

Abstract
We have isolated the promoter region and determined the start sites of transcription for the gene encoding the chicken m2 (cm2) muscarinic acetylcholine receptor. Transfection experiments, using cm2-luciferase reporter gene constructs, demonstrated that a 789-bp genomic fragment was sufficient to drive high level expression in chicken heart primary cultures, while an additional 1.2-kb region was required for maximal expression in mouse septal/neuroblastoma (SN56) cells. Treatment of SN56 cells with the cytokines ciliary neurotrophic factor and leukemia inhibitory factor increases expression of endogenous muscarinic acetylcholine receptors and results in a 4- to 6-fold induction of cm2 promoter driven luciferase expression. We have mapped a region of the cm2 promoter that is necessary for induction by cytokines.
The muscarinic acetylcholine receptors (mAChR) belong to the superfamily of receptors, predicted to contain seven trans-membrane domains, whose biological actions are elicited via activation of G proteins (1). Five different mammalian subtypes have been identified by molecular cloning and are encoded by separate genes (2, 3). The m1, m3, and m5 subtypes preferentially couple to the stimulation of phospholipase C, and the m2 and m4 subtypes preferentially couple to the inhibition of adenylate cyclase. Four chicken mAChRs have been cloned to date: cm2, cm3, cm4, and cm5 (4–7).
The m2 subtype is the main mAChR found in mammalian heart (8), while chicken heart expresses predominantly cm2 with significant amounts of cm4 and cm3 (9). Cardiac mAChR activation causes a decrease in both the rate and force of contraction (10). These effects are elicited through the inhibition of adenylate cyclase activity, activation of an inward-rectifying potassium channel, inhibition of a calcium channel, and inhibition of the hyperpolarization-activated pacemaker current.
In the mammalian central nervous system mRNA encoding the m2 receptor and m2-specific immunoreactivity have been localized to basal forebrain/septum, striatum, hippocampus, hypothalamus, amygdala, and pontine nuclei regions. Furthermore, choline acetyltransferase (ChAT) immunoreactivity has been colocalized to subpopulations of m2-receptor expressing cells in some of these regions and activation of muscarinic receptors can result in presynaptic inhibition of acetylcholine release (see ref. 11 for review). In rat neostriatal cholinergic interneurons, activation of the m2 receptor results in reduction of N- and P-type Ca2+ currents (12) that regulate neurotransmitter release (13), potentially through a G protein βγ-dependent mechanism (14). Postsynaptic cells can be hyperpolarized in response to m2 activation. For example, cells of the rat nucleus raphe magnus are hyperpolarized by an inwardly rectifying potassium current in response to m2 activation (15). Four G protein-coupled inward rectifying K+ channel genes have been cloned from mammalian brain that form heteromultimeric functional channels (16–19). Muscarinic receptor activation of the inwardly rectifying K+ current most likely occurs through these channels.
We have isolated and begun to characterize the promoter region of the cm2 gene to determine the molecular mechanisms involved in its transcriptional regulation. We demonstrate here the transcriptional activity of cm2 promoter constructs in both primary chicken heart cultures and a mouse septal/neuroblastoma cell line SN56, the induction of cm2 reporter constructs by cytokines in neural cells and the mapping of the region of the cm2 promoter responsible for cytokine regulation.
METHODS
Isolation of cm2 cDNA Clones.
A chicken heart λ ZapII (Stratagene) cDNA library was screened with the third intracellular loop from the cm2 gene (4) using standard techniques (20). After three rounds of plaque purification positive clones were excised into pBluescript SK−, according to the manufacturer’s protocol, and subjected to sequence analysis using Sequenase (United States Biochemical) and Applied Biosystems Taq Dye Terminator Sequence kit.
Isolation of cm2 Genomic Clones.
A λFix (Stratagene) chicken genomic library was screened using a 54-base oligonucleotide (NNMR187; see Fig. 1), labeled to a specific activity of 5.0 × 108 cpm/μg with 32P, under the following conditions. Prehybridization and hybridization were carried out in 6× standard saline phosphate/EDTA (0.18 M NaCl/10 mM phosphate, pH 7.4/1 mM EDTA), 5× Denhardt’s reagent, 1% SDS, 50% formamide, 250 μg/ml salmon sperm DNA at 37°C. Filters were washed in 2× standard saline phosphate/EDTA, 0.1% SDS for 2 hr at room temperature then 1 hr at 56°C followed by 20 min at 63°C. After three rounds of plaque purification DNA was isolated from positive phage and subjected to Southern blot analysis using NNMR187. The phage inserts were mapped; several overlapping fragments were subcloned into pBluescript SK+ (Stratagene) and subjected to sequence analysis as described above.
Figure 1.
