

Abstract
The three largest known populations of amacrine cells in the rabbit retina were stained with fluorescent probes in whole mounts and counted at a series of retinal eccentricities. The retinas were counterstained using a fluorescent DNA-binding molecule and the total number of nuclei in the inner nuclear layer were counted in confocal sections. From the total number of inner nuclear layer cells and the known fraction of them occupied by amacrine cells, the fraction of amacrine cells made up by the stained populations could be calculated. Starburst cells made up 3%, indoleamine-accumulating cells made up 4%, and AII cells made up 11% of all amacrine cells. By referring four smaller populations of amacrine cells to the number of indoleamine-accumulating cells, they were estimated to make up 4% of all amacrine cells. Thus, 78% of all amacrine cells in the rabbit’s retina are known only from isolated examples, if at all. This proportion is similar in the retinas of the mouse, cat, and monkey. It is likely that a substantial fraction of the local circuit neurons present in other regions of the central nervous system are also invisible as populations to current techniques.
Keywords: starburst, indoleamine, AII cells, anatomy, cortex
It is generally acknowledged that the main classes of neuron in mammalian retinas include a large number of individual cell types. Amacrine cells, with which we will be concerned, may come in 20–30 varieties (for reviews, see refs. 1–3). They are often distinguishable by various biochemical markers, as well as by their pattern of branching and level of stratification in the inner plexiform layer. The last requires that the subpopulations of amacrine cells have different connections within the retina’s circuits, which in turn means that the multitude of cell types play different roles in creating and shaping the retina’s final physiological output. Yet most of these neurons are known only from occasional Golgi impregnations, if at all.
Even the number of amacrine cells was, until recently, unknown. Although amacrine cell numbers have sometimes been offered (see Discussion), most are based on weak evidence. The primary reason is that cytology, as viewed in nuclear stains, does not allow amacrine cells to be unambiguously identified. Another is that variability between tissue samples—notably the large differences in shrinkage during different kinds of histochemical processing—renders precise comparison of absolute cell densities impossible.
In a previous study (4), we proposed an approach to this problem. Samples of rabbit retina were fixed, osmicated, stained with uranyl acetate, serially sectioned at 1 μm, and stained again using toluidine blue and basic fuchsin. In photomicrographs taken at high magnification with differential-interference-contrast optics, processes of every cell could be visualized near their exit from the soma. This allowed every neuron in the inner nuclear layer to be identified as a horizontal, bipolar, or amacrine cell. Although the absolute densities of cells varied (due to different-sized eyes and deformation during processing) the relative fractions of horizontal, bipolar, and amacrine cells were found to be quite constant.
The constancy of proportions among the major cell classes is important, because it offers a base of reference by which data obtained using different histochemical methods may be interconverted. If the total number of inner nuclear layer neurons and the number of a stained subpopulation for the same retinal location are known, one may directly calculate the fraction of all horizontal cells, bipolar cells, or amacrine cells occupied by the stained neurons. Herein, we have made those measurements for the three largest amacrine cell populations that have been well characterized. We then used the results to estimate the fraction of all amacrine cells occupied by four smaller amacrine cell populations that had been described elsewhere. These specific populations are subgroups within two generic amacrine cell groups, those containing γ-aminobutyric acid (GABA) or glycine (5, 6); for comparison, GABA- and glycine-containing cells were also counted.
METHODS
All of the methods have been described in detail elsewhere and will only be summarized (4, 7–9). The retinas were from young adult (1.8–2.3 kg) New Zealand White rabbits. Removal of the eyes was done under deep anesthesia with a combination of xylazine and ketamine, after which the animal was euthanized by an overdose of the same anesthetics. The retinas were separated from the retinal pigment epithelium. Whole mounts for histochemistry were made after fixation of the retina with 4% paraformaldehyde for 1 h.
