

Abstract
Photosynthesis, biological nitrogen fixation, and carbon dioxide assimilation are three fundamental biological processes catalyzed by photosynthetic bacteria. In the present study, it is shown that mutant strains of the nonsulfur purple photosynthetic bacteria Rhodospirillum rubrum and Rhodobacter sphaeroides, containing a blockage in the primary CO2 assimilatory pathway, derepress the synthesis of components of the nitrogen fixation enzyme complex and abrogate normal control mechanisms. The absence of the Calvin–Benson–Bassham (CBB) reductive pentose phosphate CO2 fixation pathway removes an important route for the dissipation of excess reducing power. Thus, the mutant strains develop alternative means to remove these reducing equivalents, resulting in the synthesis of large amounts of nitrogenase even in the presence of ammonia. This response is under the control of a global two-component signal transduction system previously found to regulate photosystem biosynthesis and the transcription of genes required for CO2 fixation through the CBB pathway and alternative routes. In addition, this two-component system directly controls the ability of these bacteria to grow under nitrogen-fixing conditions. These results indicate that there is a molecular link between the CBB and nitrogen fixation process, allowing the cell to overcome powerful control mechanisms to remove excess reducing power generated by photosynthesis and carbon metabolism. Furthermore, these results suggest that the two-component system integrates the expression of genes required for the three processes of photosynthesis, nitrogen fixation, and carbon dioxide fixation.
Keywords: global regulation, CO2 fixation
Three important processes that sustain the biosphere are photochemical energy conversion, the reductive assimilation of carbon dioxide, and biological nitrogen fixation. Some microorganisms (limited to certain eubacteria) catalyze all three processes, and the nonsulfur purple bacteria are particularly versatile in their metabolism. These organisms thus serve as excellent models to understand the molecular control of the biogeochemical carbon and nitrogen cycles. Although much physiological information has been collected over the years to indicate that photochemical energy generation is linked to carbon assimilation and nitrogen fixation (1, 2), there has never been an unequivocal direct molecular link that integrates the control of genes required for two or more of these processes. Certainly, the molecular integration of the control of all three of these processes has not been heretofore described.
The major route by which CO2 is metabolized in the environment is the Calvin–Benson–Bassham (CBB) reductive pentose phosphate pathway, the key enzyme of which is ribulose 1,5-bisphosphate carboxylase/oxygenase (RubisCO). RubisCO-mediated CO2 fixation and the CBB cycle are also extremely important to photosysnthetic bacteria in particular, and probably other organisms as well, by virtue of the fact that CO2 becomes a major electron sink when the bacteria are grown under photosynthetic conditions in the presence of organic carbon (3). Under these conditions, with simple organic compounds used as the major carbon source and electron donor, nitrogen fixation may provide all or part of the nitrogen required for growth. In this paper, we report that a specific two-component response regulator-sensor kinase signal transduction system regulates biological nitrogen fixation. The same phospho-relay system also regulates the expression of genes of the CBB system, the presence or absence of which is shown here to have a profound influence on the control of genes required for nitrogen fixation. These studies suggest a direct role of this phospho-relay system in controlling the interaction of the CBB and nitrogen fixation systems of nonsulfur purple bacteria.
MATERIALS AND METHODS
Bacterial Strains, Plasmids, and Growth Conditions.
