

Abstract
In many species, young solicit food from their parents, which respond by feeding them. Because of the difference in genetic make-up between parents and their offspring and the consequent conflict, this interaction is often studied as a paradigm for the evolution of communication. Existent theoretical models demonstrate that chick signaling and parent responding can be stable if solicitation is a costly signal. The marginal cost of producing stronger signals allows the system to converge to an equilibrium: young beg with intensity that reflects their need, and parents use this information to maximize their own inclusive fitness. However, we show that there is another equilibrium where chicks do not beg and parents’ provisioning effort is optimal with respect to the statistically probable distribution of chicks’ states. Expected fitness for parents and offspring at the nonsignaling equilibrium is higher than at the signaling equilibrium. Because nonsignaling is stable and it is likely to be the ancestral condition, we would like to know how natural systems evolved from nonsignaling to signaling. We suggest that begging may have evolved through direct sibling fighting before the establishment of a parental response, that is, that nonsignaling squabbling leads to signaling. In multiple-offspring broods, young following a condition-dependent strategy in the contest for resources provide information about their condition. Parents can use this information even though it is not an adaptation for communication, and evolution will lead the system to the signaling equilibrium. This interpretation implies that signaling evolved in multiple-offspring broods, but given that signaling is evolutionarily stable, it would also be favored in species which secondarily evolved single-chick broods.
Keywords: Nash equilibrium, parent–offspring conflict
Biological communication systems pose a number of intriguing evolutionary questions. In particular, the problem of evolutionary stability (what maintains a communication system viable in the presence of mutants using deviant strategies) is the topic of several recent studies (1–5). Stability, however, is not the only interesting question to ask. When more than one equilibrium can be found, a detailed knowledge of the equilibrium properties of present systems does not explain why the present equilibrium, and not the equally stable alternatives, have evolved. This is specially interesting when conditions that can be inferred to be more primitive than those observed today are shown to be stable. If the original system was stable, why did it evolve into what we see presently? We believe that such a problem is relevant in the case of food begging in birds.
It has long been known that the young of many bird and mammal species solicit food from their parents. This solicitation behavior is so apparent that it is commonly assumed to be costly (see refs. 6–8). As Zahavi (9, 10) points out, this very cost may be the keystone for understanding food solicitation systems. Solicitation costs may act as a handicap that determines the existence of a Nash equilibrium at which offspring solicitation levels are determined by their needs and parental investment is in turn determined by solicitation level (3). [Godfray (3) refers to the signaling equilibrium as an evolutionarily stable strategy (ESS); however, the conditions required for a Nash equilibrium to be an ESS have not been verified anywhere.] We show that in addition to Godfray’s signaling equilibrium, there is a nonsignaling equilibrium at which chicks do not beg in proportion to their needs and parents determine investment according to the expected distribution of needs among chicks in the population. Since nonsignaling is presumably the ancestral condition, this raises the question of what led to the evolutionary change toward the present signaling equilibrium. We advance the hypothesis that a noncommunicatory argument explains this displacement. Furthermore, we show that according to the assumptions in Godfray’s model, chicks and parents at the nonsignaling equilibrium have higher fitness than those at the signaling equilibrium.
The Signaling and Nonsignaling Equilibria
Assume that food solicitations are costly and that offspring condition cannot be inferred by the parents in the absence of signaling. If parents provided a fixed amount of parental care to each of their young, irrespective of their behavior, young would be selected to save as much energy as possible and hence would not solicit food in a costly manner. Likewise, if the young’s needs were not measurable by their parents through solicitation level, parents would be selected to provide a fixed investment set at some optimal level. Hence, the pair of strategies: {if young, don’t solicit food; if parent, invest a fixed amount in your offspring}, constitutes a Nash equilibrium pair (11).