The nuclei of the displaced starburst cells were stained by intraocular injection of 1 μg of 4′,6-diamidino-2-phenylindole (DAPI) 1 day prior to sacrifice (7). Staining of indoleamine-amacrine cells was by immunohistochemistry (8, 9). Retinas were incubated in vitro in Ames medium (10) in the presence of 2 μM 5-hydroxytryptamine for 1 h. After fixation, serotonin was visualized by a primary antibody against it (Incstar, Stillwater, MN), biotinylated secondary antibodies, and avidin-fluorescein isothiocyanate for visualization. AII amacrines were stained by using primary antibodies against parvalbumin (Sigma) or calretinin (Chemicon). Each retina was counterstained, after immunohistochemical staining, with 4 μM ethidium homodimer (Molecular Probes) for 2 h. The retinas were coverslipped under Vectashield (Vector Laboratories).
Counting was done from photomicrographs made at intervals along a dorsal to ventral transect passing through the optic nerve head. The final magnifications were as follows: starbursts, ×225 (color prints); indoleamine-accumulating cells, ×415; AII cells, ×650. At the same retinal locations serial optical sections were made through the inner layer using a Bio-Rad MRC600 confocal microscope. The sections were taken at Z intervals of 1 μm and had a nominal optical thickness of 1 μm. The serial optical sections were printed at a final magnification of ×2400. By following each nucleus throughout its extent (i.e., through four to six sections), every cell in the selected region was counted. For further details on counting and reconstruction, see ref. 4.
For immunohistochemistry of GABA and glycine, the anterior segment and vitreous body were removed and the eye cups were immersed in fresh oxygenated Ames medium at room temperature. The retinas were isolated from the pigmented epithelium and a strip of retina centered along the vertical meridian was flattened on filter paper. Subsequently, they were immersed in fixative consisting of 2.5% glutaraldehyde, 1% paraformaldehyde, and 0.1 M sodium phosphate (pH 7.2). Fixation was carried on at room temperature for the initial 15 min and then at 4°C for 18–20 h. The retinas were separated from the filter paper, washed extensively in 0.1 M phosphate buffer, and then cut into a ladder of horizontal strips parallel to the vertical meridian, each piece about 4 mm long and 1 mm wide. The strips of retinal tissue were rinsed in phosphate buffer, dehydrated in a graded series of ethanols, and embedded flat in Medcast (Ted Pella, Redding, CA). Serial semi-thin sections (1 μm thick) were cut from retinal blocks at selected locations with a Leica Supernova ultramicrotome, using a diamond knife. Two locations were chosen, one 3 mm ventral to the optic nerve head (near the peak of the visual streak) and one 3 mm more ventral. For each block, three series of 100 sections were cut: two were used for GABA and glycine immunohistochemistry and the third was used for counting the total number of inner nuclear layer cells.
Serial sections were collected on glass slides and dried on a hot plate (60°C) for 1–2 h. Sections were etched with a mixture of 1:3 saturated ethanolic NaOH/absolute ethanol for 12 min, followed by three 2-min rinses in absolute ethanol and then rinsed in distilled water for 15 min. The sections were rinsed again with 0.02 M Tris/phosphate buffer (TPBS, pH 7.2), for 10 min and then incubated in a solution of 10% normal goat serum and TPBS for 2–3 h to block unspecific staining. Subsequently, the sections were incubated in a solution of antisera, 5% normal goat serum, and TPBS for 12–18 h at 4°C, in humid chambers. Antisera were from Chemicon, anti-GABA (raised in rabbit) was used at a dilution of 1:2000, and anti-glycine (raised in rabbit) was used at a dilution of 1:250. The retinal sections were rinsed several times in TPBS and incubated for 3 h in 5-nm-gold-conjugated goat anti-rabbit IgG (Amersham) diluted 1:25 in TPBS with 1% normal goat serum. The sections were rinsed in TPBS for 1 h, rinsed again in water, and air-dried. Subsequently, the sections were incubated with silver intensification solution (Amersham), for 20 min, until the immunopositive cells were clearly visible under the light microscope. The sections were rinsed in water, dried, coverslipped, and photographed with a Zeiss Axioplan microscope using a ×40 or ×63 objective. The series of sections to be used for counting the total number of inner nuclear layer cells was counterstained with toluidine blue on a hot plate, coverslipped, and photographed at ×100.