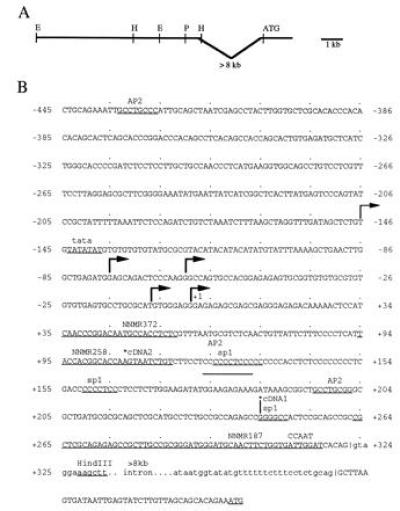
Partial restriction map, genomic organization and nucleotide sequence of cm2 gene. (A) Partial restriction map of the 8 kb sequence upstream of intron one in cm2. E, EcoRI; H, HindIII; P, PstI. The position and estimated size of intron one is indicated along with the initiator methionine. (B) Nucleotide sequence of the 789-bp PstI/HindIII fragment upstream, and sequences downstream of intron one are shown. Nucleotides are numbered from the most 3′ start site of transcription. Nucleotides in intron one are in lower case letters. Start sites of transcription are shown with arrows. Start sites determined by both primer extension and RNase protection are shown as heavy arrows while a start site determined by primer extension only is shown as thin arrow. Putative transcription factor binding sites and HindIII site are underlined below and labeled above the sequence. Positions of oligonucleotides used for cloning, making of constructs and primer extension analysis are also underlined below and labeled above sequence. End of cm2 cDNAs are indicated with an asterisk. Intron splice sites are shown with vertical line.
Cell Culture.
Primary chicken heart cultures were prepared from 9-day embryos and grown in defined media as described (21). In general, five to six dozen eggs were dissected and cardiac cells plated out in six to seven 24-well plates at density of 3–4 × 105 cells per well. SN56 cells (22) were grown in DMEM supplemented with 10% fetal bovine serum in a humidified 10% CO2 incubator.
Cell Transfection.
Primary chicken heart cultures were transfected three days after plating using Transfectam (Promega) according to the manufacturer’s protocol. Because of the wide range in size of the cm2-luciferase reporter gene constructs used, cells were transfected with a constant molar amount of reporter construct. The transfection mixture contained 0.037 pmols per well of reporter construct (134–320 ng), brought to a final amount of 320 ng per well with pGEM4 (Promega) as carrier DNA, and 100 ng per well of pSV-β-galactosidase (β-gal) (Promega). Transfectam reagent was used at a ratio of 3.5 μl/μg of DNA. The transfection cocktail was added to cells for 6–7 hr, then removed and fresh media was added. Cells were lysed 30 hr after transfection with 100 μl lysis buffer (23). Luciferase and β-gal activities were determined as described (23). SN56 cells were transfected using the calcium-phosphate procedure. Briefly, cells were plated into 24-well plates at 30,000 cells per well and transfected 24 hr later. The transfection cocktail contained 0.048 pmols of reporter construct (174–420 ng) combined with pGEM4 to a total amount of 420 ng per well, and 100 ng per well of pRSV-β-gal (24). Sixteen hours after addition of transfection cocktail, cells were treated for 32 hr with cytokines, and lysed as described above. Luciferase and β-gal activities were determined as described (23).
DNA Constructs and Oligonucleotides.
pNMR26: A 789-bp PstI/HindIII cm2 promoter fragment was subcloned into pBluescript SK+ (Stratagene) to make pNMR11. The insert was cut out using HindIII/SacI and ligated into the luciferase reporter gene vector pGL3-Basic (Promega). pNMR27: A 2-kb EcoRI/HindIII cm2 promoter fragment was subcloned into pBluescript SK+ to make pNMR12. The insert was cut out with HindIII/SacI and ligated into pGL3. pNMR28: A 3.3-kb HindIII cm2 promoter fragment was subcloned into pGL3. pNMR29: The same 3.3-kb HindIII cm2 promoter fragment was subcloned into pBluescript SK+ with the 3′ end of the cm2 promoter region proximal to the vectors T7 promoter. The resulting plasmid, pNMR4, was digested with EcoRI and a 4.3-kb fragment, containing the vector, was isolated. A 6-kb EcoRI fragment was inserted to make pNMR16. The plasmid pNMR16 was digested with XhoI/SpeI and ligated into pGL3 digested with XhoI/MluI using the adapter oligonucleotides NNMR262 and NNJJ 138 to make pNMR29. pNMR25: NNMR258 and the M13 reverse oligonucleotide were used to amplify by PCR a 700 bp fragment from pNMR11. The amplified fragment was treated with T4 polynucleotide kinase, digested with PstI, and ligated into pGEM32 (Promega) previously cut with PstI/SmeI. All constructs produced using PCR were sequenced. Orientation of constructs was determined by sequence analysis. Oligonucleotides used in this study were: NNMR187, 5′-ATC CAA TCA CCA GAA GTT GCA TCC CAT CCC GCG GCA AGC GGC TCT CTG CGA GCG-3′; NNMR262, 5′-CGC GCG AAG GGG TTC G-3′; NNMR258, 5′-GAC AGA TTA CTT GGT GCC GTG GTA-3′; NNJJ138, 5′-CTA GCG AAC CCC TTC G-3′; NNMR372, 5′-GAG AGG TGG CAT TGT CCG GGT TG-3′.