The genotype and phenotype of Rhodospirillum rubrum and Rhodobacter sphaeroides strains employed in this study are summarized in Table 1. Strain Str-2 is the wild-type Rs. rubrum strain and is capable of growth under conditions where CO2 may be the sole source of carbon in a CO2/H2 atmosphere (RubisCO+) due to the expression of the form II RubisCO structural gene (cbbM+); N2 may serve as the sole nitrogen source (N2ase+) due to the expression of the nitrogenase structural genes (nifHDK+). Strain I-19 is a RubisCO-deletion strain of Rs. rubrum (cbbM−) and was grown in the absence or the presence of plasmid pJG106, which contains the genes of the cbbII operon of R. sphaeroides and provides a functional cbbM gene (5). Three different wild-type strains of R. sphaeroides were used (2.4.1, 2.4.7, and HR). These strains are all capable of CO2-dependent growth in a CO2/H2 atmosphere due to the expression of the form I RubisCO (cbbLS+) and form II RubisCO (cbbM+) and may use N2 as the sole nitrogen source for growth. All mutants of R. sphaeroides were derived from strain HR and some strains were grown in the presence of 40 mM DMSO as alternate electron acceptor. Strain 16 is a RubisCO− strain (cbbLS−cbbM−) that cannot use CO2 as electron acceptor; strain 16PHC was derived from strain 16 (6). Strain 16PHCΩ contains a specific deletion in the regB gene (Y. Qian and F.R.T., unpublished results). Strain A25 is a transposon (Tn5) mutant of 16PHC, which has the phenotype of strain 16 due to a transposition in the regB gene (7). Strain 1312 contains a specific disruption of the cbbR gene (8) resulting in no synthesis of form I RubisCO (cbbLS−) under any growth conditions and diminished (about 30%) of form II RubisCO (cbbM) when grown photosynthetically in the presence of malate. This strain is incapable of growth in a CO2/H2 atmosphere (8).
Table 1.
Nitrogenase activity (acetylene reduction) in strains of Rs. rubrum and R. sphaeroides
| Strain | Genotype/phenotype | Nitrogenase
activity, nmol C2H4 produced per min per mg
protein
|
|
|---|---|---|---|
| Malate (ammonium sulfate) | Malate (glutamate) | ||
| Rs. rubrum | |||
| Str-2 | cbbM+/PA+PH+RubisCO+N2ase+ | 0 | 40 |
| I-19 | cbbM−/PA−PH+RubisCO−N2ase+ | 24 | 106 |
| I-19 (pJG106) | cbbM+/PA+PH+RubisCO+N2ase+ | 0 | 56 |
| R. sphaeroides | |||
| 2.4.1 | cbbLS+cbbM+regB+/PA+PH+RubisCO+N2ase+ | 0 | 52 |
| 2.4.7 | cbbLS+cbbM+regB+/PA+PH+RubisCO+N2ase+ | 0 | 60 |
| HR | cbbLS+cbbM+regB+/PA+PH+RubisCO+N2ase+ | 0 | 0 |
| HR + DMSO | cbbLS+cbbM+regB+/PA+PH+RubisCO+N2ase+ | 0 | 0 |
| 16 + DMSO | cbbLS−cbbM−regB+/PA−PH−RubisCO−N2ase− | 0 | 0 |
| 16PHC | cbbLS−cbbM−regB+/PA−PH+RubisCO−N2ase+ | 36 | 43 |
| 16PHC + DMSO | cbbLS−cbbM−regB+/PA−PH+RubisCO−N2ase+ | 29 | 40 |
| 16PHC(pJG106) | cbbLS−cbbM+regB+/PA+PH+RubisCO+N2ase+ | 0 | ND |
| 16PHCΩ + DMSO | cbbLS−cbbM−regB−/PA−PH−RubisCO−N2ase− | 0 | 0 |
| A25 + DMSO | cbbLS−cbbM−regB−/PA−PH−RubisCO−N2ase− | 0 | 0 |
| 1312 | cbbLS−cbbM+regB+/PA−PH+RubisCO+N2ase+ | 0 | 30 |
Each strain was grown under photosynthetic conditions in a growth medium containing malate and the indicated nitrogen source (4). In all cases, cells were incubated in the presence of 10% acetylene under anoxic conditions in the light, and the amount of ethylene produced was determined by gas chromatography by taking several time points over a 60-min assay. Strains HR, 16PHC, and 1312 possessed nitrogenase activity (30–50 nmol C2H4 per min per mg protein) when grown in a malate medium using N2 as sole nitrogen source. PA, growth in the absence of organic carbon under photosynthetic conditions where CO2 is the sole source of carbon. PH, growth in the presence of organic carbon under photosynthetic conditions; PA+ or PH+ refers to growth conditions where CO2 is used as an electron acceptor as well as a carbon source; PH−, growth conditions where CO2 cannot be used as an electron acceptor and some alternative acceptor [e.g., dimethyl sulfoxide (DMSO)] must be employed in a malate medium. ND, not determined.
