

Abstract
Dark-adapted rods suppress cone-mediated flicker detection. This study evaluates the effect that rod activity has on cone temporal processing by investigating whether rod mediated suppression changes the cone pathway impulse response function, regardless of the form of the temporal signal. Stimuli were generated with a 2-channel photostimulator that has four primaries for the central field and four primaries for the surround. Cone pathway temporal impulse response functions were derived from temporal contrast sensitivity data with periodic stimuli, and from two-pulse discrimination data in which pairs of briefly pulsed stimuli were presented successively at a series of stimulus onset asynchronies. Dark-adapted rods altered the amplitude and timing of cone pathway temporal impulse response functions, irrespective of whether they were derived from measurements with temporally periodic stimuli or in a brief presentation temporal resolution task with pulsed stimuli. Rod-cone interactions are a fundamental operation in visual temporal processing under mesopic light levels, acting to decrease the temporal bandwidth of the visual system.
Keywords: rod-cone interaction, impulse response, temporal contrast sensitivity, two-pulse discrimination, mesopic
1. Introduction
There are a substantial range of mesopic lighting conditions, spanning approximately four log units in natural viewing environments (CIE, 1978), where interactions between the rod and cone photoreceptor signals alter visual function. Different forms of rod-cone interactions exist (for a review see Buck, 2004). Here we concentrate on lateral suppressive rod-cone interactions in temporal processing: the suppression of cone periodic flicker sensitivity by dark-adapted rods located in the region surrounding the stimulus area (e.g. Alexander & Fishman, 1984; Cao, Zele & Pokorny, 2006; Coletta & Adams, 1984; Goldberg, Frumkes & Nygaard, 1983; Lange, Denny & Frumkes, 1997; Lythgoe & Tansley, 1929; Zele & Vingrys, 2007). We considered whether the state of rod adaptation surrounding the cone-detected stimulus solely altered sensitivity to periodic (flickering) stimuli, or if the lateral rod suppression is a more general visual phenomenon that also alters cone sensitivity to double pulsed stimuli.
The locus of the rod-cone interaction is unclear. In amphibians it is hypothesized to occur at the synapse between cones and horizontal cells, ostensibly due to an inhibitory feedback signal from rods to cones (Frumkes & Eysteinsson, 1988). Horizontal cells in primates however, are additive and synapse primarily with cones (Dacey, Lee, Stafford, Pokorny & Smith, 1996). Early reports suggested that rod-cone interactions were L-cone specific (Coletta & Adams, 1985; Frumkes, 1990; Frumkes, Naarendorp & Goldberg, 1988). More recently, using experimental conditions that control the adaptation levels of rods and cones at the same chromaticity and rod excitation level, Cao, Zele and Pokorny (2006) demonstrated that dark-adapted rods suppress both L- and M-cone mediated flicker detection. Chromatic flicker detection was largely unaffected, implicating the Magnocellular (MC) pathway as a possible site of suppressive rod-cone interaction in humans.
Past investigations of rod-cone interactions in temporal processing typically varied stimulus wavelength and/or illumination level as a means of altering rod and cone excitation (e.g. Coletta & Adams, 1984; Goldberg et al., 1983; Naarendorp & Frumkes, 1991; Temme & Frumkes, 1977). We used instrumentation that allowed independent control of rod and cone excitation at the same time-averaged chromaticity and illumination level (also see Cao et al., 2006; Cao, Zele & Pokorny, 2008; Pokorny, Smithson & Quinlan, 2004; Sun, Pokorny & Smith, 2001). To evaluate the effect that the state of rod adaptation had on cone sensitivity to periodic and pulsed visual stimuli, we compared cone pathway impulse response functions (IRFs) derived by two independent measurements, one for temporal contrast sensitivity (de Lange, 1958) and the other for two-pulse discrimination (Burr & Morrone, 1993; Ikeda, 1986). Parafoveal measurements were made for a stimulus field surrounded by darkness, following dark adaptation or during the cone plateau after termination of a partial rod bleach. Rather than the addition of a surround, a cone plateau condition was chosen to render the derived IRFs most comparable to those measured following dark adaptation. Surrounds can significantly alter low-frequency temporal contrast sensitivity (Keesey, 1970; Kelly, 1969), but not high frequency temporal contrast sensitivity. Cao et al., (2006) showed that, in comparison with the critical flicker-fusion frequency (CFF) measured for an isolated field following dark adaptation, the improvement in CFF during the cone plateau was the same as that measured in the presence of an equiluminant surround. Here, a control experiment showed the addition of an equiluminant surround following dark adaptation resulted in similar temporal contrast sensitivity and two-pulse summation functions as measured during the cone plateau.