To confirm this point formally, we need a mathematical model. For this purpose, we start from Godfray’s (3) model of signaling of need by offspring to their parents. This model assumes that parents rear a single offspring per litter or brood. Offspring can be in different condition, c, and the expected fitness of an offspring in condition c, which solicits food with intensity x and receives an amount y of food (or any other commodity the parents are providing) is assumed to be:
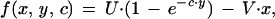 |
1 |
where the (nonnegative) parameter V relates solicitation intensity to its cost and U is a scale parameter, taken to be U = 1 in what follows. Parents have to make a trade-off between investing in their present offspring or in future ones (3); as a result of providing y to their present offspring, their expected future reproductive success is decreased to:
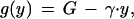 |
2 |
where G is the expected future reproductive success in the absence of breeding and γ reflects the cost of food provisioning. To find the Nash equilibria for this system, we must introduce a third function: the probability distribution function for the condition of the young, ρ(c). The expected inclusive fitness of a soliciting strategy, ℱch “in condition c, solicit food with intensity x(c)” in a population where parents provision their young according to the rule “if your offspring solicits food with intensity x, provide it with y(x)” will be:
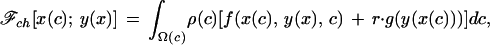 |
3 |
where Ω(c) is the domain of possible conditions and r is the coefficient of relatedness between young and future offspring of their parents. Likewise, the expected inclusive fitness of the provisioning strategy, ℱp, will be:
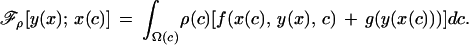 |
4 |
The pair of strategies {x*(c), y*(x)} constitutes a Nash equilibrium if:
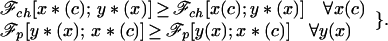 |
5 |
Because the probability distribution function for the condition of the young, ρ (c), is nonnegative on Ω(c), the signaling equilibrium strategies can be obtained by maximizing the integrand of ℱch [x(c); y(x)] with respect to x(c) and the integrand of ℱp[y(x); x(c)] with respect to y(x). By doing this, one obtains the pair of functions:
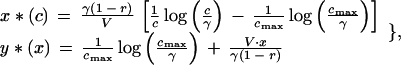 |
6 |
where cmax is the maximum condition that young could possibly attain. Following Godfray (3), let cmax = 2.5 and cmin = 0.5. A cautionary note on the meaning of young condition is required at this point. Eq. 1 tries to capture the fact that there is interindividual variability and that the same amount of parental investment, when devoted to different individuals, leads to different increases in their expected fitness. In practice, there are many features in which individuals may differ: size, age, fat reserves, and resistance to diseases, to name but a few. “Condition” c should not be identified with any one of these features in particular; rather, it is meant to summarize them all for mathematical convenience (3).
Notice that the method described above only looks for local maxima. This is the reason the nonsignaling equilibrium was not revealed: the nonsignaling equilibrium is in the boundary of the space of functions and does not constitute a local maximum. From a mathematical point of view, strategies in which offspring solicited food with negative intensity would be favored. It is only because these strategies make no biological sense that nonsignaling becomes an equilibrium point. Finding the optimal investment level when the offspring are not signaling, yns, is straightforward, because now the expected fitness of the parents is a function of a single variable, rather than a functional (12) as in the signaling case, and the optimal investment level can be obtained using standard calculus methods. If we let the parental fitness in this case, Fp, be:
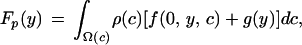 |
7 |
then yns will be determined by the condition:
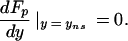 |
8 |
We have so far demonstrated that, in addition to the signaling Nash equilibrium, there is a nonsignaling Nash equilibrium.
In the next section we examine the difference in fitness accruing to each part between these two equilibria. We calculate, for different probability density functions ρ(c), the percentage increase in payoff of being in the nonsignaling equilibrium over being at the signaling equilibrium to parents (Δℱp) and young (Δℱch), defined as:
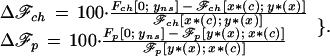 |
9 |
Next we introduce the family of distributions represented in Fig. 1. The members of this family are defined by a single parameter, the kurtosis index, k. For k = 0, the distribution is uniform, and, for k = 1, only young of condition c = 1.5 exist. All the members of the family have mean condition 1.5 and no skew. We express the benefit of being in the nonsignaling equilibrium as the percentage increase in inclusive fitness relative to fitness predicted by Godfray’s model for individuals at the signaling equilibrium. Fig. 2 shows the percentage increase in inclusive fitness derived by parents and offspring from being at the nonsignaling equilibrium plotted against the kurtosis index of the distribution of offspring condition for a wide range of values of the parameter γ. [The value of V has no effect whatsoever for the nonsignaling equilibrium. In the signaling equilibrium, the signaling intensity x appears multiplied by V (Eq. 1) and the product Vx is constant (Eq. 6), so that the value of V has an effect on signaling intensity but not on ensuing fitness.] Rather surprisingly, the expected fitness in the nonsignaling equilibrium is higher than in the signaling one in all but the most extreme conditions. The symmetric distribution of conditions, however, is probably not very realistic. In most populations, the distribution of offspring condition could be either positively or negatively skewed, and this might make signaling profitable. To study this possibility, we introduce another family of distributions, represented in Fig. 3. Again, each member is described by a single, continuous parameter: the modal condition M. With this family of distributions, 62.5% of the young are distributed in the peak around the modal value (the width of which is 25% of the range of possible conditions). However, this modification does little to change the picture that emerged with the symmetric distribution family; at the nonsignaling equilibrium, both parties have higher fitness than at the signaling equilibrium (Fig. 4). With the present fitness functions, this result generalizes to broods with any number of siblings so long as the parents have perfect control of food allocation amongst chicks.