Immunostaining of the cells appeared highly specific. Despite the presence of a certain level of background, labeled cells were readily distinguished from unlabeled cells even in single micrographs and were consistently labeled through series of consecutive sections. They were easy to count. Processes of horizontal cells were not labeled using the anti-GABA antiserum. Staining of the processes is variably reported in mammalian retinas (see ref. 3). Perhaps their pool of GABA is poorly retained under our conditions of fixation (11, 12).
RESULTS
Three specifically stained populations of amacrine cells were counted: starburst cells, indoleamine-accumulating cells, and AII amacrine cells. For each, the cells were counted in whole mounts at intervals of 1.5 mm along a vertical transect passing through the optic nerve head. At every second counting location (i.e., every 3 mm), the ethidium-counterstained nuclei of all inner nuclear layer cells were also counted.
Starburst cells were stained by their accumulation of DAPI. Fig. 1A shows the nuclei of the starburst cells in the ganglion cell layer. A regular population of bright uniformly stained neurons is visible, as described extensively in previous studies (7, 13, 14). For three retinas, the density of starburst cells ranged from 660 cells per mm2 in the visual streak to 175 cells per mm2 in the dorsal periphery. These values were comparable to those obtained previously. Starburst cells in the inner nuclear layer, which were our primary interest, are slightly more numerous than displaced cells in the visual streak but have the same number elsewhere (2, 7, 15, 16). The absolute difference is small and no correction was made; the number of starburst cells in the ganglion cell layer is taken as a measure of the conventionally placed ones. At six points across the retina, the total number of inner nuclear layer neurons was also counted (Fig. 2). From previous work (table 1 of ref. 4), the fractions of these neurons that are amacrine cells is known. At the specified distances from the optic nerve head, they are: 6 mm dorsal, 27.2%; 3 mm dorsal, 28.6%; 3 mm ventral, 34.8%; 6 mm ventral, 32.0%; 9 mm ventral, 31.5%; 12 mm ventral, 29.0%. This allowed an estimate of the percentage of all amacrines in the inner nuclear layer that were starburst cells. It changed little across the retina. On average, starburst cells made up 3% of the amacrine cells in the inner nuclear layer (Fig. 3).
Figure 1.

(Left) Staining of specific amacrine cell populations. (A) Displaced starburst amacrine cells, stained by accumulation of DAPI. (B) Serotonin-accumulating amacrine cells in the inner nuclear layer, identified by immunohistochemistry. (C) AII amacrine cells, identified by immunohistochemistry. (Bar = 50 μm.) (Right) Serial horizontal sections (by confocal microscopy) through the inner nuclear layer. The nuclei were stained with ethidium homodimer. Cells were counted by following each cell from its beginning to end in the serial sections. (Bar = 10 μm.)
Figure 2.
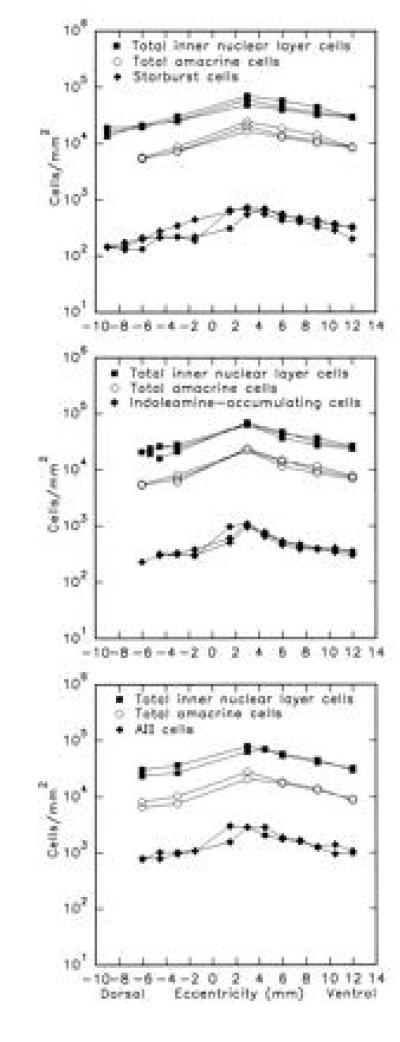
Population densities of starburst, indoleamine-accumulating, and AII amacrine cells. For each, the population of stained amacrine cells was counted in low power whole mount views (Fig. 1 A–C). In the same tissues, the total density of inner nuclear layer cells was counted at a series of seven retinal eccentricities (Fig. 1 Right). The total density of amacrine cells (open circles) was established by multiplying the total nuclei present at six retinal eccentricities by the fraction of amacrine cells known to be present at those retinal eccentricities (4).