2. Methods
2.1. Apparatus and calibration procedures
A 2-channel, 4-primary Maxwellian view photostimulator (Pokorny et al., 2004) provided independent control of the stimulations of the rods and three types of cones in the human retina (Shapiro, Pokorny and Smith, 1996). A 2° circular field was viewed at a 7.5° eccentricity in the temporal retina. For all conditions reported in this study, the cone chromaticities were metameric to the equal-energy-spectrum light [L/(L+M) = 0.667, S/(L+M) =1.0 in a relative cone Troland chromaticity space; Smith and Pokorny (1996)]. Examples of the implementation of the photostimulator are detailed in Cao, Zele and Pokorny (2007) and Zele, Cao and Pokorny (2007).
The photostimulator primaries are derived from light-emitting diode (LED)-interference filter combinations yielding dominant wavelengths of 459 nm (blue), 516 nm (green), 561 nm (greenish yellow) and 658 nm (red). The radiances of the primaries are controlled by amplitude modulation of a 20 kHz carrier feeding into an eight-channel analog output Dolby sound card (M-Audio-Revolution 7.1 PCI) with a 24-bit digital-to-analog converter (DAC) operating at a sampling rate of 192 kHz. The output of each DAC was demodulated (Puts, Pokorny, Quinlan & Glennie, 2005) and sent to a voltage to frequency converter that provided 1-μs pulses at frequencies up to 250 kHz to control the LEDs (Swanson, Ueno, Smith & Pokorny, 1987). The sound card with demodulator has a precision of greater than 16 bits (Puts et al., 2005). All stimuli were generated using custom engineered software driven by a Macintosh G5 PowerPC computer.
Observer calibration procedures were conducted at the same peripheral retinal location of the stimulus field as for the experiments, to compensate for individual differences in prereceptoral filtering and receptoral spectral sensitivities between the observer and the CIE 1964 10° standard observer. Details of the physical light calibrations and the observer calibration procedures are described elsewhere (Cao et al., 2007; Cao, Pokorny & Smith, 2005; Pokorny et al., 2004; Sun et al., 2001).
2.2. Psychophysical paradigms
We measured temporal contrast sensitivity and two-pulse discrimination. In the temporal contrast sensitivity paradigm, the waveform periodically repeated at the flicker frequency for the duration of the temporal envelope (Figure 1A). In the two-pulse discrimination paradigm, pairs of stimulus pulses were presented in discrete intervals defined by the stimulus onset asynchrony (SOA) (Figure 1B and 1C). The central 2° stimulus field was 80 photopic Td and set in a dark surround for all experimental conditions (except for one of the control experiments). The stimulus field contained a cone luminance modulation at a fixed chromaticity (L+M+S), with constant rod excitation. Temporal contrast sensitivity was measured with periodic sinusoidal stimuli modulated within a 1 sec raised cosine envelope (constant time-averaged luminance) to minimize adaptation to the flickering stimuli. Because symmetric and rectified flicker stimuli can result in different threshold outcomes (Zele & Vingrys, 2007), the flicker was modulated symmetrically above and below the mean adaptation level (80 Td) of the 2° field, therefore effecting no change in the time-average retinal illuminance (Figure 1). Temporal frequencies ranged from 3 to 26 Hz. Discrete Fourier Transforms (4096-point DFT) showed the temporal bandwidths of all flickering stimuli to be less than ±1 Hz at half height of maximum amplitude. Two-pulse discrimination (Burr & Morrone, 1993; Ikeda, 1986) was measured with a pair of 4 ms rectangular pulses displayed successively at stimulus onset asynchronies (SOA) varying between 14 to 270 ms. The SOA was defined as the time (in ms) between pulse onsets. Measurement conditions included two incremental pulses (Figure 1B) or an incremental and a decremental pulse (Figure 1C).
Figure 1.
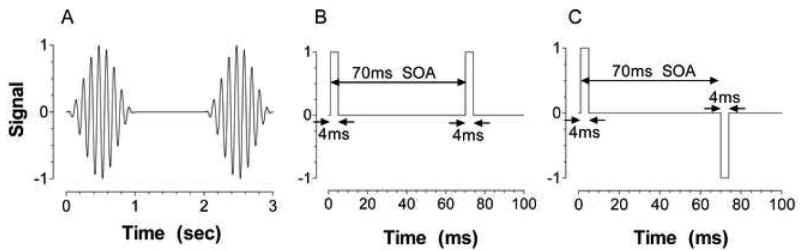
Temporal profiles of the stimuli used for the temporal contrast sensitivity (Panel A) and 2-pulse measurements (Panels B and C). Panel A gives an example of a 9 Hz periodic sinusoidal cone signal modulated within a 1 sec raised cosine envelope (constant time-averaged luminance) that alternated with a 1 sec steady field. Panel B shows a pair of 4 ms incremental pulses separated by a 70 ms stimulus onset asynchrony (SOA). The SOA was defined as the time (ms) between pulse onsets. Panel C shows an increment-decrement pulse combination separated by a 70 ms SOA.