RNA Isolation.
Whole brain and heart tissue were isolated from 1 day old White Leghorn chicks and frozen in liquid nitrogen and kept at −70° until further use. Poly(A)+ RNA was isolated using the Fastrack 2.0 kit (Invitrogen).
RNase Protection and Primer Extension.
RNase protection experiments were performed using 32P-labeled antisense probes generated from pNMR25 linearized with HindIII as described (20). Protected fragments were electrophoresed through a 6% polyacrylamide gel containing 12 M urea using a radiolabeled RNA of known length and a sequencing ladder as size markers. The dried gel was exposed to Biomax MS film (Kodak) at −70°C for 2–3 days. Primer extension analysis was performed using primer NNMR372 as described. (25). Extension products were electrophoresed through a 6% polyacrylamide gel containing 12 M urea using a sequencing ladder as size markers. The dried gel was exposed as above for 5–7 days.
Ligand Binding Analysis.
To determine endogenous cellular mAChR levels, binding assays were performed with the muscarinic antagonist [3H]quinuclidinyl benzylate (41 Ci/mmol; Amersham; 1 Ci = 37 GBq) as described (26).
Reagents.
Recombinant rat ciliary neurotrophic factor (CNTF), recombinant human leukemia inhibitory factor (LIF), and interleukin 6 were purchased from Pepro-Tech. Recombinant murine cardiotrophin 1 (CT-1) and growth promoting activity were gracious gifts from Dianne Pennica (Genentech) and Ray Nishi (Oregon Health Sciences University, Portland), respectively. The chicken heart cDNA library was kindly donated by Douglas Fambrough (John Hopkins University, Baltimore). [γ-32P]ATP, [α-32P]UTP, and [α-35S]dATP were purchased from New England Nuclear.
RESULTS
Isolation of cm2 Promoter Region.
A genomic clone containing the entire cm2 coding region had been previously isolated in our laboratory (4). A potential intron 3′ splice acceptor site is located 41 bp upstream of the initiating codon in this clone. Because of the presence of introns upstream of the initiator codons of other mAChR genes (8) we wanted to determine if sequences corresponding to the 5′ cm2 transcript were located in the original genomic clone. Attempts at amplifying the 5′ end of the cm2 transcript by PCR techniques were unsuccessful, necessitating the screening of a chicken cDNA library. A chicken heart cDNA library was screened using a region coding for the third intracellular loop under high stringency. Several clones were isolated and found to represent transcripts of the cm2 gene. Two contained sequences that diverged from the genomic clone sequence at the predicted 3′ splice acceptor site (Fig. 1). An oligonucleotide, corresponding to 54 bp of sequence upstream of the 3′splice acceptor site in both cDNA clones (NNMR187, see Fig. 1), was used to screen a Southern blot of chicken genomic DNA and the previously isolated cm2 genomic clone. The oligonucleotide was found to hybridize to a single band of chicken genomic DNA but did not hybridize to the cm2 genomic clone containing the entire coding region of cm2 (data not shown). We screened a chicken genomic library with NNMR187 under high stringency to isolate genomic regions containing sequences found at the 5′ end of the cm2 cDNAs. Several clones were isolated and restriction mapped by Southern blot analysis (Fig. 1). Overlapping fragments that hybridized to NNMR187 were subcloned. A 789-bp PstI/HindIII fragment was sequenced on both strands (Fig. 1). The PstI/HindIII fragment was found to hybridize to the same size transcript as the third intracellular loop of cm2 by Northern blot analysis, demonstrating that it contained sequences corresponding to the 5′ end of the cm2 transcript (data not shown). An EcoRI fragment >10 kb in length, containing 2 kb of sequence upstream and >8 kb of sequence downstream of the 5′ splice donor site for intron one, did not hybridize to the original cm2 λ genomic clone containing the entire coding region (data not shown). Therefore, we estimate the size of intron one to be >8 kb. The genomic organization, 5′ untranslated regions of the two cDNAs, intron-exon boundaries for intron one, and nucleotide sequence of the 5′ PstI/HindIII fragment of the cm2 gene are shown in Fig. 1.
Determination of the Start Site of Transcription.