Nitrogenase Assays.
Before assay, cells from the growth medium were washed three times under anoxic conditions and then resuspended in the same fresh medium in which they were grown. A portion of the cell suspension was then injected into vials containing an atmosphere of 10% acetylene in argon. Samples were removed from the illuminated vials, and the amount of ethylene produced was determined by gas chromatography; several time points were taken over a 60-min assay. Immunoblots, using polyclonal antibodies directed against the Rs. rubrum dinitrogenase reductase and dinitrogenase, were prepared by using established procedures (8), as was the processing of cell extracts for gel electrophoresis and subsequent immunodetection.
RESULTS AND DISCUSSION
In the nonsulfur purple bacteria Rs. rubrum and R. sphaeroides, the genes that encode RubisCO were deleted such that the resultant knock-out strains were not able to use the CBB route to assimilate CO2 (5, 6). Unlike the wild type, we recently observed that the knock-out strains produced copious quantities of molecular hydrogen, suggesting the unmasking of a hydrogenase. Because hydrogen is a well-known product of the nitrogenase enzyme complex of photosynthetic bacteria (9), we determined whether or not the Rs. rubrum and R. sphaeroides RubisCO knock-out strains (I-19 and 16PHC, respectively) possessed nitrogenase activity (Table 1). Both I-19 and 16PHC contained significant nitrogenase activity, despite the fact that both strains were grown by using ammonia as the nitrogen source. Neither of the ammonia-grown wild-type strains of Rs. rubrum or R. sphaeroides contain any detectable nitrogenase activity, consistent with the known repressive effect of ammonia metabolism on the transcription of the nif genes. Interestingly, the expression of genes carried on plasmid pJG106, which contains a fully functional RubisCO gene as well as structural genes of other enzymes of the reductive CO2 assimilatory path of R. sphaeroides, prevented Rs. rubrum strain I-19 from producing any detectable nitrogenase activity. Strain 16 of R. sphaeroides, the original RubisCO deletion strain, of which strain 16PHC is a spontaneous derivative (3), is not able to grow photosynthetically on malate without an exogenous electron acceptor such as DMSO (6). Strain 16 was also shown to be incapable of supporting nitrogenase activity when grown in the presence of ammonia (Table 1).
Because of the intriguing correlation between the absence of RubisCO and the enhancement of nitrogenase activity in strain 16PHC cells grown in the presence of ammonia, we determined whether mutations that affect RubisCO synthesis actually influence nitrogenase activity in such strains. Previous work had shown that mutations in the regB gene of R. sphaeroides blocked transcription of the cbb regulon (7), which contains genes that encode both forms of RubisCO and other enzymes of the CBB cycle. Interestingly, inactivation of the regB gene of R. sphaeroides abrogated the capacity of strain 16PHC to produce active nitrogenase in the normally repressive ammonia media; strain A25, which contains a Tn5 insertion in the regB gene (7), behaved similarly. Another mutant, strain 1312, which contains an inactivated cbbR gene (8), encoding a positive transcription regulator of the cbb regulon, has a greatly diminished capacity to synthesize enzymes of the CBB pathway and contains about 30% of the normal amount of form II RubisCO, but no form I RubisCO. It is apparent that this low level of RubisCO is enough to prevent strain 1312 from producing active nitrogenase in the presence of ammonia, presumably because the reductive CO2 assimilatory path is functional. It is also curious that strain HR and its derivatives did not contain detectable nitrogenase activity when glutamate served as the source of nitrogen, unlike other wild-type strains of R. sphaeroides. However, in contrast to parent strain HR, strain 1312 synthesized nitrogenase when grown in the presence of glutamate.
