

Abstract
The fungus Trichoderma harzianum is a potent mycoparasite of various plant pathogenic fungi. We have studied the molecular regulation of mycoparasitism in the host/mycoparasite system Botrytis cinerea/T. harzianum. Protein extracts, prepared from various stages of mycoparasitism, were used in electrophoretic mobility-shift assays (EMSAs) with two promoter fragments of the ech-42 (42-kDa endochitinase-encoding) gene of T. harzianum. This gene was chosen as a model because its expression is triggered during mycoparasitic interaction [Carsolio, C., Gutierrez, A., Jimenez, B., van Montagu, M. & Herrera-Estrella, A. (1994) Proc. Natl. Acad. Sci. USA 91, 10903–10907]. All cell-free extracts formed high-molecular weight protein–DNA complexes, but those obtained from mycelia activated for mycoparasitic attack formed a complex with greater mobility. Competition experiments, using oligonucleotides containing functional and nonfunctional consensus sites for binding of the carbon catabolite repressor Cre1, provided evidence that the complex from nonmycoparasitic mycelia involves the binding of Cre1 to both fragments of the ech-42 promoter. The presence of two and three consensus sites for binding of Cre1 in the two ech-42 promoter fragments used is consistent with these findings. In contrast, the formation of the protein–DNA complex from mycoparasitic mycelia is unaffected by the addition of the competing oligonucleotides and hence does not involve Cre1. Addition of equal amounts of protein of cell-free extracts from nonmycoparasitic mycelia converted the mycoparasitic DNA–protein complex into the nonmycoparasitic complex. The addition of the purified Cre1::glutathione S-transferase protein to mycoparasitic cell-free extracts produced the same effect. These findings suggest that ech-42 expression in T. harzianum is regulated by (i) binding of Cre1 to two single sites in the ech-42 promoter, (ii) binding of a “mycoparasitic” protein–protein complex to the ech-42 promoter in vicinity of the Cre1 binding sites, and (iii) functional inactivation of Cre1 upon mycoparasitic interaction to enable the formation of the mycoparasitic protein–DNA complex.
Biological control of plant pathogens is an attractive alternative to the strong dependence of modern agriculture on chemical fungicides, which cause environmental pollution and development of resistant strains. The saprophytic soil fungus Trichoderma harzianum Rifai is a potent mycoparasite of several economically important plant pathogenic fungi, and has therefore been tested for biocontrol potential in many field and greenhouse trials. Unfortunately, the efficacy of biocontrol strains has shown to be, in most of the cases, less predictable and not competitive with that of chemical fungicides. Knowledge of the biochemical events that determine mycoparasitism and their regulation would suggest strategies to improve the reliability of T. harzianum as a biocontrol agent. To accomplish this goal, a number of genes, specifically induced under mycoparasitic conditions, have been cloned from T. harzianum (1–9). Most of these genes encode chitinolytic and glucanolytic enzymes (2–4, 6, 8, 9); this reflects the need of the mycoparasite to penetrate the cell wall of the plant pathogen, which, in many cases, consists of chitin and β-1,3-glucans as major structural components. Chitinases have been proven essential for biocontrol in vivo and in vitro, and fungal chitinase-encoding genes are therefore considered as attractive tools to reinforce plant defenses (10, 11).
Despite this importance of chitinases, there have been few attempts to understand the regulation of chitinase expression in T. harzianum. ech-42 transcription is induced by chitin and repressed by glucose (3, 4), but these data were obtained with T. harzianum growing in submerged culture on chitin or cell walls of plant pathogenic fungi, which do not reflect natural conditions.
Because T. harzianum is not capable of sexual reproduction, genetically defined mutants that could be used to study gene regulation are not available and cannot be easily obtained. Therefore, we have used an alternative approach to obtain information on the regulation of ech-42 expression under mycoparasitic conditions: we studied proteins binding to the ech-42 promoter in vitro by using cell-free extracts from mycelia of T. harzianum during the mycoparasitic interaction with mycelia of Botrytis cinerea, an important plant pathogen. Protein–DNA complexes, whose binding is competitively preempted by the carbon catabolite regulator Cre1 and favored under mycoparasitic conditions, were detected.