The start site of transcription of the cm2 gene was determined by both RNase protection and primer extension analyses (Fig. 2). Three major and three or more minor start sites of transcription were mapped in chicken brain while one major and several minor sites were mapped in chicken heart by RNase protection studies (Fig. 2A). The one major site start site found in chicken heart and several of the minor sites found in chicken heart are the same as those found in chicken brain. The protected fragments range in size from ≈70–196 bp. No protected fragments were seen in samples using yeast tRNA (data not shown). Similar results were seen in two additional experiments (data not shown). Primer extension analysis resulted in several extension products ranging from 68–202 nucleotides or more (Fig. 2B). Major extension products of ≈68 and 138 nucleotides were seen in both chicken heart and brain. An additional product of 134 nucleotides was seen in chicken heart. Several longer extension products were observed for chicken brain (major products) and chicken heart (minor products). No extension products were seen with yeast tRNA (data not shown). Similar results were seen in two additional experiments. The major extensions products correspond to the two regions of protected fragments found by RNase protection. From both RNase protection and primer extension analysis we conclude that there are two major regions, ≈337 and 393 bp upstream of intron one, where transcription begins in both chicken heart and brain (Fig. 1). Our results suggest that additional major start sites and a few minor sites, located farther upstream, are utilized in chicken brain and in chicken heart, respectively.
Figure 2.
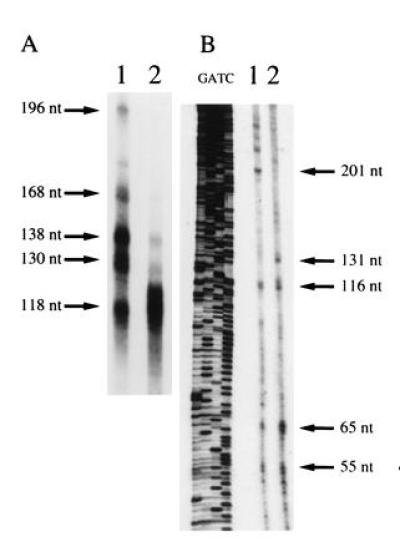
Determination of start site of transcription by RNase protection and primer extension analysis. (A) RNase protection experimented performed with 4 μg of poly(A)+ RNA isolated from 21 day chicken brain (lane 1) and chicken heart (lane 2). Protected fragments and approximate size, estimated from the mobility of a RNA of known size and a DNA sequencing ladder, are shown. Numbers correspond to nucleotides from position of downstream end of NNMR258 (see Fig. 1). No protected fragments were seen when 4 μg of yeast tRNA was used. Similar results were seen in two additional experiments. (B) Primer extension analysis. Oligonucleotide NNMR372 was hybridized to 5 μg of 21-day chicken brain (lane 1) and chicken heart (lane 2) poly(A)+ RNA at 50°C. Extension reactions were performed at 50°C with avian myeblastosis virus reverse transcriptase (Promega). A sequencing ladder generated using NNMR372 and a cm2 genomic clone is shown. Size of extension products are shown (nucleotide, nt). No extension products were seen when experiment was performed with 5 μg of yeast tRNA. Similar results were seen in two additional experiments.
Determination of Basal Levels of Expression for cm2 Reporter Gene Constructs.
To determine if sequences upstream of intron one contained promoter activity, constructs containing portions of the cm2 genomic region fused to the firefly luciferase gene were tested for expression of the luciferase gene in primary chicken heart cultures (Fig. 3). Constructs were transiently transfected into 9 day embryonic chicken heart cultures and assayed for luciferase activity. A constitutive simian virus 40–β-gal reporter plasmid was included in the transfection cocktail to correct for minor differences in transfection efficiency. A 789-bp fragment was capable of driving transcription of the firefly luciferase gene at levels 46 ± 4.6 (SEM, n = 13) times higher than vector alone. A construct containing an additional sequences 1.2 kb of upstream sequence (pNMR27) was as active, 43.6 ± 3.7, as the 789-bp construct (pNMR26); however, constructs containing an additional 2.5 kb (pNMR28) and 7.2 kb (pNMR29) were ≈40% less active. Both the 3.3- and the 8-kb constructs were equally active at promoting transcription (27.9 ± 2.3 and 28.3 ± 4.1, respectively).
Figure 3.
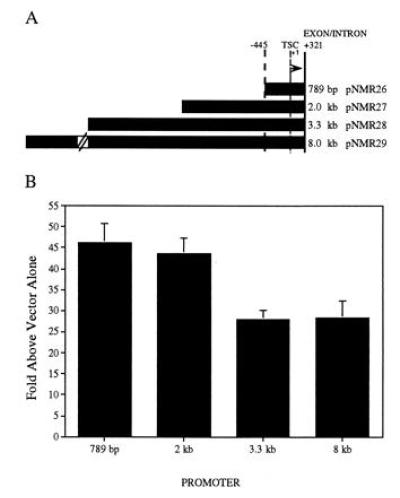
Diagram of reporter gene constructs used for transfection studies and basal level expression of constructs in chicken heart primary cultures. (A) Map of cm2 genomic regions fused to the firefly luciferase gene. Solid line labeled +321 indicates exon/intron boundary. Thin dashed line indicates the most 3′ start site of transcription. Thick dashed line labeled 445 indicates PstI site. Construct names and approximate size of cm2 genomic fragments are shown at right. (B) Basal level of transcription for reporter gene constructs transfected into chicken heart cultures. Levels of luciferase expression normalized to β-gal expression (see Materials and Methods) are shown as fold increase over reporter gene vector (pGL3-Basic) containing no promoter. Results are averages ± SEM from 13 separate experiments each performed with six replicate cultures.