The effect of rods in the area surrounding the cone-mediated stimulus was evaluated by comparing measurements from two pre-adaptation conditions: (1) following 30 minutes of dark adaptation (rods were fully sensitive, we call this the dark adaptation condition) and (2) during the first 4-5 minutes following the termination of 2 minutes exposure to a 10,000 Td broadband light. We call this the cone plateau condition. The light was a 24-V, 150-W tungsten halogen lamp that, in combination with a colour correcting filter (Lee 80A), produced a correlated colour temperature of 5000°K. The ratio of photopic to scotopic excitation for the light was 0.44. The 2 minutes exposure produced about a 12.5% rod bleach and would be expected to lead to a cone plateau duration of 4–5 min (Pugh, 1975; Wolf & Zigler, 1954). Cao, Zele and Pokorny (2006) provide further details of the instrumentation and calibration for the partial rod bleach protocol.
2.3. Control Experiments
We performed two control experiments. The first showed that the change in cone sensitivity was not due to light adaptation of the stimulus field in and of itself. For both the temporal contrast sensitivity and the 2-pulse paradigms, adding an equiluminant surround (13° in diameter) following dark adaptation resulted in similar temporal contrast sensitivity and two-pulse summation functions when compared to the data measured during the cone plateau. The second control experiment showed a negligible effect of scattered light from the central 2° stimulus field into the dark surround; following dark-adaptation, the critical fusion frequency (CFF) for a cone (L+M+S) signal was independent of the temporal phase of a dim surround (0.05 photopic Td) with (L+M+S+R) modulated at the same frequency as the central modulation.
2.4. Impulse Response Functions
Cone pathway impulse response functions were derived from the periodic and pulsed data using two independent techniques. For the temporal contrast sensitivity data, the IRFs were derived using a Kramers-Kronig relation to reconstruct the temporal phase spectrum with a minimum phase assumption (Stork & Falk, 1987). Scaling and extrapolations at the low and high frequencies were conducted according to procedures described by Swanson et al., (1987). For the two-pulse summation data, we estimated the IRF using the exponentially damped, frequency modulated sinusoid model without assuming a minimum phase (Burr & Morrone, 1993). Cao, Zele and Pokorny (2007) provide details of the procedures for deriving the IRFs, and discuss several caveats concerning the methodology.
2.5. Procedure
Observers binocularly dark-adapted for 30 min prior to the beginning of data collection. A chin rest maintained head position and refractive correction (if required) was inserted on the instrument side of the 2 mm artificial pupil. Observers used their right eye for all measurements. For the cone plateau condition, the observer viewed the 10,000 photopic Td adapting light for 2 min prior to the start of the test session. The time needed to complete a single condition was less than 5 min. Control experiments were conducted during separate sessions on separate days.
Trials were specified according to a two-yes-one-no double random alternating staircase procedure with a yes/no paradigm. One adaptation condition was evaluated during each session. All conditions were repeated a minimum of three times. For the temporal contrast sensitivity measurements, the observer reported seeing a flickering or steady field in each trial. For the two-pulse discrimination measurements, the observer reported seeing a single or double-pulsed stimulus. The paradigms included 10% blank trials. No change in the central 2° field was present during a blank trial. Both observers made less than 5% false positive responses. The staircase procedures updated the stimulus contrast and ended following 10 reversals at the criterion step size. The last six reversals were averaged as the measured threshold for that session.
2.6. Observers
Two experienced psychophysical observers, the authors AJZ and DC, participated. Both observers have normal colour vision (assessed by the Neitz OT anomaloscope) and hue discrimination (assessed by the Farnsworth-Munsell 100-Hue test). The Institutional Review Boards of the University of Chicago and Queensland University of Technology approved all experimental procedures and participants provided informed consent.
3. Results
Figure 2 shows the temporal contrast sensitivity data for the two observers (left and right panels). The average standard error of measurement (SEM) for the temporal contrast sensitivity was 1.65 for AJZ and 1.86 for DC. During the cone plateau, temporal contrast sensitivity was bandpass (open circles). After dark adaptation, temporal contrast sensitivity was attenuated (closed squares). The log contrast sensitivity difference between the two adaptation conditions showed that temporal frequencies greater than 6 - 8 Hz were attenuated following dark adaptation (inserts to Figure 1).
Figure 2.