To establish that the appearance of nitrogenase activity by cbb mutant cells grown in the presence of ammonia was the result of enzyme synthesis, we used antibodies to each component of the nitrogenase enzyme complex and examined the proteins from crude extracts of several strains. It was apparent that wild-type Rs. rubrum, grown with ammonium sulfate as the source of nitrogen, did not synthesize either component I, the molybdo-iron protein (dinitrogenase), or component II, the iron protein (dinitrogenase reductase) of the complex (Fig. 1 A and B, lanes 2 and 6). However, strain I-19 synthesized both components, in both malate and acetate-containing growth media (Fig. 1 A and B, lanes 3 and 7). Both the wild type and strain I-19 synthesized nitrogenase components in the presence of glutamate, as expected. Strain I-19, complemented with plasmid pJG106 (Fig. 1 A and B, lanes 8) did not synthesize nitrogenase components when grown in the presence of ammonia, presumably because expression of the plasmid-borne form II RubisCO gene led to a complete reductive assimilatory pathway, much like the wild type. A strain of Rs. rubrum, I-32, containing copies of both the inactivated and wild-type RubisCO gene (5), also did not synthesize nitrogenase in the presence of ammonia. In R. sphaeroides, several more mutants are available and provide additional insights on the regulation of the system. Similar to Rs. rubrum, the 16PHC mutant of R. sphaeroides synthesized high levels of nitrogenase components when cultured in the presence of ammonia (Fig. 2 A and C, lanes 4 and 5), conditions that failed to support nitrogenase synthesis for the wild type and strain 16 (Fig. 2 A and C; lanes 2, 3, and 7). However, strain 16PHCΩ, in which the regB gene was disrupted (Y. Qian and F.R.T., unpublished results) was incapable of derepressing the synthesis of nitrogenase in the presence of ammonia (Fig. 2 A and C; lanes 8). A similar result was obtained for strain A25, which is a transposon (Tn5) mutant strain of 16PHC (7), selected for its inability to grow using CO2 as electron acceptor, much like parent strain 16. Since the transposon was found to be inserted within the coding sequence of the regB gene of strain A25 and is phenotypically similar to strain 16PHCΩ, it is apparent that this sensor kinase gene may be involved in the derepression of nitrogenase in response to the intracellular redox level, even in the presence of normal repressive levels of ammonia. The cbbR mutant, strain 1312, did not synthesize nitrogenase under these conditions, in agreement with the activity measurements (Table 1).
Figure 1.
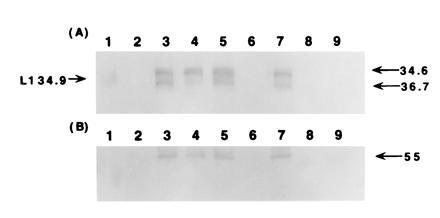
Immunoblot analysis of dinitrogenase and dinitrogenase reductase synthesis in Rs. rubrum. The gel blot was reacted with antiserum to Rs. rubrum dinitrogenase reductase (A) and dinitrogenase (B). Samples were of crude extracts from the wild-type grown on malate and ammonium sulfate (lanes 2), malate and glutamate (lanes 4), and acetate and ammonium sulfate (lanes 6); strain I-19 grown on malate and ammonium sulfate (lanes 3), malate and glutamate (lanes 5), and acetate and ammonium sulfate (lanes 7); strain I-19 (pJG106) grown on malate and ammonium sulfate (lanes 8); and strain I-32, grown on malate and ammonium sulfate (lanes 9). A prestained molecular weight marker of 34.9 kDa (lanes 1) is indicated by the left arrow in the margin and the position of the calculated molecular weights (34.6 and 36.7 kDa) of the subunits of dinitrogenase reductase, presumably modified and unmodified (10), are indicated by the arrows in the right margin.
Figure 2.