We also determined if the cm2 reporter gene constructs were capable of driving transcription in neural cells. SN56 cells, a mouse neuroblastoma/septal cell line, were used as a model system for expression in the central nervous system. Transient transfections of the reporter constructs into SN56 cells resulted in luciferase expression between 5.6- and 13.6-fold higher than vector alone (Fig. 4). The 789-bp reporter construct was found to drive transcription 5.6 ± 1.2-fold higher than vector alone. Interestingly, the 2-kb reporter construct (pNMR27) demonstrated luciferase activity 13.6 ± 3.1-fold higher than vector alone. Constructs containing 3.3 kb and 8 kb fragments were capable of driving transcription at levels 5.9 ± 1.1 and 8.0 ± 1.5 higher than vector alone. All of the constructs were also capable of driving transcription in the human neuroblastoma cell line IMR-32 (data not shown).
Figure 4.
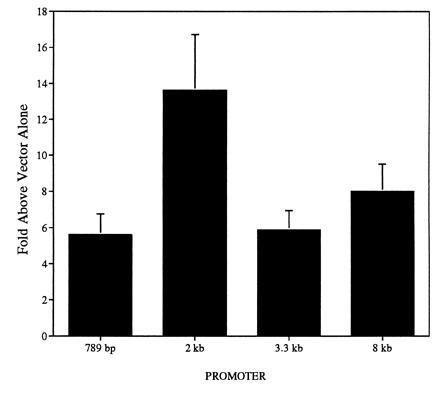
Basal level of expression of cm2 reporter gene constructs in SN56 cells. Basal level of expression of cm2 reporter gene constructs transfected into the mouse septal/neuroblastoma cell line SN56. Levels of luciferase expression normalized to β-gal expression (see Materials and Methods) are shown as fold increase over reporter gene vector (pGL3-Basic) containing no promoter. Results are averages ± SEM from four to five separate experiments each with three to six replicate cultures.
CNTF and LIF Induce Endogenous mAChR Expression in the Mouse Neuroblastoma/Septal Line SN56.
Since expression of m2 has been colocalized to regions of the central nervous system expressing ChAT (11) and ChAT expression has been shown to be induced by CNTF and LIF (23, 27, 28), we determined if CNTF and LIF regulated mAChR expression in the SN56 cells. Ligand binding assays were performed to determine if CNTF and LIF regulated the expression of endogenous mAChRs. Treatment with CNTF or LIF (25 ng/ml final concentration) for 48 hr resulted in a 39% ± 3.0 (SEM, n = 3) and a 40% ± 8.5 increase, respectively, in total mAChR binding sites.
The cm2 Promoter Is Induced by CNTF and LIF in SN56 Cells.
To test whether CNTF and LIF could induce transcription of the cm2 promoter, SN56 cells were transiently transfected with the cm2 reporter gene constructs and treated with cytokines (Fig. 5). Treatment with CNTF or LIF (25 ng/ml final concentration) for 32 hr resulted in an increase in transcription between 3.8- to 6.3-fold over control for the 2-, 3.3-, and 8-kb constructs while, the 789-bp construct and vector alone were only increased by 1.6–1.8-fold and 1.1–1.8-fold, respectively. The minimum region of the cm2 promoter necessary for induction by CNTF or LIF would appear to reside between 789 bp and 2 kb downstream of intron one (Fig. 5).
Figure 5.
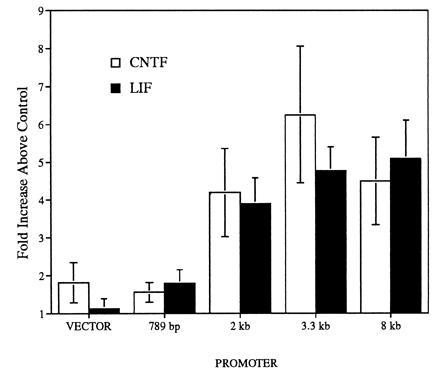
Induction of cm2 reporter gene constructs by CNTF and LIF in SN56 cells. Cells transfected with cm2 reporter gene constructs were treated for 32 hr, with CNTF or LIF at a final concentration of 25 ng/ml. Levels of luciferase expression normalized to β-gal expression (see Materials and Methods) are shown as fold increase over carrier treated cells. Results are averages from four to five separate experiments ± SEM each with three to six replicate cultures.