Figure 3.
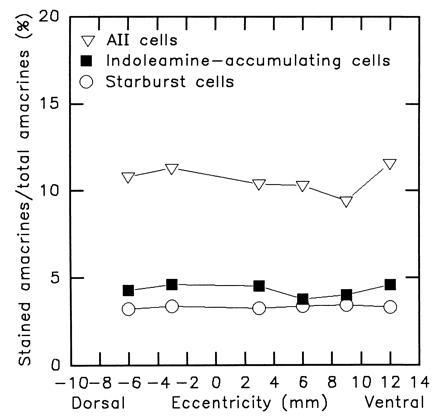
Fraction of all amacrine cells occupied by starburst, indoleamine-accumulating, and AII amacrine cells. These percentages were calculated from the data of Fig. 2.
Indoleamine-accumulating amacrine cells were stained by immunohistochemistry after uptake of serotonin. The mosaic of indoleamine-accumulating amacrine cells is shown in Fig. 1B. The density of indoleamine-accumulating cells ranged from 1043 cells per mm2 in the central retina to 231 cells per mm2 in the dorsal periphery. These values are in accord with those obtained in previous studies (2, 8). The retinas were counterstained and the total number of cells in the inner nuclear layer was established for six retinal locations. On average, indoleamine-accumulating amacrine cells made up 4% of the total amacrine cell population.
AII amacrine cells were counted by immunohistochemistry using antibodies directed against parvalbumin. The mosaic of AII amacrine cells is shown in Fig. 1C. A regular population of cell bodies is visible, with the lobular appendages of their dendritic arbors slightly out of focus. Other retinas were stained with antibodies against calretinin, which gives a brighter staining. However, a second population of amacrine cells is also stained with calretinin and this made counting more difficult, especially in the central retina. With parvalbumin, these second cells were less brightly stained and could more readily be distinguished from the AII cells. Data shown are from parvalbumin-stained retinas. Perhaps because of this issue, the data obtained in counting AII amacrines were more variable than for the other populations. However, the results obtained were in good agreement with those obtained by other workers (17, 18). The cells had a peak density of 2887 cells per mm2 near the visual streak and a density of 795 cells per mm2 in the dorsal periphery. Comparison with the total number of inner nuclear layer neurons in these retinas indicated that AII amacrine cells made up 11% of the total amacrine cell population.
GABA- and glycine-containing cells were studied using sections from glutaraldehyde-fixed embedded tissue (Fig. 4). Immunohistochemical identification of the cell populations was done by post-embedding immunohistochemistry (11, 22) and the total number of cells in the inner nuclear layer was learned from adjacent series stained with toluidine blue. In general, staining for GABA and glycine conformed to that reported previously in the rabbit and other mammals (3, 6). GABA was sharply localized in cells at the inner margin of the inner nuclear layer. No GABA-containing horizontal cells were observed, but only a handful of horizontal cell bodies would be expected in these series and the GABA staining of horizontal cell processes in mammals is usually weak (3, 6, 23, 24). A few glycine-containing cells were located at the outer margin of the inner nuclear layer. These presumably correspond to the glycine-containing bipolar cells reported in other mammalian species (25–27) but were negligible in number and are not included in the quantitative analysis. The levels of glycine were more variable than GABA.
Figure 4.