Immunoblot analysis of dinitrogenase and dinitrogenase reductase synthesis in R. sphaeroides. Antisera to dinitrogenase (A and B) and dinitrogenase reductase (C and D) were used in immunoblots of samples from R. sphaeroides grown in a malate medium using ammonium sulfate (A and C) and glutamate (B and D) as the nitrogen source: wild-type, strain HR grown in the absence (lanes 2) or presence (lanes 3) of DMSO as electron acceptor; strain 16PHC in the absence (lanes 4) or presence (lanes 5) of DMSO; strain PHG, a RubisCO− derivative of 16PHC which does not use CO2 as electron acceptor (lanes 6); strain 16 plus DMSO (lanes 7); strain 16PHCΩ plus DMSO (lanes 8); strain A25 plus DMSO (lanes 9); and strain 1312 (lanes 10). Prestained molecular weight markers (lanes 1) and approximate molecular weights of dinitrogenase and dinitrogenase reductase are shown in the right margin.
To ensure that the derepression of nitrogenase synthesis in the cbb mutants was not merely the result of a defect in ammonia transport or metabolism, we first examined the growth rates of Rs. rubrum Str-2 and I-19 in the presence of 15 mM ammonium sulfate. Both strains grew with precisely the same doubling time. Moreover, analysis of the spent growth medium of cultures of both strains harvested at similar turbidities in stationary phase indicated that both cultures contained 6 mM ammonia; however, only cultures of strain I-19 showed substantial gas production. Similar results were obtained with R. sphaeroides HR and 16PHC (Y. Qian and F.R.T., unpublished observations). These results, with both Rs. rubrum and R. sphaeroides, suggest that growth and ammonia uptake and metabolism is normal in strains I-19 and 16PHC; however, further metabolic studies with both R. sphaeroides and Rs. rubrum are warranted because products of ammonia assimilation have historically been implicated in the regulation of nitrogen metabolism (11).
The results obtained with R. sphaeroides are particularly revealing because they indicate the importance of the two-component, response regulator (regA/prrA)-sensor kinase (regB/prrB) system (12–15) for the derepression of nitrogenase synthesis in cbb strain 16PHC. Specific knock-out mutants of regB in wild-type strain HR and strain 16PHC, as well as Tn5 mutant strain A25, all fail to grow using molecular nitrogen (Fig. 3). Thus, mutations in the regB gene exhibit two specific effects relative to the nitrogenase system of R. sphaeroides, the capacity of strain 16PHC to derepress nitrogenase synthesis and the ability of strains to grow with dinitrogen in the presence or absence of a functional CBB CO2 reduction system. Derepression occurs only in strains that have retained the ability to grow on malate in the absence of the alternative electron acceptor (DMSO) yet lack functional RubisCO (the key enzyme of the CBB reductive CO2 assimilatory path). In the absence of a complete CBB cycle, strain 16PHC is unable to remove the excess reducing equivalents derived from photosynthesis and the oxidation of malate. Thus CO2 is no longer capable of functioning as the major electron sink, and some alternative redox system for the dissipation of the large amount of reducing power must be employed. It is apparent from our results that this alternative redox system is the nitrogenase complex (Fig. 4). This enzyme system is responsible for reducing atmospheric dinitrogen to ammonia and protons to hydrogen; this system requires much reducing power and energy according to the following relationship (17):
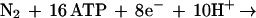 |
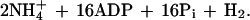 |
Under normal nonrepressive conditions—e.g., in the absence of ammonia—the nitrogenase enzyme complex (encoded by the nifHDK structural genes) is synthesized at extremely high levels (11, 18) to enable the cell to obtain all required nitrogen via the reduction of dinitrogen. Most important, at least 25% of the electron throughput is used to reduce protons to molecular hydrogen (19), which is released from the cells by photosynthetic bacteria under anoxic conditions. Thus, the nitrogenase, and its inherent hydrogenase activity, acts as a safety valve for the dissipation of excess reducing equivalents (2) and in the absence of N2 these reducing equivalents may be funneled totally to the reduction of