DISCUSSION
We have isolated the promoter region of the cm2 gene and demonstrated that it can drive transcription in both chicken heart cells and mammalian neural cell lines. The promoter region contains both a TATA box and CAAT box at positions −146 and +310, respectively, to the most downstream start site of transcription (Fig. 1) (29, 30). The position of these boxes relative to each other and to the start sites of transcription would make them unlikely candidates for elements involved in the initiation of transcription. Three AP2 sites (31) are found at positions −434, +123 and +195. An extremely cytosine-rich region is found between positions +117 and +171 containing 76% cytosine nucleotides. Within this region, two Sp1 sites, identical to sites found in the NPY gene (32), are located at positions +126 and +159. A third Sp1 site, identical to a site found in gammaglobin gene (33), is also found at position +254. Although we have not tested whether TATA-box binding protein or the TFIID complex can bind to and initiate transcription of the cm2 promoter, our data would suggest that the cm2 promoter is a TATA-less promoter or, a very unusual TATA-containing promoter (34, 35).
The cm2 promoter has multiple initiation sites for transcription, consistent with it being a TATA-less promoter. Some of these sites are used in both chicken heart and brain, but are preferentially used either in brain (sites farther upstream) or in heart (sites farther downstream) (Fig. 2). Our data suggests that additional start sites of transcription located farther upstream are utilized in chicken brain. We have been unable to detect any cm2 transcripts containing sequences between −445 bp and −1.7 kb, by Northern blot analysis in heart or brain (data not shown), suggesting that the majority of transcription initiation occurs within the 789-bp proximal to intron one. The porcine m2 gene has two exons containing noncoding sequences (8). While we cannot rule out that another upstream exon is in the cm2 gene, it would seem unlikely that another exon is located further upstream since the 789 bp PstI/HindIII fragment is capable of driving transcription in both chicken heart primary cultures and SN56 cells.
Different patterns of basal levels of expression are seen for the reporter constructs tested in chicken heart primary cultures and SN56 cells (Figs. 3 and 4). In chicken heart primary cultures the 789-bp and 2-kb constructs gave the highest level of expression with lower levels produced by the 3.3- and 8-kb constructs (Fig. 3). The 789-bp constructs appears to contain all the information sufficient for high level transcription while the 3.3- and 8-kb constructs may contain a silencer element, presumably located between the 2 and 3.3 kb of sequences upstream of intron one. In the SN56 cells, the 2-kb construct gave the highest level of expression while the other constructs showed roughly equal expression levels (Fig. 3). These results suggest the presence of a neural-specific activator located between 789 bp and 2 kb and, similar to the results from studies performed in heart cultures, the presence of a silencer element located between 2 and 3.3 kb upstream of intron one.
Our results demonstrate that both CNTF and LIF increase endogenous muscarinic receptor number and induce transcription of cm2 reporter constructs in SN56 cells, a murine septal/neuroblastoma hybrid cell line (Fig. 4). CNTF and LIF have been reported previously to block the increase in m2 mRNA levels that occurs after dissociation and culturing of neonatal rat superior cervical ganglion cells (36). This discrepancy probably results from phenotypic differences in the cell types used. The increase in total muscarinic binding (40%) sites is less than the increase in cm2 transcription (4- to 6-fold) consistent with the expression of multiple subtypes in the septum (11), some of which are not induced by CNTF and LIF. Interestingly, CNTF and LIF do not increase the expression of putative mouse m1 promoter constructs in SN56 cells (data not shown), consistent with subtype-specific effects of LIF and CNTF on muscarinic receptor expression. LIF and CNTF have been shown to increase ChAT expression in SN56 cells (29). Coordinate induction of both m2 receptors and ChAT would facilitate the feedback regulation of acetylcholine release since the m2 receptor is thought to act as an autoreceptor resulting in decreased acetylcholine release in some cell types of the brain.
CT-1, a newly isolated member of cytokine family, has been shown to induce cardiac hypertrophy in vitro. CT-1 can bind to and activate the LIF receptor complex resulting in some of the same biological activities as LIF (37), and requires the gp130/LIFRβ heterodimer for induction of hypertrophy in neonatal rat cardiomyocytes (38). In chicken heart primary cultures neither CNTF, LIF, growth promoting activity (a chicken homolog of CNTF, see ref. 39) or murine CT-1 regulated cm2 reporter gene constructs (data not shown). Cotransfection of a human interleukin 6 receptor, which also couples to and activates gp130, did not cause interleukin 6-mediated cm2 induction in heart cultures but is capable of producing interleukin 6-dependent induction of cm2 reporter gene constructs in SN56 cells (data not shown). The lack of response in heart cells may be a result of species specificity differences in the cytokines used or absence of the receptors and/or downstream signaling molecules at the developmental stage at which our assays were performed.