Figure 1.
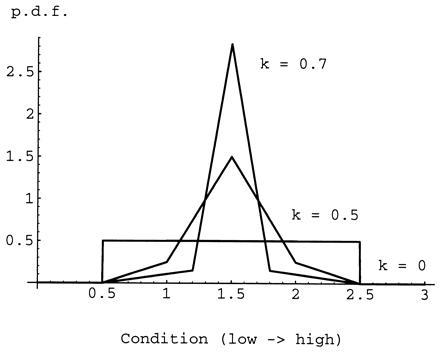
The probability density functions (p.d.f.) for three distributions of offspring condition. Higher values of the kurtosis index (k) result in a higher and narrower peak centered on the mean value.
Figure 2.
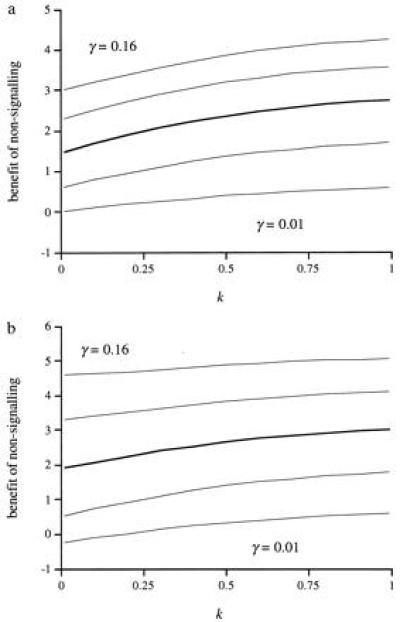
Percentage increase in inclusive fitness that (a) offspring and (b) parents derive from nonsignaling plotted against the kurtosis index (k) of the distribution of offspring conditions. The thick line represents the parameter values used by Godfray (3); U = 1, γ = 0.08, and V = 0.1. Thin lines are γ = 0.16, 0.12, 0.04, and 0.01.
Figure 3.
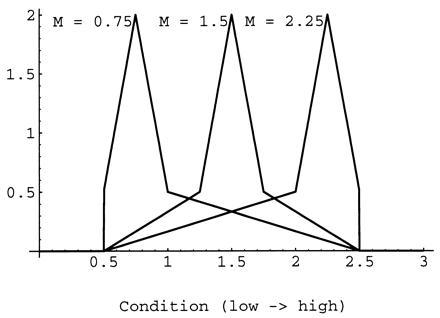
Probability density functions (p.d.f.) with different modal values (M).
Figure 4.
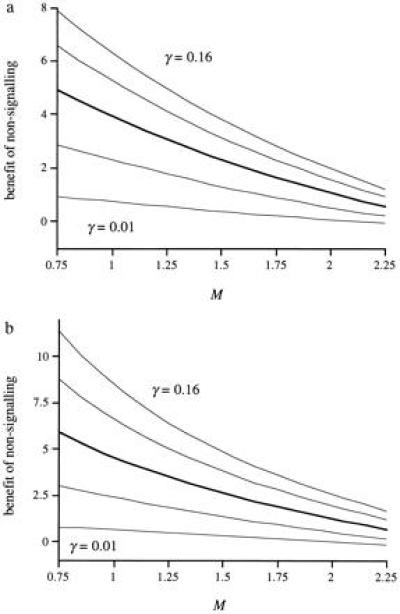
Percentage increase in inclusive fitness that (a) offspring and (b) parents derive from nonsignaling plotted against the mode of the distributions of offspring condition (M). Parameter values are as in Fig. 2.
