- 16.Griffiss JM, et al. Physical heterogeneity of neisserial lipooligosaccharides reflects oligosaccharides that differ in apparent molecular weight, chemical composition, and antigenic expression. Infect Immun. 1987;55:1792–1800. doi: 10.1128/iai.55.8.1792-1800.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Takayama K, et al. Characterization of a structural series of lipid A obtained from the lipopolysaccharides of Neisseria gonorrhoeae. Combined laser desorption and fast atom bombardment mass spectral analysis of high performance liquid chromatography-purified dimethyl derivatives. J Biol Chem. 1986;261:10624–10631. [PubMed] [Google Scholar]
- 18.Erwin AL, Mandrell RE, Munford RS. Enzymatically deacylated Neisseria lipopolysaccharide (LPS) inhibits murine splenocyte mitogenesis induced by LPS. Infect Immun. 1991;59:1881–1887. doi: 10.1128/iai.59.6.1881-1887.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Roth RI, Yamasaki R, Mandrell RE, Griffiss JM. Ability of gonococcal and meningococcal lipooligosaccharides to clot Limulus amebocyte lysate. Infect Immun. 1992;60:762–767. doi: 10.1128/iai.60.3.762-767.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Colonna M. TREMs in the immune system and beyond. Nat Rev Immunol. 2003;3:445–453. doi: 10.1038/nri1106. [DOI] [PubMed] [Google Scholar]
- 21.Bouchon A, Dietrich J, Colonna M. Cutting edge: inflammatory responses can be triggered by TREM-1, a novel receptor expressed on neutrophils and monocytes. J Immunol. 2000;164:4991–4995. doi: 10.4049/jimmunol.164.10.4991. [DOI] [PubMed] [Google Scholar]
- 22.Bouchon A, Hernandez-Munain C, Cella M, Colonna M. A DAP12-mediated pathway regulates expression of CC chemokine receptor 7 and maturation of human dendritic cells. J Exp Med. 2001;194:1111–1122. doi: 10.1084/jem.194.8.1111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Daws MR, Lanier LL, Seaman WE, Ryan JC. Cloning and characterization of a novel mouse myeloid DAP12-associated receptor family. Eur J Immunol. 2001;31:783–791. doi: 10.1002/1521-4141(200103)31:3<783::aid-immu783>3.0.co;2-u. [DOI] [PubMed] [Google Scholar]
- 24.Bouchon A, Facchetti F, Weigand MA, Colonna M. TREM-1 amplifies inflammation and is a crucial mediator of septic shock. Nature. 2001;410:1103–1107. doi: 10.1038/35074114. [DOI] [PubMed] [Google Scholar]
- 25.Daws MR, et al. Pattern recognition by TREM-2: binding of anionic ligands. J Immunol. 2003;171:594–599. doi: 10.4049/jimmunol.171.2.594. [DOI] [PubMed] [Google Scholar]
- 26.Swanson KV, Griffiss JM. Separation and identification of neisserial lipooligosaccharide oligosaccharides using high-performance anion-exchange chromatography with pulsed amperometric detection. Carbohydr Res. 2006;341:388–396. doi: 10.1016/j.carres.2005.11.017. [DOI] [PubMed] [Google Scholar]
- 27.John CM, et al. Galectin-3 binds lactosaminylated lipooligosaccharides from Neisseria gonorrhoeae and is selectively expressed by mucosal epithelial cells that are infected. Cell Microbiol. 2002;4:649–662. doi: 10.1046/j.1462-5822.2002.00219.x. [DOI] [PubMed] [Google Scholar]