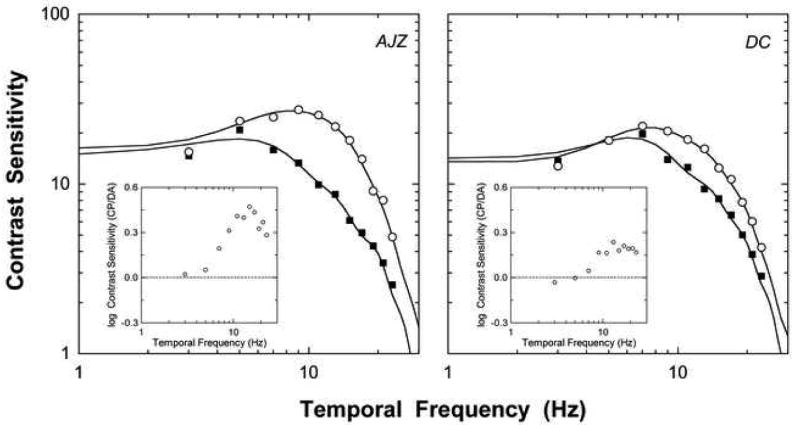
The effect of the state of rod adaptation on cone pathway sensitivity to periodic stimuli. Left and right panels show the data for two experienced psychophysical observers. Open circles show contrast sensitivity as a function of temporal frequency (Hz) for luminance modulated stimuli (L+M+S) during the cone plateau and the closed squares show contrast sensitivity after dark adaptation. The average standard error of measurement (SEM) was 1.65 for AJZ and 1.86 for DC. Panel inserts show the difference in log contrast sensitivity between the cone plateau (CP) and dark-adapted viewing conditions (CP/DA) for each observer. The solid lines show the Fourier transformation of the estimated IRF with the periodic stimulus (Fig 4, upper panels).
Figure 3 shows the two-pulse discrimination data for the two observers measured after dark adaptation (square symbols) and during the cone plateau (circular symbols). Open symbols show the data for the incremental pulses, and closed symbols for the incremental and decremental pulse combination. The two observers contrast sensitivity differed by about 0.2 log units. The average SEM was 0.67 for AJZ and 0.26 for DC. During the cone plateau, incremental pulse sensitivity was highest at the shortest interpulse interval and summation decreased with increasing stimulus asynchrony, reaching a minimum at ∼65 ms for AJZ and ∼70 ms for DC. For the incremental-decremental pulse combination, sensitivity was lowest at the shortest interpulse interval and improved with increasing interpulse interval to reach peak sensitivity at ∼45 ms for AJZ and ∼60 ms for DC. After dark adaptation, 2-pulse contrast sensitivity was reduced and the timing was altered. For the incremental pulses, maximum sensitivity occurred at ∼80 ms for AJZ and ∼65 ms for DC. For the incremental-decremental pulses, the minimum sensitivity occurred at ∼60 ms for AJZ and ∼65 ms for DC.
Figure 3.
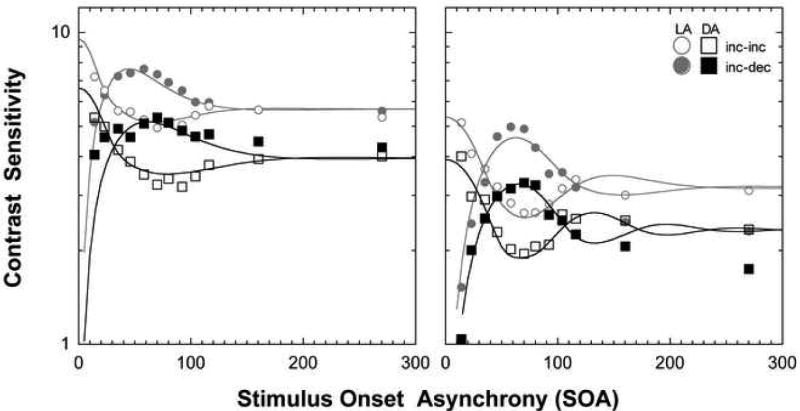
The effect of the state of rod adaptation on cone pathway sensitivity to pulsed stimuli. Left and right panels show the data for two experienced psychophysical observers. Circular symbols show the two-pulse discrimination data as a function of the stimulus onset asynchrony (SOA) of the two 4 ms pulses measured during the cone plateau, the square symbols after dark adaptation. Measurement conditions included two incremental pulses (open symbols) and one incremental and one decremental pulse (closed symbols). The average SEM was 0.67 for AJZ and 0.26 for DC. The solid lines show the predicted two pulse discrimination sensitivity based on the IRF with the pulsed stimuli (Fig 4, lower panels).
The cone pathway impulse response functions derived from the temporal contrast sensitivity and two-pulse discrimination data are shown in the upper and lower panels of Figure 4, respectively. The grey lines show the IRFs during the cone plateau, the black lines after dark adaptation. After dark-adaptation, the amplitude of the cone impulse response functions decreased and showed a delay in the time-to-peak relative to the cone plateau condition. For observer AJZ, the IRF derived from the periodic stimuli showed a 43% decrease in amplitude and a 9 ms delay in the time-to peak (upper left panel); the IRF derived from the pulsed stimuli showed a 34% decease in amplitude and 6 ms delay in the time-to peak (lower left panel). For observer DC, the IRF derived from the periodic stimuli showed a 21% decrease in amplitude and a 4 ms delay in the time-to peak (upper right panel); the IRF derived from the pulsed stimuli showed a 27% decrease in amplitude and a 9 ms delay in the time-to-peak (lower right panel).
Figure 4.