Discussion
We have seen that, according to Godfray’s (3) model of signaling of need, there are two Nash equilibria: a signaling and a nonsignaling equilibrium. For most of the parameter values explored, the expected fitness for parents and their young was higher at the (ancestral) nonsignaling equilibrium than at the prevalent signaling equilibrium. If nothing else, the result that expected fitness is lower at the signaling than at the nonsignaling equilibrium highlights an important feature of the definition of signaling. Maynard Smith and Harper (13) define a signal as “an action or structure that increases the fitness of an individual by altering the behavior of other organisms detecting it, and that has characteristics that have evolved because they have that effect” (our italics). At least in the case of begging, signaling seems to have more to do with evolutionary stability than with increasing fitness.
Because parental care presumably evolved from a condition where fertilized eggs were released into the environment and not cared for, we must assume that the ancestral condition for food-soliciting species was nonsignaling. Therefore, it is a challenge to explain how and why these species shifted from the nonsignaling Nash equilibrium to the signaling one. It is important to bear in mind that this shift requires an explanation, regardless of the comparison between expected fitness at the signaling and the nonsignaling equilibria. In a way, the situation is similar to the common-sense and paradoxical ESS of the hawk–dove game with uncorrelated asymmetries (14): if, due to historical factors a species is trapped in a paradoxical ESS, it will normally not be able to jump to the common-sense ESS. Likewise with the signaling Nash equilibrium: by the very definition of a Nash equilibrium, a species should not be able to swap from one to the other. Changes from a high- to low-payoff equilibrium may be more counterintuitive than changes from a low- to high-payoff equilibrium, but they pose the same mathematical problem. What is compelling in this case is that we strongly believe that one equilibrium is ancestral to the other.
A possible scenario for the evolution of signaling of need is provided by the exploitation of sensory biases (15, 16). A strategy of the kind “if offspring does not signal treat it as an average offspring, if it does signal increase provisioning” would be selectively neutral in a population at the nonsignaling equilibrium and could spread by genetic drift. If the increase in provisioning was smaller or equal to that of the signaling-equilibrium strategy, this would be the end of the story: offspring would be favored by nonsignaling. If, on the other hand, the increase in provisioning of the mutant strategy was high enough, offspring might be able to increase their expected fitness by signaling at very low intensity. This contingency would trigger a cascade of events leading to the signaling equilibrium: chicks in higher need would be selected to signal with higher intensity, thus providing information to their parents about their state. Parents would then be selected to modulate their provisioning effort according to the signaling intensity of their offspring, and in due time the population would stabilize at the signaling equilibrium.
Although the exploitation of sensory biases provides a possible explanation for the evolution of signaling of need, it forces us to postulate an initial sensory bias so pronounced that parents in the ancestral population would have had to respond to the signals of their offspring more strongly than at the signaling Nash equilibrium. This is unlikely.
An alternative explanation can work without initial sensory biases. Food solicitations may have evolved in species rearing several offspring simultaneously, as a result of physical competition between siblings for the resources provided by the nondiscriminating parents. The argument goes thus: in the ancestral, nonsignaling condition, the parents would bring food to the nest according to availability and not according to chick behavior. Offspring can bias food allocation among themselves by monopolizing positions close to the feeding parent or by interfering in any other way in the feeding of their siblings. This is known to happen in the present day in both hole-nesting and open-nesting birds (for example, see refs. 17–19). As a result, young will compete with each other trying to increase their share of resources. Parents need not respond to this aggressive behavior for it to be selected: so long as the young have partial control over food allocation, this behavior will evolve. At the ESS, and assuming that all chicks have the same competitive ability, chicks will invest some energy into fighting their nestmates but will get as much food as if none of the chicks was fighting for food (20). This behavior of the chicks will select for an increase in the total amount of food the parents bring to the nest (21), but no communication has yet been established. We know, however, that the competitive ability of a chick should be reflected in the intensity with which it fights its nestmates at the ESS (22). As the expected payoff from being fed is condition-dependent, chicks in different condition will also fight their siblings with different intensities (23, 24). Hence, parents are receiving information about the condition and fighting ability of their offspring, and they will be selected to modulate investment in the brood and bias food so as to maximize their own fitness. A communicatory role for physical expressions of need has now emerged, and the system will converge to a signaling equilibrium. The exact properties of this equilibrium remain to be derived, but the above reasoning makes it clear that in multichick broods, so long as parents cannot avoid sibling competition, there is no nonsignaling Nash equilibrium. Theoretical models of begging have recently been extended to multichick scenarios (4). However, even these models restrict the possible strategies. They are “bidding games” in which each chick simultaneously makes a signal, after which the parent(s) decide how much food to bring and how to apportion it. Direct sibling competition by physical means (positional contests or overt aggression) is not included. In our opinion, direct sibling competition must have been an important component in the evolution of signaling of need, perhaps the main single factor that triggered the evolution of signaling from a nonsignaling equilibrium.