To our knowledge, this is the first report of the isolation of a m2 promoter region and the demonstration of induction of muscarinic receptors by cytokines. It needs to be determined what mechanisms may mediate this induction. The isolation and characterization of the rat m4 promoter has been recently reported by two groups (40, 41). The cm2 promoter is a GC rich TATA-less promoter and may contain a silencer element like the rat m4 promoter. The cm2 promoter should provide a useful tool for investigating the regulation of mAChRs in both cardiac and neural cells.
Acknowledgments
We thank Dr. Douglas Fambrough for his donation of the chicken heart cDNA library, and Drs. Dianne Pennica and Ray Nishi for their generous gifts of CT-1 and growth promoting activity, respectively. We thank Dr. Jan Krzysztof Blusztajn for providing SN56 cells. We thank members of our laboratory for many useful discussions. This work was supported by grants from the National Institutes of Health and an American Heart Association of Washington Postdoctoral Fellowship to M.L.R.
Footnotes
Abbreviations: mAChR, muscarinic acetylcholine receptor; ChAT, choline acetyltransferase; LIF, leukemia inhibitory factor; CNTF, ciliary neurotrophic factor; CT-1, cardiotrophin 1; β-gal, β-galactosidase.
Data deposition: The sequence reported in this paper has been deposited in the GenBank data base (accession no. U61850U61850).
References
- 1.Nathanson N M. Annu Rev Neurosci. 1987;10:195–236. doi: 10.1146/annurev.ne.10.030187.001211. [DOI] [PubMed] [Google Scholar]
- 2.Bonner T I. Trends Neurosci. 1989;12:148–151. doi: 10.1016/0166-2236(89)90054-4. [DOI] [PubMed] [Google Scholar]
- 3.Hulme E C, Birdsall N J M, Buckley N J. Annu Rev Pharmacol Toxicol. 1990;30:633–673. doi: 10.1146/annurev.pa.30.040190.003221. [DOI] [PubMed] [Google Scholar]
- 4.Tietje K M, Nathanson N M. J Biol Chem. 1991;266:17382–17387. [PubMed] [Google Scholar]
- 5.Gadbut A P, Galper J B. J Biol Chem. 1994;269:25823–25829. [PubMed] [Google Scholar]
- 6.Tietje K M, Goldman P S, Nathanson N M. J Biol Chem. 1990;265:2828–2834. [PubMed] [Google Scholar]
- 7.Habecker B A, Tietje K M, van Koppen C J, Creason S A, Goldman P S, Migeon J C, Parenteau L A, Nathanson N M. Life Sci. 1993;52:429–432. doi: 10.1016/0024-3205(93)90298-h. [DOI] [PubMed] [Google Scholar]
- 8.Peralta E G, Winslow J W, Peterson G L, Smith D H, Ashenazi A, Ramachandran J, Schimerlik M I, Capon D J. Science. 1987;236:600–605. doi: 10.1126/science.3107123. [DOI] [PubMed] [Google Scholar]
- 9.McKinnon L A, Nathanson N M. J Biol Chem. 1995;270:20636–20642. doi: 10.1074/jbc.270.35.20636. [DOI] [PubMed] [Google Scholar]
- 10.Katz A M. Physiology of the Heart. New York: Raven; 1992. [Google Scholar]
- 11.Caufield M P. Pharmacol Ther. 1993;58:319–379. doi: 10.1016/0163-7258(93)90027-b. [DOI] [PubMed] [Google Scholar]
- 12.Yan Z, Surmeier D J. J Neurosci. 1996;16:2592–2604. doi: 10.1523/JNEUROSCI.16-08-02592.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Dunlap K, Luebke J I, Turner T J. Trends Neurosci. 1995;18:89–98. [PubMed] [Google Scholar]
- 14.Herlitze S, Garcia D E, Mackie K, Hille B, Scheuer T, Catterall W A. Nature (London) 1996;380:258–262. doi: 10.1038/380258a0. [DOI] [PubMed] [Google Scholar]
- 15.Pan Z Z, Williams J T. J Neurosci. 1994;14:1332–1338. doi: 10.1523/JNEUROSCI.14-03-01332.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Spauschus A, Lentes K U, Wischmeyer E, Dissmann E, Karschin C, Karschin A. J Neurosci. 1996;16:930–938. doi: 10.1523/JNEUROSCI.16-03-00930.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kobayashi T, Ikeda K, Ichikawa T, Abe S, Togashi S, Kumanishi T. Biochem Biophys Res Commun. 1995;208:1166–1173. doi: 10.1006/bbrc.1995.1456. [DOI] [PubMed] [Google Scholar]