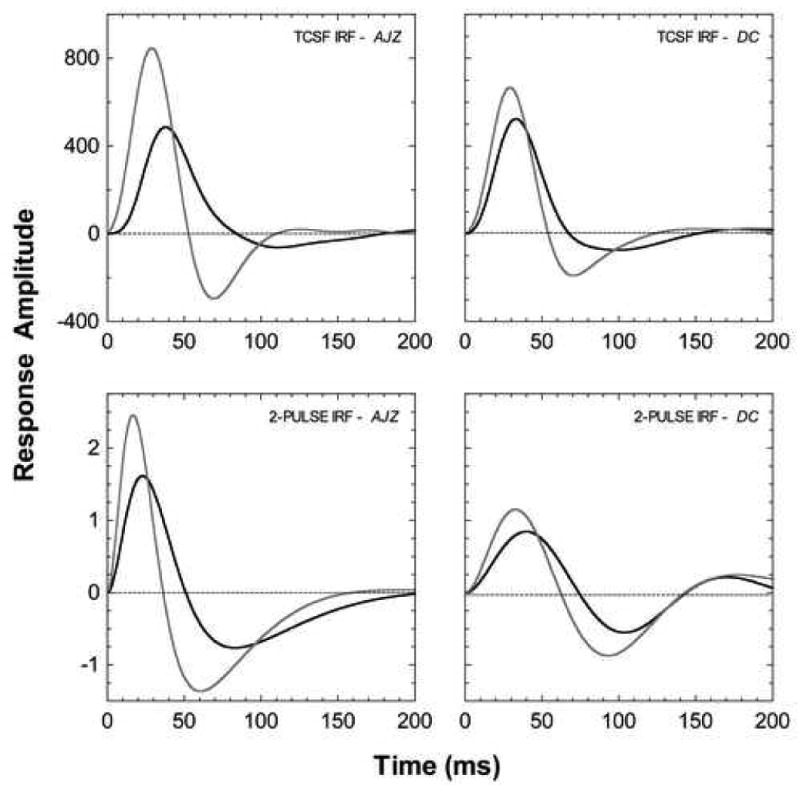
The effect of dark-adapted rods on the cone pathway temporal impulse response functions. Upper panels: Impulse response functions derived from the temporal contrast sensitivity data in Figure 2. Lower panels: Impulse response functions derived from the 2-pulse data in Figure 3. The grey lines show the IRFs during the cone plateau, the black lines show the IRFs after dark adaptation. The state of rod adaptation alters the amplitude and time time-to-peak of the cone pathway impulse response.
4. Discussion
This study was designed to test whether lateral suppressive rod-cone interactions were stimulus specific, that is, only occurred with the use of temporally periodic stimuli, or if the suppression was a more general visual phenomenon that could also alter cone sensitivity to double pulsed stimuli. To test this proposal, cone pathway contrast sensitivity was measured using periodic and double pulsed stimuli under viewing conditions that altered the level of rod activity in the area surrounding the stimulus. After dark-adaptation, temporal contrast sensitivity was attenuated at frequencies greater than 6 - 8 Hz, two-pulse contrast sensitivity decreased and the timing was altered. The mathematically derived IRFs demonstrate that after dark adaptation, the cone pathway IRF amplitude decreased and the time-to-peak was delayed.
Rod-cone interactions alter an observer's threshold for many types of visual stimuli (e.g. Alexander & Fishman, 1984; Kremers & Meierkord, 1999; MacLeod, 1972; Sun et al., 2001) and the properties of rod-cone interactions show wide variation (Buck, 2004). This is not surprising, given that rod and cone photoreceptor signals are transmitted to the brain in shared pathways (Daw, Jensen & Bunken, 1990; Sharpe & Stockman, 1999), permitting multiple sites of interaction. The MC pathway is the primary transmitter of information about spatial contrast at mesopic and scotopic illumination levels (Purpura, Kaplan & Shapley, 1988). In this study, we used (L+M+S) cone stimuli based on our previous observation (Cao et al., 2006) that lateral suppressive rod-cone interactions were strongest for stimuli containing luminance variations. From this, we concluded that the change in sensitivity to such stimuli was likely mediated via the MC pathway, consistent with evidence from physiological studies that rod inputs are predominant in MC ganglion cells (Lee, Smith, Pokorny & Kremers, 1997).
Lateral rod-cone interactions in temporal processing depend critically on the spatial frequency of spatially extended sinusoidal grating stimuli (Lange et al., 1997). For spatial frequencies of 1 or 2 cpd, the largest suppression occurs at 8-16 Hz (Lange et al., 1997), consistent with our finding in the temporal contrast sensitivity comparison between the dark adapted and cone plateau conditions (see inserts to Figure 2). At lower temporal frequencies (below 6- 8 Hz), the sinusoidal temporal contrast sensitivity data converged (Figure 2). Evidence for qualitative asymmetries in flicker sensitivity at light onset and offset with low temporal frequency stimuli show a dependence on the adaptation conditions and modulation type. Under light adapted conditions, cone sensitivity to rapid-off sawtooth flicker may be greater than sensitivity to rapid-on flicker (Bowen, Pokorny & Smith, 1989; Bowen, Pokorny, Smith & Fowler, 1992; Frumkes, Lange, Denny & Beczkowska, 1992). When rods are progressively dark-adapted, rapid-on and rapid-off sawtooth flicker sensitivity converge at all temporal frequencies (Frumkes et al., 1992). With sinusoidal stimulus modulations, low temporal frequency contrast sensitivity is independent of the illumination level (Bowen et al., 1992; Kelly, 1961), consistent with the data in Figure 2.