MATERIALS AND METHODS
Strains and Plasmids.
T. harzianum strain P1 (ATCC 74058), isolated from wood chips, and B. cinerea Pers.:Fr.strain 12 isolated from grapes were used as a mycoparasite and its host, respectively, since P1 is able to limit the activity of B. cinerea as a pathogen on apple, grapes and other crops (12). Plasmids pBluescript (Stratagene) and pCRII (Invitrogen) were used for cloning PCR-generated fragments.
Plate Confrontation Assays and Protein Extracts.
Plate confrontation assays, where the mycelia of T. harzianum and B. cinerea were allowed to intermingle, were performed in the dark on potato dextrose agar or minimal medium with 2% agar plus 0.3% glucose (3). For extraction of DNA-binding proteins, T. harzianum and B. cinerea were grown on confrontation plates, and mycelia of T. harzianum collected at different stages of interaction: (i) 2 days after inoculation before contact with B. cinerea (Trichoderma or Botrytis mycelia, 5 mm apart from the original inoculum and without spores were collected); (ii) 12 h, (iii) 24 h, and (iv) 72 h after contact with mycelium of B. cinerea; and (v) 12–24 h after hyphal contact with mycelium of another colony of T. harzianum strain P1 of the same age. Mycelium of B. cinerea was also collected immediately after contact with T. harzianum. All the mycelia in the area of interaction or close to it were recovered. Equivalent zones were collected from control plates inoculated only with T. harzianum or B. cinerea. T. harzianum was also grown in liquid culture on minimal medium plus 0.3% glucose or malt extract, and the mycelium was collected by centrifugation 4 days after the cultures were started. Cell-free extracts were obtained as described (13) and stored at −70°C. A glutathione S-transferase (GST)::Cre1 fusion protein from Trichoderma reesei E. Simmonds was used to investigate the competitive effect of Cre1 on the binding of cell-free extracts to the promoter fragments. The preparation of this fusion protein has been described elsewhere (14); briefly, a 700-bp DNA fragment encoding the DNA-binding domain of the Cre1-protein was expressed as a GST::Cre1 fusion in Escherichia coli LC137, and the protein purified as described in the manufacturer protocols (Pharmacia–LKB). Molecular biological standard techniques were carried out according to Sambrook et al. (15).
Cloning of the ech-42 Promoter.
A 829-bp fragment, comprising nucleotides −1 to −829 from the start codon of ech-42, was PCR amplified from genomic DNA of T. harzianum. Three nested primers were designed for the region containing the 5′ end of ech-42 (ref. 2; Table 1); namely: E1, which anneals at nucleotide +133; E2, which anneals upstream of E1 and includes the start codon; and E3, which anneals at nucleotide −60. PCR conditions were as follows: one cycle of 3 min at 94°C and of 1 min at 55°C, followed by 34 cycles of 1 min at 94°C; 1 min at 55°C and 3 min at 72°C; and a final cycle of 1 min at 94°C, 1 min at 55°C, and 7 min at 72°C. E1 and random primer 221 (Table 1) were first used to amplify a putative promoter fragment, which was identified by probing with a 700-bp fragment of ech-42 containing the 5′ end of the gene. The band hybridizing with the probe was purified from the gel and re-amplified first with oligonucleotides E2 plus 221 and then with E3 plus 221. Two amplicons showing the expected sizes (803 and 829 bp, respectively) were purified from the gel and subcloned. At least two PCR products, obtained independently from different primers and combinations, were cloned in pCRII and sequenced from both ends. The final sequence was also verified by sequencing two overlapping fragments of the promoter, which were obtained by cleavage with BamHI and XbaI or with SalI and EcoRV.
Table 1.
Oligonucleotide used for PCR amplification or for EMSA
| Name | Sequence | Ref. |
|---|---|---|
| 221 | 5′-aatcgggctg-3′ | This study |
| E1 | 5′-ctggctctcttctcaaca-3′ | This study |
| E2 | 5′-atggtgaagtgtttgaga-3′ | This study |
| E3 | 5′-ttgaaaggaagaagttgc-3′ | This study |
| Cre1so | 5′-atattatgcggagaccccagaatgtttctcc-3′ | 14 |
| Cre1wt | 5′-gccggatgcaccccagatctggggaacgcgccgc-3′ | 14 |
| Cre1mu | 5′-gccggatgcacccaagatcttgggaacgcgccgc-3′ | 14 |
EMSA, electrophoretic mobility-shift assay.