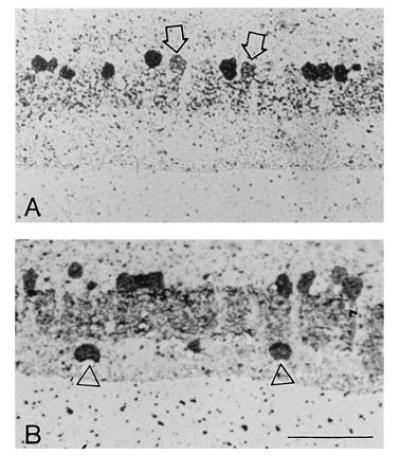
Staining of etched semi-thin sections for endogenous GABA and glycine. Location 6 mm ventral to the optic nerve head. (A) Glycine-accumulating cells. (B) GABA-accumulating cells. In both, the ganglion cell layer is at the bottom. Arrows in A point to weakly labeled cells at the top of the inner nuclear layer, presumably glycine-accumulating bipolar cells. Triangles in B point to GABA-accumulating displaced amacrine cells, almost certainly starburst cells. They are shown as a positive control, since they are known to contain GABA (19–21). (They were not counted, because the analysis is concerned with the inner nuclear layer.)
The overall densities of the cells (both neurotransmitter-labeled and the total cell population) appeared higher than for the cells studied in whole mounts, because of shrinkage of the tissue during dehydration. (A small linear shrinkage becomes a large areal shrinkage.) In the central retina (3 mm ventral to the optic nerve), the total density of cells was 169,600 cells per mm2. Of these, 34.8% or 59,020 cells per mm2 should be amacrine cells (4). The density of the GABA immunoreactive cells was 22,422 cells per mm2 and the density of the glycine immunoreactive cells was 26,250 cells per mm2. Of the total amacrine cells, GABA-immunoreactive cells were thus 37.9% and glycine cells were 44.5%. Six millimeters ventral to the optic nerve head, the total cells were 105,600 cells per mm2. Thirty-two percent of these or 33,792 cells per mm2 should be amacrine cells (4). GABA-immunoreactive cells made up 37.4% (12,611 cells per mm2) and glycine cells made up 51.9% (17,526 cells per mm2) of that total.
DISCUSSION
Our major finding was that starburst, indoleamine-accumulating, and AII amacrine cells make up 3%, 4%, and 11%, respectively, of all amacrine cells in the rabbit’s retina. The staining was clear (Fig. 1) and the spatial densities agree well with those observed in whole mounts by others [starbursts (2, 7, 15), indoleamine-accumulating (8, 28), and AII, (17, 18)]. The total number of amacrine cells was obtained by multiplying the total density of inner nuclear layer neurons by the known fraction of amacrine cells (4) and this sets a limit to the precision of the current estimates. A small error in the fraction of amacrine cells previously estimated would translate to a larger error in the fraction of all amacrine cells represented by the subpopulation studied here. However, the total number of cells was easy to learn from confocal series, and the fractions established previously were learned, among other means, by electron microscopy of serial sections. The imaginable errors would affect our fundamental conclusion little: starbursts, indoleamine-accumulating cells, and AII amacrines are a small fraction of all amacrine cells (Fig. 5).
Figure 5.
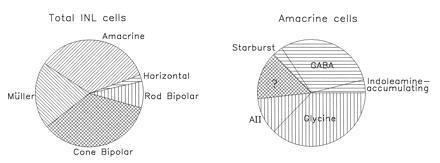
Major classes of cell present in the inner nuclear layer of the rabbit’s retina. Data for the generic amacrine cell classes (GABAergic and glycinergic) and for the three specifically stained amacrine cell populations are from the present study. The remaining information is from ref. 4.
Published work indicates that the numbers of cells in other stained populations of amacrine cell are even fewer than those studied herein. The original research in those studies lacked a base of reference, so that the fractions of all amacrine cells cannot be precisely stated, but as an approximation, data collected from immunohistochemistry of lightly fixed whole mounts mounted in aqueous solutions or glycerol can be compared without gross error. Four such populations of amacrine cell are the cells containing NADPH diaphorase (29–31), those containing catecholamines (31–33), those containing vasoactive intestinal peptide (30), and the DAPI-3 cells (2, 34). At a point in the midperiphery (9 mm ventral to the optic nerve head), the respective population densities of these cells were: 70, 19, 22, and 245 cells per mm2. At the same eccentricity the indoleamine-accumulating cells had a density of 398 cells per mm2, representing 4% of the total amacrine cell population at that eccentricity (Figs. 2 and 3). Using these proportions, and the present results, the four other populations make up the following fractions of the total population of amacrine cells as follows: NADPH-diaphorase, 0.7%; catecholamines, 0.2%; vasoactive intestinal peptide, 0.2%; DAPI-3, 2.5%. In total, the identified amacrine cells in our measurements and others’ make up 22% of the total amacrine cell population. Thus 78% of all amacrine cells are known only from Golgi-stained exemplars, if at all.