The temporal impulse response functions derived from both the pulsed and periodic data showed similar patterns of change when the level of rod activity in the surround was altered. The average time-to-peak of the temporal impulse response of the cone pathway was 28 ms during the cone plateau, similar to estimates derived from published temporal contrast sensitivity and 2-pulse data for comparable retinal illuminances (see Cao et al., 2007). After dark-adaptation, the time-to-peak was delayed by ∼7 ms. Physiological (Schneeweis & Schnapf, 1995; Verweij, Peterson, Dacey & Buck, 1999) and psychophysical (Cao et al., 2007; Sun et al., 2001) latency difference estimates of the rod and cone systems under comparable mesopic light levels are less than 20 ms. However, when stimulus conditions include high cone stimulus contrasts and/or greater cone light adaptation, rod-cone latency differences are in the order of 60 - 80 ms (Barbur, 1982; MacLeod, 1972; Sharpe, Stockman & MacLeod, 1989; van den Berg & Spekreijse, 1977). The delay in the time-to-peak of the cone temporal impulse response suggests that lateral suppression of cone temporal vision by dark-adapted rods may be a mechanism for reducing latency differences between the rod and cone systems. This could improve the processing of temporal signals under conditions where both rods and cones contribute to vision. The change in cone sensitivity caused by dark-adapted rods in the area surrounding a stimulus of fixed retinal illuminance is similar to those changes observed when the ambient illumination decreases. In both cases, the temporal response of human vision slows down, temporal integration increases and for the associated IRFs, the time from onset to peak becomes longer (Barlow, 1958; Kelly, 1961; Kelly, 1971; Swanson et al., 1987).
Acknowledgments
Australian Research Council Discovery Project DP0773544, National Eye Institute Research Grant EY00901-35 and a Research to Prevent Blindness Challenge Grant supported this work. Andrew J. Zele is an Australian Research Council APD Fellow. We benefited from discussions with Steven K. Shevell and Vivianne C. Smith.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Alexander KR, Fishman GA. Rod-cone interaction in flicker perimetry. British Journal of Ophthalmology. 1984;68:303–309. doi: 10.1136/bjo.68.5.303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barbur JL. Reaction-time determination of the latency between visual signals generated by rods and cones. Ophthalmic and Physiological Optics. 1982;2:179–185. [PubMed] [Google Scholar]
- Barlow HB. Temporal and spatial summation in human vision at different background intensities. Journal of Physiology (London) 1958;141:337–350. doi: 10.1113/jphysiol.1958.sp005978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bowen RW, Pokorny J, Smith VC. Sawtooth contrast sensitivity: Decrements have the edge. Vision Research. 1989;29:1501–1509. doi: 10.1016/0042-6989(89)90134-x. [DOI] [PubMed] [Google Scholar]
- Bowen RW, Pokorny J, Smith VC, Fowler MA. Sawtooth contrast sensitivity: Effects of mean illuminance and low temporal frequencies. Vision Research. 1992;32:1239–1247. doi: 10.1016/0042-6989(92)90218-8. [DOI] [PubMed] [Google Scholar]
- Buck SL. Rod-cone interactions in human vision. In: Chalupa LM, Werner JS, editors. The Visual Neurosciences. Vol. 1. Cambridge MA: MIT Press; 2004. pp. 863–878. [Google Scholar]
- Burr DC, Morrone MC. Impulse-Response Functions for Chromatic and Achromatic Stimuli. Journal of the Optical Society of America A. 1993;10(8):1706–1713. [Google Scholar]
- Cao D, Zele AJ, Pokorny J. Dark-adapted rod suppression of cone flicker detection: Evaluation of receptoral and postreceptoral interactions. Visual Neuroscience. 2006;23(34):531–537. doi: 10.1017/S0952523806233376. [DOI] [PubMed] [Google Scholar]
- Cao D, Zele AJ, Pokorny J. Linking impulse response functions to reaction time: Rod and cone reaction time data and a computational model. Vision Research. 2007;47(8):1060–1074. doi: 10.1016/j.visres.2006.11.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cao D, Zele AJ, Pokorny J. Chromatic discrimination in the presence of incremental and decremental rod pedestals. Visual Neuroscience. 2008 doi: 10.1017/S0952523808080425. In Press. [DOI] [PubMed] [Google Scholar]