Preparation of DNA Fragments EMSA.
Three fragments of the promoter of the T. reesei xyn1 (xylanase I-encoding) gene generated by restriction of a 538-bp DNA fragment (14) were also used for EMSA. The fragments used for EMSA were end-labeled with the appropriate [γ-32P]dNTP using Sequenase version 2.0 (United States Biochemical). Synthetic double-stranded oligonucleotides, used for competition experiments, are given in Table 1, and were prepared as described by Strauss et al. (14). The DNA–protein binding and the EMSA were performed as described by Stangl et al. (13). DNA–protein binding was performed at 4°C for 5 min and extracts obtained from different mycoparasitism stages were always tested at the same protein concentration for each experiment or mixed at different ratios.
Electrophoresis.
Samples to be analyzed by SDS/PAGE were boiled in SDS/PAGE buffer and electrophoresed in 10% acrylamide gels. Bands were visualized by Coomassie blue staining.
RESULTS
Mycoparasitic Interaction Is Accompanied by a Major Change in the Intracellular Protein Composition of the Mycoparasite.
Cell-free extracts containing nuclear proteins were prepared from T. harzianum strain P1 at various stages of interaction with Botrytis cinerea—i.e., 2 days before and 12, 24, and 72 h after contact with B. cinerea (Fig. 1A). During the latter stages, triggering of expression of ech-42 biosynthesis occurred (ref. 3, and data not shown) and the mycelium of host lysis was completed after 72 h. SDS/PAGE showed that the mycoparasitic process was paralleled by a striking change in the protein composition of the extracts (Fig. 1B). The major changes were observed already 12 h after the initial contact and little variation occurred thereafter. No alterations in the SDS/PAGE pattern were observed with protein extracts from mycelia after prolonged growth in either liquid or solid media, and in mycelia of T. harzianum confronted with other mycelia of T. harzianum (Fig. 1B and data not shown). Therefore, regulatory events triggering mycoparasitism probably take place within the first 12 h of interaction.
Figure 1.
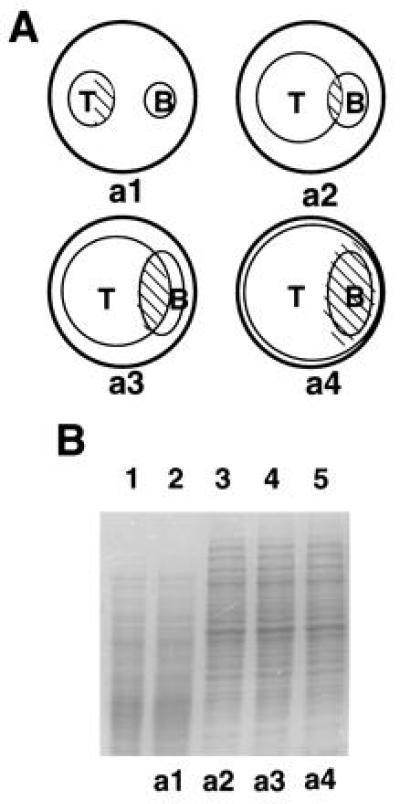
Plate confrontation assay of T. harzianum and B. cinerea, and SDS/PAGE of cell-free extracts of mycelia collected during different stages of mycoparasitism. (A) Schematic description of the plate assay used to prepare cell-free extracts from different stages of mycoparasitism. Small circles represent T. harzianum (T) or B. cinerea (B) colonies growing on agar medium in a Petri plate (large circles). The mycelium was collected from the pattern-filled zones before the contact (a1), and 12 h (a2), 24 h (a3), and 72 h (a4) after the contact. Samples were also collected from other conditions (see Materials and Methods). (B) SDS/PAGE of T. harzianum cell-free extracts from different conditions. Lanes: 1, extract from a T. harzianum–T. harzianum interaction 12–24 h after the contact; 2, T. harzianum extract from mycelium in stage a1; and 3–5, T. harzianum extracts of mycelia from stages a2, a3, and a4, respectively. Twenty micrograms of protein was applied per slot.