This fraction pertains only to the amacrine cells located in the inner nuclear layer. Including those displaced to the ganglion cell layer would change the value, but only by 2–3% (7, 35).
In a review of amacrine cell mosaics, Vaney (2) estimated that the rabbit’s retina contains 5,000,000 amacrine cells. The estimate derived from counting DAPI-labeled cells at the “inner margin” of the inner nuclear layer for one field and extrapolating along unspecified density contours to the whole retina. This method is too subjective to be generally useful. [For example, in the cat the total number of amacrine cells estimated in the same way ranged from 5,200,000 to 8,200,000 for different retinas (2, 36, 37).] Even so, Vaney estimated that identified populations account for only one quarter of amacrines in the rabbit retina, a conclusion nearly identical to ours.
Having many unidentified amacrine cells is not a peculiarity of the rabbit’s retina. In the mouse, amacrine cells have been estimated to make up 39% of the cells in the inner nuclear layer (38). In the cat, the exact fraction has apparently not been reported, but from published absolute densities (2, 36), it should be roughly 20%. Both of these estimates derive from counts based on nuclear cytology. Better data are available for the monkey (39), where the fraction of amacrine cells for a point 5 mm from the fovea was estimated by a combination of cytology, electron microscopy, and immunohistochemistry to be 28%. The absolute spatial densities of the known populations of amacrine cells in these retinas are as low as those described herein in rabbit; no populations much more dense than those found in rabbits have yet been observed (2, 3). We conclude that the large majority of amacrine cells in most mammalian retinas are unidentified.
To put this in perspective, horizontal cells make up 2% of all cells in the rabbit’s inner nuclear layer (4). The number of unidentified amacrine cells is thus approximately 14 times the total number of horizontal cells. Even if one assumes that only half of the unidentified amacrine cells have wide dendritic arbors (6), amacrine cells seem likely to be the more important laterally conducting pathway in the retina.
What is the identity of the missing amacrine cells? Our results show, confirming work in goldfish by Marc and coworkers (6, 12), that a large majority of all amacrine cells contain glycine or GABA. These populations include the starburst (GABA), indoleamine-accumulating (GABA), and AII (glycine) amacrine cells and must include many other morphological (and functional) types of amacrine cells as well. As was pointed out by Marc (6), the cell bodies of the glycinergic cells tend to be smaller than those of the GABAergic, and this implies that they have smaller dendritic arbors. Quite possibly many of the glycinergic cells are narrow-field amacrines. Other than occasional reports from Golgi catalogues, there are few further clues. Molecules concerned with intracellular signaling may provide markers for other populations, as parvalbumin has for AII.
Local circuit neurons analogous in some ways to amacrine cells are present in most regions of the central nervous system. For example, the neocortex contains a variety of small neurons that communicate locally. Many of them contain GABA (40), but this fact alone is no more restrictive in the cortex than in the retina: careful work using the Golgi method by Lund and colleagues (41–43) demonstrated 17 morphologically distinct varieties of local circuit neuron in layers 4C and 5 of the striate cortex. In this context, “morphologically distinct” does not mean only that the cells have a pattern of processes recognizable to an anatomist’s eye: the processes run to different locations within the cortex and this requires that they interact synaptically with different sets of other neurons. They are not only morphologically distinct, they carry out different functions within the cortex.
Because of the limitations of the Golgi method, the numbers of each cell type are unknown (43). It is quite possible that numerically important cells are mixed together in these descriptions with numerically sparse ones. Indeed, there is no guarantee that important populations of local circuit neurons have not escaped staining altogether. [The first starburst amacrine cell was apparently stained in 1979—almost a century after Golgi investigation of the retina began (44).] A way to avoid such risks would be to begin to develop quantitative information about the cell populations of the cortex.