- Cao DC, Pokorny J, Smith VC. Matching rod percepts with cone stimuli. Vision Research. 2005;45(16):2119–2128. doi: 10.1016/j.visres.2005.01.034. [DOI] [PubMed] [Google Scholar]
- CIE. Publ CIE No.41 (TC-1.4) Bureau Central de la CIE; Paris: 1978. Light as a true visual quantity: principles of measurement. TC-1.4. [Google Scholar]
- Coletta NJ, Adams AJ. Rod Cone Interaction in Flicker Detection. Vision Research. 1984;24(10):1333–1340. doi: 10.1016/0042-6989(84)90188-3. [DOI] [PubMed] [Google Scholar]
- Coletta NJ, Adams AJ. Loss of flicker sensitivity on dim backgrounds in normal and dichromatic observers. Investigative Ophthalmology and Visual Science. 1985;26:S187. [Google Scholar]
- Dacey DM, Lee BB, Stafford DK, Pokorny J, Smith VC. Horizontal cells of the primate retina: Cone specificity without spectral opponency. Science. 1996;271:656–659. doi: 10.1126/science.271.5249.656. [DOI] [PubMed] [Google Scholar]
- Daw NW, Jensen EJ, Bunken WJ. Rod pathways in the mammalian retinae. Trends in Neuroscience. 1990;13(3):110–115. doi: 10.1016/0166-2236(90)90187-f. [DOI] [PubMed] [Google Scholar]
- de Lange H. Research into the dynamic nature of the human fovea-cortex systems with intermittent and modulated light. I. Attenuation characteristics with white and colored light. Journal of the Optical Society of America. 1958;48:777–784. doi: 10.1364/josa.48.000777. [DOI] [PubMed] [Google Scholar]
- Frumkes TE. The Science of Vision. New York, NY: Springer-Verlag New York, Inc.; 1990. Suppressive rod-cone interaction; pp. 194–210. [Google Scholar]
- Frumkes TE, Eysteinsson T. The Cellular Basis for Suppressive Rod Cone Interaction. Visual Neuroscience. 1988;1(3):263–273. doi: 10.1017/s0952523800001929. [DOI] [PubMed] [Google Scholar]
- Frumkes TE, Lange G, Denny N, Beczkowska I. Influence of rod adaptation upon cone responses to light offset in humans: I. Results in normal observers. Visual Neuroscience. 1992;8(2):83–89. doi: 10.1017/s095252380000924x. [DOI] [PubMed] [Google Scholar]
- Frumkes TE, Naarendorp F, Goldberg SH. Abnormalities in retinal neurocircuitry in protanopes: Evidence provided by psychophysical investigation of temporal-spatial interaction. Investigative Ophthalmology and Visual Science. 1988;29:S163. [Google Scholar]
- Goldberg SH, Frumkes TE, Nygaard RW. Inhibitory influence of unstimulated rods in the human retina: evidence provided by examining cone flicker. Science. 1983;221:180–182. doi: 10.1126/science.6857279. [DOI] [PubMed] [Google Scholar]
- Ikeda M. Temporal impulse response. Vision Research. 1986;26:1431–1440. doi: 10.1016/0042-6989(86)90166-5. [DOI] [PubMed] [Google Scholar]
- Keesey UT. Variables determining flicker sensitivity in small fields. Journal of the Optical Society of America. 1970;60:390–398. doi: 10.1364/josa.60.000390. [DOI] [PubMed] [Google Scholar]
- Kelly DH. Visual responses to time-dependent stimuli: I. Amplitude sensitivity measurements. Journal of the Optical Society of America. 1961;51:422–429. doi: 10.1364/josa.51.000422. [DOI] [PubMed] [Google Scholar]
- Kelly DH. Flickering patterns and lateral inhibition. Journal of the Optical Society of America. 1969:1361–1365. [Google Scholar]
- Kelly DH. Theory of flicker and transient responses. Journal of the Optical Society of America. 1971;61:537–546. doi: 10.1364/josa.61.000537. [DOI] [PubMed] [Google Scholar]
- Kremers J, Meierkord S. Rod-cone-interactions in deuteranopic observers: models and dynamics. Vision Research. 1999;39:3372–3385. doi: 10.1016/s0042-6989(99)00027-9. [DOI] [PubMed] [Google Scholar]
- Lange G, Denny N, Frumkes TE. Suppressive rod-cone interactions: evidence for separate retinal (temporal) and extraretinal (spatial) mechanisms in achromatic vision. Journal of the Optical Society of America A. 1997;14(9):2487–2498. doi: 10.1364/josaa.14.002487. [DOI] [PubMed] [Google Scholar]
- Lee BB, Smith VC, Pokorny J, Kremers J. Rod inputs to macaque ganglion cells. Vision Research. 1997;37:2813–2828. doi: 10.1016/s0042-6989(97)00108-9. [DOI] [PubMed] [Google Scholar]
- Lythgoe RJ, Tansley K. The relation of the critical frequency of flicker to the adaptation of the eye. Proceedings of the Royal Society (London) 1929;105B:60–92. [Google Scholar]
- MacLeod DI. Rods cancel cones in flicker. Nature. 1972;235(5334):173–174. doi: 10.1038/235173a0. [DOI] [PubMed] [Google Scholar]