Cloning and Sequence Analysis of ech-42 5′-Upstream Sequences.
To investigate the alterations in the interaction of proteins with the regulatory regions of mycoparasitism genes during shift from mycelial growth to mycoparasitism, ech-42 was used as a model. The expression of this gene during transition to mycoparasitism has already been shown (3) under experimental conditions similar to the ones used here. Therefore, a promoter fragment of the ech-42 gene encoding for an endochitinase was obtained by PCR from T. harzianum genomic DNA using a nested primer from the 5′ end of the encoding sequence and a random primer. The larger band, which was 829 bp, was purified and sequenced. The sequence was identical to the already known (3) 350 bp of the ech-42 promoter of T. harzianum, with the exception of one bp at nucleotide −327 (C instead of T). The 829 bp (Fig. 2) displayed several single consensus sites (5′-SYGGRG-3′; refs. 16 and 17) for binding of the carbon catabolite repressor CreA (which corresponds to the Trichoderma Cre1; ref. 14). No other known consensus sites for transcriptional regulators could be identified with certainty.
Figure 2.

Sequence of the ech-42 promoter region. The TATA boxes are printed in boldface type and underlined. Putative consensus sequences for binding of Cre1 are underlined, and the arrow indicates their positions on the coding strand (→) and on the complementary strand (←).
T. harzianum Proteins Form Mycoparasitism-Dependent Complexes with ech-42 Nucleotide Fragments.
Two SalI fragments S1 and S2 (480 and 349 bp, respectively) of the ech-42 5′-end noncoding sequences were analyzed by EMSA for their ability to bind proteins present in extracts from different stages of the mycoparasitic process. Two different high-molecular-weight DNA–protein complexes were found with cell-free extracts from mycoparasitic and nonmycoparasitic interaction, respectively. Of these two the DNA–protein complex observed under mycoparasitic conditions exhibited greater mobility than that formed under nonmycoparasitic conditions (Fig. 3). Controls using extracts from T. harzianum confronted with itself did not show the shift in size of the complex typical for mycoparasitism, indicating that its appearance was not the result of nutrient (i.e., glucose) starvation. The formation of DNA-protein complexes with cell-free extracts of B. cinerea likewise was not responsible for the “mycoparasitic shift,” because the controls showed that this complex did not penetrate into the gel under the conditions applied here. We therefore conclude that mycoparasitic conditions are characterized by an alteration in the binding of proteins to the ech-42 promoter.
Figure 3.
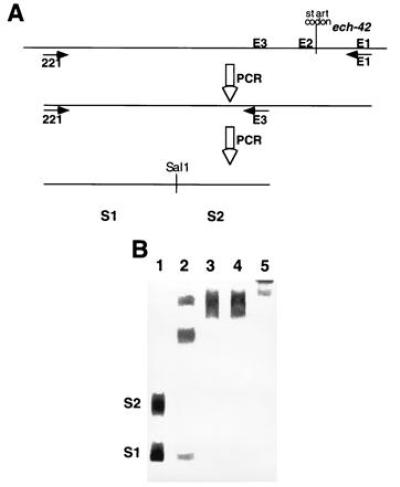
In vitro binding of proteins to fragments of the ech-42 promoter. (A) Schematic representation of the method used to prepare the DNA fragments for EMSA. PCR was performed with combinations of primers 221, E1, E2, and E3 as described. The final 221 plus E3 amplification product was sequenced and digested with SalI to obtain the two fragments used for EMSA (S1 and S2). (B) EMSA, using protein extracts of T. harzianum and two fragments of the ech-42 promoter region. Lanes: 1, free DNA; 2, DNA plus extract from a3; 3, DNA plus extract from a1; 4, DNA plus protein extracts from the mycelium of a T. harzianum–T. harzianum interaction; and 5, DNA plus protein extracts from mycelium of B. cinerea during a2.
The Nonmycoparasitic DNA–Protein Complex Involves the DNA-Binding of Cre1.