- 28.Zollinger WD, Mandrell RE, Griffiss JM, Altieri P, Berman S. Complex of meningococcal group B polysaccharide and type 2 outer membrane protein immunogenic in man. J Clin Invest. 1979;63:836–848. doi: 10.1172/JCI109383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Apicella MA, Griffiss JM, Schneider H. Isolation and characterization of lipopolysaccharides, lipooligosaccharides, and lipid A. Methods Enzymol. 1994;235:242–52. 242–252. doi: 10.1016/0076-6879(94)35145-7. [DOI] [PubMed] [Google Scholar]
- 30.John CM, Schneider H, Griffiss JM. Neisseria gonorrhoeae that infect men have lipooligosaccharides with terminal N-acetyllactosamine repeats. J Biol Chem. 1999;274:1017–1025. doi: 10.1074/jbc.274.2.1017. [DOI] [PubMed] [Google Scholar]
- 31.Griffiss JM, Lammel CJ, Wang J, Dekker NP, Brooks GF. Neisseria gonorrhoeae coordinately uses Pili and Opa to activate HEC-1-B cell microvilli, which causes engulfment of the gonococci. Infect Immun. 1999;67:3469–3480. doi: 10.1128/iai.67.7.3469-3480.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Estabrook MM, Jack DL, Klein NJ, Jarvis GA. Mannose-binding lectin binds to two major outer membrane proteins, opacity protein and porin, of Neisseria meningitidis. J Immunol. 2004;172:3784–3792. doi: 10.4049/jimmunol.172.6.3784. [DOI] [PubMed] [Google Scholar]
- 33.Estabrook MM, Griffiss JM, Jarvis GA. Sialylation of Neisseria meningitidis lipooligosaccharide inhibits serum bactericidal activity by masking lacto-N-neotetraose. Infect Immun. 1997;65:4436–4444. doi: 10.1128/iai.65.11.4436-4444.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Takamatsu D, et al. Binding of the Streptococcus gordonii surface glycoproteins GspB and Hsa to specific carbohydrate structures on platelet membrane glycoprotein Ibα. Mol Microbiol. 2005;58:380–392. doi: 10.1111/j.1365-2958.2005.04830.x. [DOI] [PubMed] [Google Scholar]
- 35.Sanderson S, Shastri N. LacZ inducible, antigen/MHC-specific T cell hybrids Int Immunol. 1994;6:369–376. doi: 10.1093/intimm/6.3.369. [DOI] [PubMed] [Google Scholar]
- 36.Cooper MD, Rapp J, Jeffery-Wiseman C, Barnes RC, Stephens DS. Chlamydia trachomatis infection of human fallopian tube organ cultures. J Gen Microbiol. 1990;136:1109–1115. doi: 10.1099/00221287-136-6-1109. [DOI] [PubMed] [Google Scholar]
- 37.Chung DH, Seaman WE, Daws MR. Characterization of TREM-3, an activating receptor on mouse macrophages: definition of a family of single Ig domain receptors on mouse chromosome 17. Eur J Immunol. 2002;32:59–66. doi: 10.1002/1521-4141(200201)32:1<59::AID-IMMU59>3.0.CO;2-U. [DOI] [PubMed] [Google Scholar]
- 38.Fichorova RN, Cronin AO, Lien E, Anderson DJ, Ingalls RR. Response to Neisseria gonorrhoeae by cervicovaginal epithelial cells occurs in the absence of toll-like receptor 4-mediated signaling. J Immunol. 2002;168:2424–2432. doi: 10.4049/jimmunol.168.5.2424. [DOI] [PubMed] [Google Scholar]
- 39.Cooper MD, McGee ZA, Mulks MH, Koomey JM, Hindman TL. Attachment to and invasion of human fallopian tube mucosa by an IgA1 protease-deficient mutant of Neisseria gonorrhoeae and its wild-type parent. J Infect Dis. 1984;150:737–744. doi: 10.1093/infdis/150.5.737. [DOI] [PubMed] [Google Scholar]
- 40.Allcock RJ, Barrow AD, Forbes S, Beck S, Trowsdale J. The human TREM gene cluster at 6p21.1 encodes both activating and inhibitory single IgV domain receptors and includes NKp44. Eur J Immunol. 2003;33:567–577. doi: 10.1002/immu.200310033. [DOI] [PubMed] [Google Scholar]