- Naarendorp F, Frumkes T. The influence of short-term adaptation of human rods and cones on cone-mediated grating visibility. Journal of Physiology (London) 1991;432:521–541. doi: 10.1113/jphysiol.1991.sp018398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pokorny J, Smithson H, Quinlan J. Photostimulator allowing independent control of rods and the three cone types. Visual Neuroscience. 2004;21(3):263–267. doi: 10.1017/s0952523804213207. [DOI] [PubMed] [Google Scholar]
- Pugh EN. Rushton's paradox: Rod dark adaptation after flash photolysis. Journal of Physiology (London) 1975;248:413–431. doi: 10.1113/jphysiol.1975.sp010982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Purpura K, Kaplan E, Shapley RM. Background light and the contrast gain of primate P and M retinal ganglion cells. Proceedings of the National Academy of Sciences USA. 1988;85:4534–4537. doi: 10.1073/pnas.85.12.4534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Puts MJ, Pokorny J, Quinlan J, Glennie L. Audiophile hardware in vision science; the soundcard as a digital to analog converter. Journal of Neuroscience Methods. 2005;142(1):77–81. doi: 10.1016/j.jneumeth.2004.07.013. [DOI] [PubMed] [Google Scholar]
- Schneeweis DM, Schnapf JL. Photovoltage of rods and cones in the macaque retina. Science. 1995;268:1053–1056. doi: 10.1126/science.7754386. [DOI] [PubMed] [Google Scholar]
- Shapiro AG, Pokorny J, Smith VC. Cone-Rod receptor spaces, with illustrations that use CRT phosphor and light-emitting-diode spectra. Journal of the Optical Society of America A. 1996;13:2319–2328. doi: 10.1364/josaa.13.002319. [DOI] [PubMed] [Google Scholar]
- Sharpe LT, Stockman A. Rod pathways: the importance of seeing nothing. Trends in Neuroscience. 1999;22:497–504. doi: 10.1016/s0166-2236(99)01458-7. [DOI] [PubMed] [Google Scholar]
- Sharpe LT, Stockman A, MacLeod DI. Rod flicker perception: scotopic duality, phase lags and destructive interference. Vision Research. 1989;29:1539–1559. doi: 10.1016/0042-6989(89)90137-5. [DOI] [PubMed] [Google Scholar]
- Smith VC, Pokorny J. The design and use of a cone chromaticity space. Color Research and Application. 1996;21:375–383. [Google Scholar]
- Stork DG, Falk DS. Visual temporal impulse response functions from flicker sensitivities. Journal of the Optical Society of America A. 1987;4:1130–1135. doi: 10.1364/josaa.4.001130. [DOI] [PubMed] [Google Scholar]
- Sun H, Pokorny J, Smith VC. Rod-cone interaction assessed in inferred magnocellular and parvocellular postreceptoral pathways. Journal of Vision. 2001;1:42–54. doi: 10.1167/1.1.5. http://www.journalofvision.org/1/1/5/ [DOI] [PubMed]
- Swanson WH, Ueno T, Smith VC, Pokorny J. Temporal modulation sensitivity and pulse detection thresholds for chromatic and luminance perturbations. Journal of the Optical Society of America A. 1987;4:1992–2005. doi: 10.1364/josaa.4.001992. [DOI] [PubMed] [Google Scholar]
- Temme LA, Frumkes TE. Rod-cone interaction in human scotopic vision--III: Rods influence cone increment thresholds. Vision Research. 1977;17(6):681–685. doi: 10.1016/s0042-6989(77)80002-3. [DOI] [PubMed] [Google Scholar]
- van den Berg TJTP, Spekreijse H. Interaction between rod and cone signals studied with temporal sine wave stimulation. Journal of the Optical Society of America. 1977;67:1210–1217. doi: 10.1364/josa.67.001210. [DOI] [PubMed] [Google Scholar]
- Verweij J, Peterson BB, Dacey DM, Buck SL. Sensitivity and dynamics of rod signals in H1 horizontal cells of the macaque monkey retina. Vision Research. 1999;39:3662–3672. doi: 10.1016/s0042-6989(99)00093-0. [DOI] [PubMed] [Google Scholar]
- Wolf E, Zigler MJ. Location of the break in the dark adaptation curve in relation to pre-exposure brightness and pre-exposure time. Journal of the Optical Society of America. 1954;44(11):875–879. doi: 10.1364/josa.44.000875. [DOI] [PubMed] [Google Scholar]
- Zele AJ, Cao D, Pokorny J. Threshold units: A correct metric for reaction time? Vision Research. 2007;47(5):608–611. doi: 10.1016/j.visres.2006.12.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zele AJ, Vingrys AJ. Defining the detection mechanisms for symmetric and rectified flicker stimuli. Vision Research. 2007;47(21):2700–2713. doi: 10.1016/j.visres.2007.05.005. [DOI] [PubMed] [Google Scholar]