Since the two ech-42 promoter fragments showed the presence of consensus sequences for binding of the carbon catabolite repressor protein Cre1, we investigated whether these sites are involved in the formation of the DNA–protein complexes observed with cell-free extracts obtained from mycelia grown under mycoparasitism or nonmycoparasitism conditions, respectively. To this end, increasing amounts of an unlabeled double-stranded fragment of the xyn1 promoter of T. reesei, containing a double and a single Cre1 binding site (Cre1so; ref. 14), were added to the EMSA-incubation mixtures. Fig. 4 shows that Cre1so competitively inhibited the binding of the extracts from nonmycoparasitic, but not that from the mycoparasitic mycelia, to ech-42 promoter fragments. Complete inhibition was obtained with a molar ratio of 1:50 (genomic DNA vs oligonucleotide, respectively). To demonstrate the specific involvement of Cre1, another oligonucleotide (Cre1mu), which contains a mutation in the Cre1 binding site that substantially reduces the binding of Cre1 in vitro and in vivo (14, 18), was used (Fig. 4B). No competition was observed, even with 100 molar excess of Cre1mu. These results clearly indicate that the formation of nonmycoparasitic, but not the mycoparasitic, DNA–protein complex involves binding of Cre1 to ech42.
Figure 4.
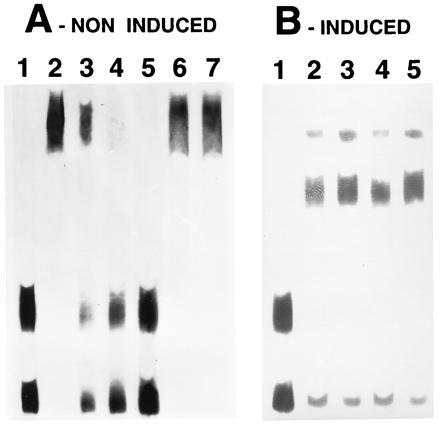
Demonstration of the involvement of Cre1 in protein–DNA binding during EMSA with protein extracts of T. harzianum and the ech-42 fragments S1 and S2. Two oligonucleotides, one containing a double Cre1-binding site (Cre1wt) and one containing the same site mutated and unable to bind Cre1 (Cre1mu), were used. (A) Lanes: 1, free DNA; 2, DNA plus protein extracts from a1 (see Fig. 1); 3–5, DNA plus protein extracts from a1 in the presence of a 5-fold (lane 3), 10-fold (lane 4), and 50-fold (lane 5) molar excess of Cre1wt; and 6 and 7, DNA plus protein extracts from a1 in the presence of a 50-fold (lane 6) and 100-fold (lane 7) molar excess of Cre1mu. (B) Lanes: 1, free DNA; 2, DNA plus protein extracts from a3 (see Fig. 1); 3 and 4, DNA plus protein extracts from a3 in the presence of a 50-fold (lane 3) and 100-fold (lane 4) molar excess of Cre1wt; and 5, DNA plus protein extracts from a3 plus a 100-fold molar excess of Cre1mu.
Mycoparasitic Interaction Prevents Cre1-Binding to its Target Sequence in the ech-42 Promoter.
To obtain information on how binding of the mycoparasitism-specific protein complex takes place at the expense of Cre1-binding, protein extracts from both the mycoparasitic and the nonmycoparasitic conditions were mixed in various proportions and binding to ech-42 analyzed by EMSA. The presence of equal amounts of protein from both types of extracts yielded the nonmycoparasitic protein–DNA complex, which, therefore, at similar protein concentrations, displayed a higher stability than the mycoparasitic protein–DNA complex (Fig. 5). To find out whether the protein-DNA complex formed under mycoparasitism conditions requires functional inactivation of Cre1, varying amounts of a GST::Cre1 fusion protein (14) were added to the extract from mycoparasitic mycelia. As shown (Fig. 5), the fusion protein strongly competed with these protein extracts. Controls showed that the GST::Cre1 fusion protein alone formed complexes of a size similar to that of complexes from nonmycoparasitic cell-free extracts. The simultaneous addition of GST::Cre1 fusion protein and of protein extracts from nonmycoparasitic mycelia synergistically inhibited formation of the mycoparasitic complex. These data indicate that Cre1 successfully competes with the “mycoparasitic” DNA-binding protein(s) for both ech-42 fragments, suggesting that Cre1 must become functionally impaired during transition to mycoparasitism.