Acknowledgments
We thank Charles Godfray, Peter Hammerstein, Rufus Johnstone, David Krakauer, Arnon Lotem, and Martin Nowak for useful discussion and comments. M.A.R.-G. was funded by a F.P.U. grant from the Ministerio de Educación y Ciencia (Spain). P.A.C. is funded by Natural Environment Research Council (U.K.) Grant No. GR3/9239 to A.K.
Footnotes
Abbreviation: ESS, evolutionarily stable strategy.
References
- 1.Grafen A. J Theor Biol. 1990;144:517–546. doi: 10.1016/s0022-5193(05)80088-8. [DOI] [PubMed] [Google Scholar]
- 2.Grafen A. J Theor Biol. 1990;144:473–516. doi: 10.1016/s0022-5193(05)80087-6. [DOI] [PubMed] [Google Scholar]
- 3.Godfray H C J. Nature (London) 1991;352:328–330. [Google Scholar]
- 4.Godfray H C J. Am Nat. 1995;146:1–24. [Google Scholar]
- 5.Zahavi A. In: The Theory of Signal Selection and Some of Its Implications. Delfino V P, editor. Bari, Italy: Adriatica Edetricia; 1987. pp. 305–325. [Google Scholar]
- 6.McCarty J P. Auk. 1996;113:178–188. [Google Scholar]
- 7.Redondo T, Castro F. Ethology. 1992;92:193–204. [Google Scholar]
- 8.Haskell D. Proc R Soc London Ser B. 1994;257:161–164. [Google Scholar]
- 9.Zahavi A. J Theor Biol. 1977;67:146–151. doi: 10.1016/0022-5193(77)90061-3. [DOI] [PubMed] [Google Scholar]
- 10.Zahavi A. J Theor Biol. 1975;53:205–214. doi: 10.1016/0022-5193(75)90111-3. [DOI] [PubMed] [Google Scholar]
- 11.Aubin T. Behaviour. 1989;108:57–72. [Google Scholar]
- 12.Rudin W. Functional Analysis. New York: McGraw–Hill; 1991. [Google Scholar]
- 13.Maynard Smith J, Harper D G C. J Theor Biol. 1995;177:305–311. [Google Scholar]
- 14.Maynard Smith J, Parker G A. Anim Behav. 1976;24:159–175. [Google Scholar]
- 15.Ryan M J. In: Sexual Selection, Sensory Systems and Sensory Exploitation. Futuyma D J, Antonovics J, editors. Vol. 7. Oxford: Oxford University Press; 1990. pp. 157–195. [Google Scholar]
- 16.Krakauer D C, Johnstone R A. Philos Trans Roy Soc London B. 1995;348:355–361. doi: 10.1098/rstb.1995.0073. [DOI] [PubMed] [Google Scholar]
- 17.Teather K L. Behav Ecol Sociobiol. 1992;31:81–87. [Google Scholar]
- 18.McRae S B, Weatherhead P J, Montgomerie R. Behav Ecol Sociobiol. 1993;33:101–106. [Google Scholar]
- 19.Kacelnik A, Cotton P A, Stirling L, Wright J. Proc R Soc London Ser B. 1995;259:259–263. [Google Scholar]
- 20.Macnair M R, Parker G A. Anim Behav. 1979;27:1202–1209. [Google Scholar]
- 21.Parker G A, Macnair M R. Anim Behav. 1979;27:1210–1235. [Google Scholar]
- 22.Parker G A, Mock D W, Lamey T C. Am Nat. 1989;133:846–868. [Google Scholar]
- 23.Enquist M, Leimar O. J Theor Biol. 1987;127:187–205. [Google Scholar]
- 24.Rodríguez-Gironés M A, Drummond H, Kacelnik A. Behav Ecol. 1996;7:82–88. [Google Scholar]