Acknowledgments
We thank Mr. Alberto Bertini for photography. This research was supported by National Institutes of Health Grant EY01075 and an Italy-USA grant from the Consiglio Nazionale delle Ricerche of Italy.
Footnotes
Abbreviations: GABA, γ-aminobutyric acid; DAPI, 4′,6-diamidino-2-phenylindole.
References
- 1.Masland R H. Trends Neurosci. 1988;11:405–410. doi: 10.1016/0166-2236(88)90078-1. [DOI] [PubMed] [Google Scholar]
- 2.Vaney D I. Prog Retinal Res. 1991;9:1–28. [Google Scholar]
- 3.Wässle H, Boycott B B. Physiol Rev. 1991;71:447–480. doi: 10.1152/physrev.1991.71.2.447. [DOI] [PubMed] [Google Scholar]
- 4.Strettoi E, Masland R H. J Neurosci. 1995;15:875–888. doi: 10.1523/JNEUROSCI.15-01-00875.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Massey S C, Redburn D A. Prog Neurobiol. 1987;28:55–96. doi: 10.1016/0301-0082(87)90005-0. [DOI] [PubMed] [Google Scholar]
- 6.Marc R E. Prog Retinal Res. 1989;8:67–107. [Google Scholar]
- 7.Masland R H, Mills J W, Hayden S A. Proc R Soc London B. 1984;223:79–100. doi: 10.1098/rspb.1984.0084. [DOI] [PubMed] [Google Scholar]
- 8.Sandell J H, Masland R H. J Neurosci. 1986;6:3331–3347. doi: 10.1523/JNEUROSCI.06-11-03331.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Masland R H, Rizzo J F, III, Sandell J H. J Neurosci. 1993;13:5194–5202. doi: 10.1523/JNEUROSCI.13-12-05194.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ames A I, Nesbett F B. J Neurochem. 1981;37:867–877. doi: 10.1111/j.1471-4159.1981.tb04473.x. [DOI] [PubMed] [Google Scholar]
- 11.Marc R E, Liu W L, Kalloniatis M, Raiguel S F, van Haesendonck E. J Neurosci. 1990;10:4006–4034. doi: 10.1523/JNEUROSCI.10-12-04006.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Marc R E, Murry R F, Basinger S F. J Neurosci. 1995;15:5106–5129. doi: 10.1523/JNEUROSCI.15-07-05106.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Tauchi M, Masland R H. Proc R Soc London B. 1984;223:101–119. doi: 10.1098/rspb.1984.0085. [DOI] [PubMed] [Google Scholar]
- 14.Vaney D I. Proc R Soc London B. 1984;220:501–508. doi: 10.1098/rspb.1984.0016. [DOI] [PubMed] [Google Scholar]
- 15.Famiglietti E V, Tumosa N. Brain Res. 1987;413:398–403. doi: 10.1016/0006-8993(87)91037-7. [DOI] [PubMed] [Google Scholar]
- 16.Famiglietti E V. J Neurosci. 1985;5:562–577. doi: 10.1523/JNEUROSCI.05-02-00562.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Vaney D I, Gynther I C, Young H M. J Comp Neurol. 1991;310:154–169. doi: 10.1002/cne.903100203. [DOI] [PubMed] [Google Scholar]
- 18.Casini G, Rickman D W, Brecha N C. J Comp Neurol. 1995;356:132–142. doi: 10.1002/cne.903560109. [DOI] [PubMed] [Google Scholar]
- 19.Kosaka T, Tauchi M, Dahl J L. Exp Eye Res. 1988;70:605–617. doi: 10.1007/BF00247609. [DOI] [PubMed] [Google Scholar]