Figure 5.
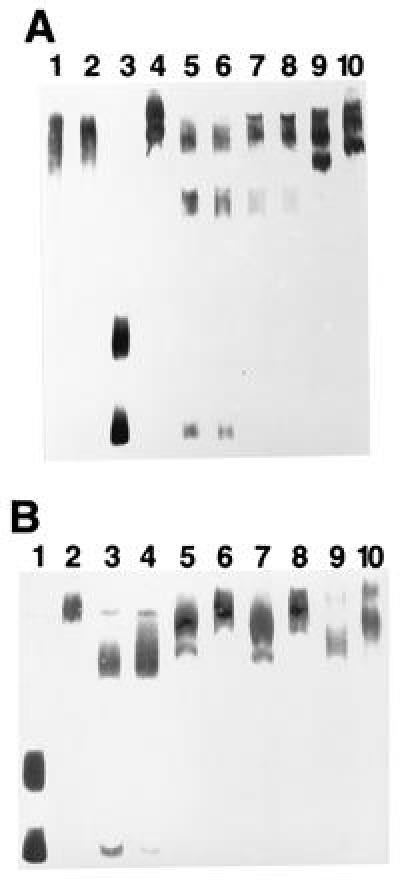
EMSA of protein extracts of T. harzianum with fragments S1 and S2 of the ech-42 promoter in the presence of GST::Cre1 fusion protein of T. reesei. (A) Lanes: 1, DNA plus protein extract from a1 (see Fig. 1) in the presence of 10 μg of GST::Cre1; 2, as in line 1 but in the absence of Cre1::GST; 3, free DNA; 4, DNA plus 10 μg of GST::Cre1 in the absence of protein extracts from T. harzianum; 5, DNA plus protein extracts from a3 (see Fig. 1); 6–10, DNA plus cell-free extracts from a3 in the presence of 0.01 (lane 6), 0.1 (lane 7), 0.5 (lane 8), 1.0 (lane 9), and 10 μg (lane 10) of GST::Cre1. (B) EMSA with DNA and various ratios of protein extracts from a1 and a3. Lanes: 1, free DNA; 2, protein extracts from a1; 3, protein extracts from a3; 4–6, extracts from a1 and a3, added at a protein ratio (wt/wt) of 5:1 (lane 4), 1:1 (lane 5), and 1:5 (lane 6); 7, protein extracts from a1 and a3 at a ratio of 5:1 in the presence of 0.1 μg of GST::Cre1; 8, protein extracts from a1 and a3 at a ratio of 3:1 in the presence of 0.1 μg of GST::Cre1; and 9 and 10, EMSA with 0.1 (lane 9) and 0.3 μg (lane 10) of GST::Cre1 only.
DISCUSSION
The results presented in this paper provide insight into the molecular mechanism triggering mycoparasitism. Using ech-42, a gene expressed in hyphae of T. harzianum upon contact with its host (3), we show that this mycoparasitic interaction is accompanied by a change in the ability of nuclear proteins of the fungus to bind to the ech-42 promoter. Two different protein–DNA complexes, corresponding respectively to mycelia before and during mycoparasitism, were found with cell-free extracts. In the case of nonmycoparasitic mycelia, competition analysis showed that the carbon catabolite repressor Cre1 participates in this DNA–protein complex. At least five consensus-target sites for Cre1 were detected in the 5′-upstream sequences of ech-42. Further studies, including analysis of deletions, will be undertaken to show which of those sites is actually functional in vivo.