protons. However, the nitrogenase complex is not normally synthesized when the cells are provided with fixed nitrogen in the form of ammonia (18). Thus, it is apparent from the results of this study that the normal molecular control mechanisms that serve to prevent nif transcription and nitrogenase synthesis in the presence of ammonia are overcome by the organism’s need to remove excess reducing equivalents in the absence of a functional CBB cycle, controlled by the cbb regulon (20) (Fig. 4). Exactly how nif transcription control is altered in the RubisCO− strains remains to be discovered. However, it is apparent that control is somehow mediated by the RegA/RegB system. The fact that a regB mutation in both wild type (RubisCO positive) and strain 16PHC (RubisCO−) were unable to grow at the expense of dinitrogen (Fig. 3) indicates that the absence of regB prevents nif transcription under otherwise favorable conditions, similar to its effect on both cbb transcription (7) and the transcription of genes required for photosystem biosynthesis (12–15). It is important to emphasize that the RegA/RegB system contributes to the ability of R. sphaeroides to regulate derepression of the nif system in the presence of ammonia only in the absence of a functional CBB reductive CO2 assimilatory pathway. The significance of regB is underlined by the fact that only strain 16PHC, and not strains 16PHCΩ or A25, which both contain mutations in regB, is able to derepress nitrogenase synthesis. For unknown reasons, strain 16, although it is a RubisCO− strain, cannot employ the nitrogenase complex as an alternative electron acceptor. Once the molecular basis for the change from strain 16 to strain 16PHC (3) is understood, it should become apparent why strain 16 cannot employ the nitrogenase enzyme complex as an alternative electron acceptor. It is also apparent that strain 16PHC is a key to elucidating the precise mechanism by which cbb transcription influences the expression of genes involved in nitrogenase biosynthesis. The current studies suggest that control of the cbb regulon, and a functioning CBB cycle (7), are extremely important to these organisms, not just in terms of carbon metabolism, but also for maintaining redox control, with the RegA/RegB two-component system implicated in controlling cbb and nif transcription both individually and in tandem. It should be emphasized that the studies reported here take advantage of the nutritional diversity of the nonsulfur purple bacteria, where knock-out mutations in genes specifying key steps in each of the three processes are nonlethal under nonrestrictive growth conditions. Thus, Rhodobacter and Rhodospirillum represent excellent model systems to study the interplay of photosynthesis, carbon dioxide assimilation, and nitrogen fixation, three processes that are especially important for the cycling of carbon and nitrogen in the earth’s biosphere; the discovery of a common regulatory circuit should lead to further advances on how these processes are linked at the molecular level. Although these bacteria themselves are probably not significant on a global scale, close phylogenetic relatives of Rhodobacter such as symbiotic root nodule bacteria, Rhizobium and Bradyrhizobium species (21), play significant roles in global nitrogen transformations, while various lithoautotrophic bacteria substantially influence carbon cycling. Because these organisms use the CBB cycle to reduce CO2, it may be that the cbb and nif systems are similarly coregulated in these bacteria as well. Reciprocal diel regulation of nitrogen fixation, photosynthesis, and carbon dioxide fixation is also well established in various cyanobacteria and picoplankton (22) and at least one molecular link between nitrogen metabolism (nitrate reduction) and carbon dioxide fixation has been found in certain species (23). By extension to the current studies, it may be fruitful to determine if a similar global regulator system functions in these organisms.
Figure 3.
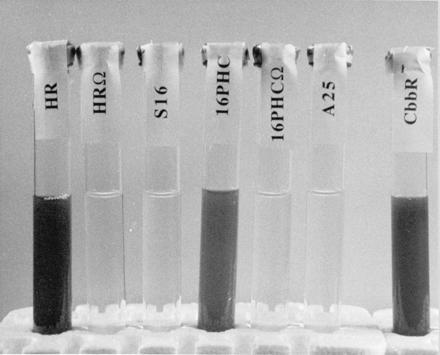
Growth of R. sphaeroides in the absence of fixed nitrogen. The indicated strains were grown in a 2% H2/98% N2 atmosphere in a malate minimal medium in the light after equilibrating and inoculating growth tubes in an anoxic glove box of the same atmosphere.
Figure 4.