- 20.O’Malley D M, Sandell J H, Masland R H. J Neurosci. 1992;12:1394–1408. doi: 10.1523/JNEUROSCI.12-04-01394.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Brecha N, Johnson D, Peichl L, Wässle H. Proc Natl Acad Sci USA. 1988;85:6187–6191. doi: 10.1073/pnas.85.16.6187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Ehinger B, Ottersen O P, Storm-Mathisen J, Dowling J E. Proc Natl Acad Sci USA. 1988;85:8321–8325. doi: 10.1073/pnas.85.21.8321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Chun M-H, Wässle H, Brecha N. Neurosci Lett. 1988;94:259–263. doi: 10.1016/0304-3940(88)90027-4. [DOI] [PubMed] [Google Scholar]
- 24.Chun M H, Wässle H. J Comp Neurol. 1989;279:55–67. doi: 10.1002/cne.902790106. [DOI] [PubMed] [Google Scholar]
- 25.Hendrickson A E, Koontz M A, Pourcho R G, Sarthy P V, Goebel D J. J Comp Neurol. 1988;273:473–487. doi: 10.1002/cne.902730404. [DOI] [PubMed] [Google Scholar]
- 26.Pourcho R G, Goebel D H. J Neurosci. 1987;7:1189–1197. doi: 10.1523/JNEUROSCI.07-04-01189.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Pourcho R G, Goebel D J. J Neurosci. 1987;7:1178–1188. doi: 10.1523/JNEUROSCI.07-04-01178.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Negishi K, Teranishi T, Kato S. Neurosci Lett. 1984;45:27–32. doi: 10.1016/0304-3940(84)90324-0. [DOI] [PubMed] [Google Scholar]
- 29.Sandell J H. J Comp Neurol. 1985;238:466–472. doi: 10.1002/cne.902380410. [DOI] [PubMed] [Google Scholar]
- 30.Sagar S M. Brain Res. 1987;426:157–163. doi: 10.1016/0006-8993(87)90435-5. [DOI] [PubMed] [Google Scholar]
- 31.Vaney D I, Young H M. Brain Res. 1988;474:380–385. doi: 10.1016/0006-8993(88)90455-6. [DOI] [PubMed] [Google Scholar]
- 32.Tauchi M, Madigan N M, Masland R H. J Comp Neurol. 1990;293:178–189. doi: 10.1002/cne.902930203. [DOI] [PubMed] [Google Scholar]
- 33.Brecha N C, Oyster C W, Takahashi E S. Invest Ophthalmol Visual Sci. 1984;25:66–70. [PubMed] [Google Scholar]
- 34.Ehinger B, Zucker C L. Invest Ophthalmol Visual Sci. 1996;37:138. (abstr.). [Google Scholar]
- 35.Hughes A, Vaney D I. J Comp Neurol. 1980;189:169–189. doi: 10.1002/cne.901890110. [DOI] [PubMed] [Google Scholar]
- 36.Wässle H, Schafer Trenkler I, Voigt T. J Neurosci. 1986;6:594–604. doi: 10.1523/JNEUROSCI.06-02-00594.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Wässle H, Chun M H, Müller F. J Comp Neurol. 1987;265:391–408. doi: 10.1002/cne.902650308. [DOI] [PubMed] [Google Scholar]
- 38.Young R W. Anat Rec. 1985;212:199–205. doi: 10.1002/ar.1092120215. [DOI] [PubMed] [Google Scholar]
- 39.Martin P R, Grünert U. J Comp Neurol. 1992;323:269–287. doi: 10.1002/cne.903230210. [DOI] [PubMed] [Google Scholar]
- 40.Fitzpatrick D, Lund J S, Schmechel D E, Towles A C. J Comp Neurol. 1987;264:73–91. doi: 10.1002/cne.902640107. [DOI] [PubMed] [Google Scholar]
- 41.Lund J S. J Comp Neurol. 1987;257:60–92. doi: 10.1002/cne.902570106. [DOI] [PubMed] [Google Scholar]
- 42.Lund J S, Hawken M J, Parker A J. J Comp Neurol. 1988;276:1–29. doi: 10.1002/cne.902760102. [DOI] [PubMed] [Google Scholar]
- 43.Lund J S, Yoshioka T. J Comp Neurol. 1991;311:234–258. doi: 10.1002/cne.903110206. [DOI] [PubMed] [Google Scholar]
- 44.Perry V H. Proc R Soc London B. 1979;204:363–375. doi: 10.1098/rspb.1979.0033. [DOI] [PubMed] [Google Scholar]