Upon contact of T. harzianum with its host, a new DNA–protein complex was formed with the ech-42 promoter, whose binding could not be competitively inhibited with Cre1-specific oligonucleotide sequences. This indicates that during the first 12 h of mycelial interaction, the Cre1-containing protein–DNA complex characteristic of the nonparasitic condition is displaced by a “mycoparasitism-specific” complex. Nothing is yet known about the target sequences for this complex. The fact that formation of this complex could be competitively inhibited by the addition of the Cre1::GST fusion protein suggests that both complexes may have overlapping or contiguous target sequences. However, it is equally possible that proteins, specifically present during the mycoparasitic and nonmycoparasitic conditions, respectively, compete for binding to a protein component of the TATA-binding protein–RNA polymerase II complex rather than directly for a DNA target. Interestingly, the formation of the mycoparasitism-specific protein–DNA complex obviously requires a functional inactivation of Cre1, because the addition of Cre1 to cell-free extracts from mycoparasitic mycelia replaces the mycoparasitism-specific protein–DNA complex with the Cre1-specific protein–DNA complex. Unfortunately, the regulation of Cre1 function is still not understood; there is evidence that its transcription is constitutive and autoregulated (19) and its function in vivo may therefore be regulated by posttranslational modification. The possible occurrence of such a regulation (i.e., by phosphorylation) has been shown for the yeast counterpart of Cre1–Mig1p (20).
Based on the present results, we present a model which—albeit still hypothetical—accounts for all the observations made, and which should be useful to design experiments to study in vivo the mechanism that triggers expression of ech-42 and other mycoparasitism genes in T. harzianum upon contact with its host (Fig. 6). Before contact with the plant pathogen, Cre1 binds to its target sequence (RSA) and precludes binding of the mycoparasitism regulator to RSB, which has lower affinity to its target than that of Cre1 (Fig. 6A). Upon contact with the plant pathogen, Cre1 becomes functionally impaired and dissociates from RSA, thereby enabling the mycoparasitism regulator to bind to RSB (Fig. 6B). This model bears striking similarity to the regulation of alcR expression in Aspergillus nidulans (21), where the regulatory sequences for binding of CreA and the positive autoregulator AlcR are so closely located that the occupation by the respective protein precludes binding of the other. Furthermore, it has been demonstrated that, under glucose-repressed conditions, CreA has a higher affinity for its target than AlcR (21). Since the relevance of the A. nidulans/alcR model to the in vivo situation has been established, we expect, by analogy, that our model based on in vitro investigations also reflects the in vivo situation.
Figure 6.
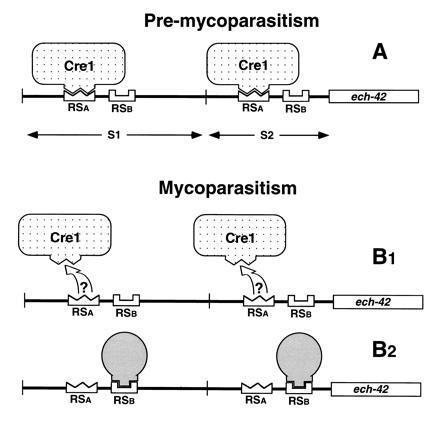
Model to explain the regulation of the expression of ech-42 during mycoparasitism. RSa, target sequences for binding of Cre1-specific complexes (nonmycoparasitic); RSb, target sequences for binding of the mycoparasitic regulator. S1 and S2 = SalI fragments of the ech-42 5′-upstream sequence (see Discussion).
Note Added in Proof.
While this manuscript was in press, we became aware of a paper by Margolles-Clark et al. (22) reporting on the effect of different carbon sources on ech-42 expression in T. harzianum.
Acknowledgments
This work was supported in part by a grant of the Austrian Science Foundation (P 10793-GEN) and by the Austrian Ministry of Science, Research, and Arts (Contract GZ 49.953/1-Pr/4/95) to C.P.K. and by the Italian Ministry of Agriculture (Project “Biotecnologie Vegetale”) to M.L. M.L. was a collaborator under a fellowship from the Organization for Economic and Cooperative Development Cooperative Research Programme: Biological Resource Management for Sustainable Agricultural Systems, which is gratefully acknowledged.