- 41.Thomas CJ, et al. Evidence of a trimolecular complex involving LPS, LPS binding protein and soluble CD14 as an effector of LPS response. FEBS Lett. 2002;531:184–188. doi: 10.1016/s0014-5793(02)03499-3. [DOI] [PubMed] [Google Scholar]
- 42.Levels JH, et al. Lipopolysaccharide is transferred from high-density to low-density lipoproteins by lipopolysaccharide-binding protein and phospholipid transfer protein. Infect Immun. 2005;73:2321–2326. doi: 10.1128/IAI.73.4.2321-2326.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Begum NA, et al. Mycobacterium bovis BCG cell wall-specific differentially expressed genes identified by differential display and cDNA subtraction in human macrophages. Infect Immun. 2004;72:937–948. doi: 10.1128/IAI.72.2.937-948.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Zhang P, et al. DC-SIGN (CD209) recognition of Neisseria gonorrhoeae is circumvented by lipooligosaccharide variation. J Leukoc Biol. 2006;79:731–738. doi: 10.1189/jlb.0405184. [DOI] [PubMed] [Google Scholar]
- 45.Harvey HA, Jennings MP, Campbell CA, Williams R, Apicella MA. Receptor-mediated endocytosis of Neisseria gonorrhoeae into primary human urethral epithelial cells: the role of the asialoglycoprotein receptor. Mol Microbiol. 2001;42:659–672. doi: 10.1046/j.1365-2958.2001.02666.x. [DOI] [PubMed] [Google Scholar]
- 46.Porat N, Apicella MA, Blake MS. A lipooligosaccharide-binding site on HepG2 cells similar to the gonococcal opacity-associated surface protein Opa. Infect Immun. 1995;63:2164–2172. doi: 10.1128/iai.63.6.2164-2172.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Virji M, Watt SM, Barker S, Makepeace K, Doyonnas R. The N-domain of the human CD66a adhesion molecule is a target for Opa proteins of Neisseria meningitidis and Neisseria gonorrhoeae. Mol Microbiol. 1996;22:929–939. doi: 10.1046/j.1365-2958.1996.01548.x. [DOI] [PubMed] [Google Scholar]
- 48.Bouchon A, Hernandez-Munain C, Cella M, Colonna M. A DAP12-mediated pathway regulates expression of CC chemokine receptor 7 and maturation of human dendritic cells. J Exp Med. 2001;194:1111–1122. doi: 10.1084/jem.194.8.1111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Zhang J, et al. Neisseria gonorrhoeae enhances infection of dendritic cells by HIV type 1. J Immunol. 2005;174:7995–8002. doi: 10.4049/jimmunol.174.12.7995. [DOI] [PubMed] [Google Scholar]
- 50.Pater MM, Pater A. Human papillomavirus types 16 and 18 sequences in carcinoma cell lines of the cervix. Virology. 1985;145:313–318. doi: 10.1016/0042-6822(85)90164-3. [DOI] [PubMed] [Google Scholar]
- 51.Fichorova RN, Rheinwald JG, Anderson DJ. Generation of papillomavirus-immortalized cell lines from normal human ectocervical, endocervical, and vaginal epithelium that maintain expression of tissue-specific differentiation proteins. Biol Reprod. 1997;57:847–855. doi: 10.1095/biolreprod57.4.847. [DOI] [PubMed] [Google Scholar]
- 52.Hamerman JA, et al. Cutting edge: Inhibition of TLR and FcR responses in macrophages by triggering receptor expressed on myeloid cells (TREM)-2 and DAP12. J Immunol. 2006;177:2051–2055. doi: 10.4049/jimmunol.177.4.2051. [DOI] [PubMed] [Google Scholar]
- 53.Hedges SR, Sibley DA, Mayo MS, Hook EW, III, Russell MW. Cytokine and antibody responses in women infected with Neisseria gonorrhoeae: effects of concomitant infections. J Infect Dis. 1998;178:742–751. doi: 10.1086/515372. [DOI] [PubMed] [Google Scholar]