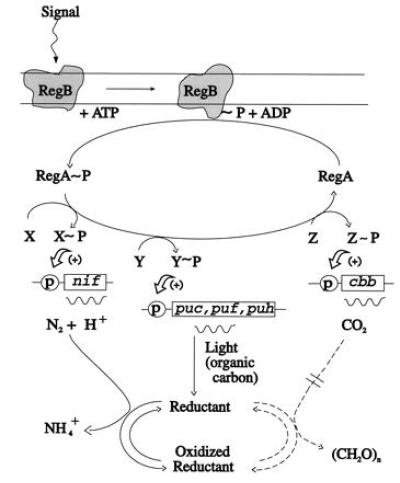
A model which illustrates the super regulation in R. sphaeroides of three processes: photosynthesis, CO2 assimilation, and N2 fixation. Regulation of each process is affected by the RegA/RegB two component signal transduction system with RegB the sensor kinase and RegA the response regulator component. For each process, hypothetical unknown component(s) (X, Y, and Z) are required to transduce the signal from RegA to specific sequences that control the Calvin–Benson–Bassham (cbb), photosystem (puf, puh, puc), and nitrogen (nif) regulons, respectively. Photochemical reactions and organic carbon oxidation both might generate large amounts of reductant. When the CBB pathway is blocked in the RubisCO deletion mutants, excess reducing power is directed to the nitrogenase system [also see Mosley et al. (13) and Bauer and Bird (16)].
Because the RegA/RegB system was originally discovered to control the ability of Rhodobacter to respond to different intensities of light (12–15), it is apparent that a common signal is transduced through this two-component regulatory system to influence the molecular events required for photosynthesis (photosystem biosynthesis) (12–15), carbon dioxide reduction (7), and now nitrogen fixation (Fig. 4). The model illustrates the importance of the phospho-relay signal transduction system in regulating each process and also shows how nif derepression in the presence of ammonia is influenced by cbb transcription and RegB. Thus, the need for these organisms to seek alternative mechanisms to dispense with the large amounts of reducing power generated via photosynthesis and the oxidation of organic carbon (malate) is paramount, with the nif system turned on when the CBB route is blocked, even in the presence of normally repressive levels of ammonia. By the same token, the expression of a functional RubisCO gene to complete the CBB pathway in strain 16PHC keeps the nif system under ammonia control. RegB is thought to be membrane-associated (24); it is autophosphorylated and it is required to phosphorylate the response regulator, RegA (24). Presumably some external signal initiates the phospho-relay system, with the regulatory cascade influenced by the redox state of the cells, the level of oxygen, and the presence of various carbon and nitrogen sources (7, 12–15). Secondary kinases may “cross-talk” with the system (25) and there must be additional components since RegA does not exhibit known DNA binding motifs (16). Thus, in order for this system to function, some means of transducing the signal from RegA to specific regions of the genome must be available. We thus predict that separate, individual molecular component(s) for each regulon (depicted as X, Y, and Z) may be linked to RegA; obviously further work will be necessary to verify this. In addition, because all three processes of nitrogen fixation, photochemical energy conversion, and carbon dioxide assimilation are dependent on the intracellular redox potential, there may be an additional component(s) that is influenced by the reductant pool that may then influence, in a hierarchial way, the individual regulons through X, Y, and Z.
Acknowledgments
We thank Y. Qian and J. L. Gibson for mutant strains and T. Wahlund and T.-J. Chow for their assistance with the nitrogenase assays. The generous gift of antibodies to dinitrogenase reductase and dinitrogenase by Paul Ludden is gratefully acknowledged. This work was supported by Grant DE-FG02-91ER20033 from the Department of Energy and Grant GM45404 from the National Institutes of Health.
Footnotes
Abbreviations: CBB pathway, Calvin–Benson–Bassham pathway; RubisCO, ribulose 1,5-bisphosphate carboxylase/oxygenase; DMSO, dimethyl sulfoxide.