Footnotes
References
- 1.Geremia R A, Goldman G H, Jacobs D, Ardiles W, Via S B, Van Montagu M, Herrera-Estrella A. Mol Microbiol. 1993;8:603–613. doi: 10.1111/j.1365-2958.1993.tb01604.x. [DOI] [PubMed] [Google Scholar]
- 2.Hayes C K, Klemsdal S, Lorito M, Di Pietro A, Peterbauer C K, Nakas J P, Tronsmo A, Harman G E. Gene. 1994;138:143–148. doi: 10.1016/0378-1119(94)90797-8. [DOI] [PubMed] [Google Scholar]
- 3.Carsolio C, Gutierrez A, Jimenez B, van Montagu M, Herrera-Estrella A. Proc Natl Acad Sci USA. 1994;91:10903–10907. doi: 10.1073/pnas.91.23.10903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Garcia I, Lora J M, de la Cruz J, Benitez T, Llobell A, Pintor-Toro J A. Curr Genet. 1994;27:83–89. doi: 10.1007/BF00326583. [DOI] [PubMed] [Google Scholar]
- 5.Goldman G H, Vasseur V, Contreras R, Van Montagu M. Gene. 1994;144:113–117. doi: 10.1016/0378-1119(94)90213-5. [DOI] [PubMed] [Google Scholar]
- 6.Lora J M, de la Cruz J, Benitez T, Llobell A, Pintor-Toro J A. Mol Gen Genet. 1994;242:461–466. doi: 10.1007/BF00281797. [DOI] [PubMed] [Google Scholar]
- 7.Vasseur V, Van Montagu M, Goldman G. Microbiology (Reading, UK) 1995;141:767–774. doi: 10.1099/13500872-141-4-767. [DOI] [PubMed] [Google Scholar]
- 8.Limon M C, Lora J M, Garcia I, de la Cruz J, Benitez T, Llobell A, Pintor-Toro J A. Curr Genet. 1995;28:478–483. doi: 10.1007/BF00310819. [DOI] [PubMed] [Google Scholar]
- 9.Lora J M, De la Cruz J, Benitez T, Pintor-Toto J A. Mol Gen Genet. 1995;247:639–647. doi: 10.1007/BF00290356. [DOI] [PubMed] [Google Scholar]
- 10.Chet I, Schickler H, Haran S, Appenheim A B. In: International Symposium of Chitin Enzymology. Mozzarelli R, editor. Italy: Senigallia; 1993. pp. 47–48. [Google Scholar]
- 11.Sundheim L. In: Biological Control of Plant Diseases. James E C, Papavizas G C, Cook R J, editors. New York: Plenum; 1992. pp. 331–332. [Google Scholar]
- 12.Tronsmo A. Biol Control. 1991;1:59–62. [Google Scholar]
- 13.Stangl H, Gruber F, Kubicek C P. Curr Genet. 1993;23:115–122. doi: 10.1007/BF00352009. [DOI] [PubMed] [Google Scholar]
- 14.Strauss J, Mach R L, Zeilinger S, Hartler G, Stöffler G, Wolschek M, Kubicek C P. FEBS Lett. 1995;376:103–107. doi: 10.1016/0014-5793(95)01255-5. [DOI] [PubMed] [Google Scholar]
- 15.Sambrook J, Fritsch E F, Maniatis T. Molecular Cloning: A Laboratory Manual. 2nd Ed. Plainview, NY: Cold Spring Harbor Lab. Press; 1989. [Google Scholar]
- 16.Kulmburg P, Mathieu M, Dowzer C, Kelly J, Felenbok B. Mol Microbiol. 1993;7:847–857. doi: 10.1111/j.1365-2958.1993.tb01175.x. [DOI] [PubMed] [Google Scholar]
- 17.Cubero B, Scazzocchio C. EMBO J. 1994;13:407–415. doi: 10.1002/j.1460-2075.1994.tb06275.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Mach R L, Zeilinger S, Strauss J, Schindler M, Kubicek C P. Mol Microbiol. 1996;121:1273–1281. doi: 10.1046/j.1365-2958.1996.00094.x. [DOI] [PubMed] [Google Scholar]
- 19.Ilmen M, Thrane C, Penttilä M. Mol Gen Genet. 1996;251:451–460. doi: 10.1007/BF02172374. [DOI] [PubMed] [Google Scholar]
- 20.Treitel M A, Carlson M. Proc Natl Acad Sci USA. 1995;92:3132–3136. doi: 10.1073/pnas.92.8.3132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Mathieu M, Felenbok B. EMBO J. 1994;13:4022–4027. doi: 10.1002/j.1460-2075.1994.tb06718.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Margolles-Clark E, Harman G E, Penttilä M. Appl Environ Microbiol. 1996;62:2152–2155. doi: 10.1128/aem.62.6.2152-2155.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]