- 18.Lesage F, Duprat F, Fink M, Guillemare E, Coppola T, Lzdunski M, Hugnot J P. FEBS Lett. 1994;353:37–42. doi: 10.1016/0014-5793(94)01007-2. [DOI] [PubMed] [Google Scholar]
- 19.Duprat F, Lesage F, Guillemare E, Fink M, Hugnor JP, Bigay J, Lazdunski M, Romey G, Barhanin J. Biochem Biophys Res Commun. 1995;212:657–663. doi: 10.1006/bbrc.1995.2019. [DOI] [PubMed] [Google Scholar]
- 20.Sambrook J, Fritsch E F, Maniatis T. Molecular Cloning: A Laboratory Manual. 2nd Ed. Plainview, NY: Cold Spring Harbor Lab. Press; 1989. [Google Scholar]
- 21.Subers E M, Nathanson N M. J Mol Cardiol. 1988;269:131–140. doi: 10.1016/s0022-2828(88)80026-9. [DOI] [PubMed] [Google Scholar]
- 22.Lee H J, Hammond D N, Large T H, Wainer B H. Dev Brain Res. 1990;52:219–228. doi: 10.1016/0165-3806(90)90238-t. [DOI] [PubMed] [Google Scholar]
- 23.Johnson J A, Nathanson N M. J Biol Chem. 1994;269:18856–18863. [PubMed] [Google Scholar]
- 24.Edlund T, Walker M D, Barr P J, Rutter W J. Science. 1985;230:912–916. doi: 10.1126/science.3904002. [DOI] [PubMed] [Google Scholar]
- 25.Boorstein W R, Craig E A. Methods Enzymol. 1989;180:347–369. doi: 10.1016/0076-6879(89)80111-9. [DOI] [PubMed] [Google Scholar]
- 26.Halvorsen S W, Nathanson N M. J Biol Chem. 1981;256:7941–7948. [PubMed] [Google Scholar]
- 27.Symes A J, Rao M S, Lewis S E, Landis S C, Hyman S E, Fink J S. Proc Natl Acad Sci USA. 1993;90:572–576. doi: 10.1073/pnas.90.2.572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Berse B, Blusztajn J K. J Biol Chem. 1995;270:22101–22104. doi: 10.1074/jbc.270.38.22101. [DOI] [PubMed] [Google Scholar]
- 29.Breathnach R, Chambon P. Annu Rev Biochem. 1981;50:349–383. doi: 10.1146/annurev.bi.50.070181.002025. [DOI] [PubMed] [Google Scholar]
- 30.Gustafson T A, Kedes L. Mol Cell Biol. 1989;9:3269–3283. doi: 10.1128/mcb.9.8.3269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Mitchell P J, Wang C, Tjian R. Cell. 1987;50:847–861. doi: 10.1016/0092-8674(87)90512-5. [DOI] [PubMed] [Google Scholar]
- 32.Minth C D, Dixon J E. J Biol Chem. 1990;265:12933–12939. [PubMed] [Google Scholar]
- 33.Gumucio D L, Heilsteadt-Williamson H, Gray T A, Tarle S A, Shelton D A, Tagle D A, Slightom J L, Goodman M, Collins F. Mol Cell Biol. 1992;12:4919–4929. doi: 10.1128/mcb.12.11.4919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Pugh B F, Tjian R. Cell. 1990;61:1187–1197. doi: 10.1016/0092-8674(90)90683-6. [DOI] [PubMed] [Google Scholar]
- 35.Pugh B F, Tjian R. Genes Dev. 1991;5:1935–1945. doi: 10.1101/gad.5.11.1935. [DOI] [PubMed] [Google Scholar]
- 36.Ludlam W H, Zang Z, McCarson K E, Krause J E, Spray D C, Kessler J A. Dev Biol. 1994;164:528–539. doi: 10.1006/dbio.1994.1221. [DOI] [PubMed] [Google Scholar]
- 37.Pennica D, Shaw K J, Swanson T A, Moore M W, Shelton D L, Zioncheck K A, Rosenthal A, Taga T, Paoni N F, Wood W I. J Biol Chem. 1995;270:10915–10922. doi: 10.1074/jbc.270.18.10915. [DOI] [PubMed] [Google Scholar]
- 38.Wollert K A, Taga T, Saito M, Narazaki M, Kishimoto T, Glembotski C C, Vernallis A B, Heath J K, Pennica D, Wood W I, Chien K R. J Biol Chem. 1996;271:9535–9545. doi: 10.1074/jbc.271.16.9535. [DOI] [PubMed] [Google Scholar]
- 39.Leung D W, Parent A S, Cachianes G, Esch F, Coulombe J N, Nikolics K, Eckenstein F P, Nishi R. Neuron. 1992;8:1045–1053. doi: 10.1016/0896-6273(92)90126-x. [DOI] [PubMed] [Google Scholar]
- 40.Meida M, Haga T, Saffen D W. J Biol Chem. 1996;271:5177–5182. doi: 10.1074/jbc.271.9.5177. [DOI] [PubMed] [Google Scholar]
- 41.Wood I C, Roopra A, Harrington C, Buckley N J. J Biol Chem. 1995;270:30933–30940. doi: 10.1074/jbc.270.52.30933. [DOI] [PubMed] [Google Scholar]