References
- 1.Ormerod J G, Gest H. Bacteriol Rev. 1962;26:51–66. doi: 10.1128/br.26.1.51-66.1962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Hillmer P, Gest H. J Bacteriol. 1977;129:724–731. doi: 10.1128/jb.129.2.724-731.1977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Wang X, Falcone D L, Tabita F R. J Bacteriol. 1993;175:3372–3379. doi: 10.1128/jb.175.11.3372-3379.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Weaver K E, Tabita F R. J Bacteriol. 1983;156:507–515. doi: 10.1128/jb.156.2.507-515.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Falcone D L, Tabita F R. J Bacteriol. 1993;176:5066–5077. doi: 10.1128/jb.175.16.5066-5077.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Falcone D L, Tabita F R. J Bacteriol. 1991;173:2099–2108. doi: 10.1128/jb.173.6.2099-2108.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Qian Y, Tabita F R. J Bacteriol. 1996;178:12–18. doi: 10.1128/jb.178.1.12-18.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Gibson J L, Tabita F R. J Bacteriol. 1993;175:5778–5784. doi: 10.1128/jb.175.18.5778-5784.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Gest H, Kamen M D. Science. 1954;109:558–559. doi: 10.1126/science.109.2840.558. [DOI] [PubMed] [Google Scholar]
- 10.Ludden P W, Roberts G P. In: Anoxygenic Photosynthetic Bacteria. Blankenship R E, Madigan M T, Bauer C E, editors. Dordrecht, The Netherlands: Kluwer; 1995. pp. 929–947. [Google Scholar]
- 11.Nielsen A H, Nordlund S J. Gen Microbiol. 1975;91:53–62. doi: 10.1099/00221287-91-1-53. [DOI] [PubMed] [Google Scholar]
- 12.Sganga M W, Bauer C E. Cell. 1992;68:945–954. doi: 10.1016/0092-8674(92)90037-d. [DOI] [PubMed] [Google Scholar]
- 13.Mosley C S, Suzuki J Y, Bauer C E. J Bacteriol. 1994;176:7566–7573. doi: 10.1128/jb.176.24.7566-7573.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Eraso J M, Kaplan S. J Bacteriol. 1994;176:32–43. doi: 10.1128/jb.176.1.32-43.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Eraso J M, Kaplan S. J Bacteriol. 1995;177:1258–1267. doi: 10.1128/jb.177.10.2695-2706.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Bauer C E, Bird T H. Cell. 1996;86:5–8. doi: 10.1016/s0092-8674(00)81074-0. [DOI] [PubMed] [Google Scholar]
- 17.Burris R H. J Biol Chem. 1991;266:9339–9342. [PubMed] [Google Scholar]
- 18.Kranz R G, Cullen P J. In: Anoxygenic Photosynthetic Bacteria. Blankenship R E, Madigan M T, Bauer C E, editors. Dordrecht, The Netherlands: Kluwer; 1995. pp. 1191–1208. [Google Scholar]
- 19.Simpson F B, Burris R H. Science. 1984;224:1095–1097. doi: 10.1126/science.6585956. [DOI] [PubMed] [Google Scholar]
- 20.Gibson J L. In: Anoxygenic Photosynthetic Bacteria. Blankenship R E, Madigan M T, Bauer C E, editors. Dordrecht, The Netherlands: Kluwer; 1995. pp. 1107–1124. [Google Scholar]
- 21.Woese C R. Microbiol Rev. 1987;51:221–271. doi: 10.1128/mr.51.2.221-271.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Chow T-J, Tabita F R. J Bacteriol. 1994;176:6281–6285. doi: 10.1128/jb.176.20.6281-6285.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Forchhammer K, Tandeau de Marsac H. J Bacteriol. 1995;177:2033–2040. doi: 10.1128/jb.177.8.2033-2040.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Inoue K, Kouadio C S, Mosley C S, Bauer C E. Biochemistry. 1995;34:391–396. doi: 10.1021/bi00002a002. [DOI] [PubMed] [Google Scholar]
- 25.Gomelsky M, Kaplan S. Microbiology. 1995;141:1805–1819. doi: 10.1099/13500872-141-8-1805. [DOI] [PubMed] [Google Scholar]