

Abstract
Attempts to rescue retinal ganglion cells from retrograde degeneration have had limited success, and the residual function of surviving neurons is not known. Recently, it has been found that axotomized retinal ganglion cells die by apoptotic mechanisms. We have used adult transgenic mice overexpressing the Bcl-2 protein, a powerful inhibitor of apoptosis, as a model for preventing injury-induced cell death in vivo. Several months after axotomy, the majority of retinal ganglion cells survived and exhibited normal visual responses. In control wild-type mice, the vast majority of axotomized retinal ganglion cells degenerated, and the physiological responses were abolished. These results suggest that strategies aimed at increasing Bcl-2 expression, or mimicking its function, might effectively counteract trauma-induced cell death in the central nervous system. Neuronal survival is a necessary condition in the challenge for promoting regeneration and eventually restoring neuronal function.
Prevention of degeneration of axotomized retinal ganglion cells (RGCs) in adult mammals has been attempted several times using different approaches such as treatment with neurotrophic factors, microglia-suppressing factors, transplant of embryonic tissue, and graft of peripheral nerve sheaths (1–8). As yet, long-term survival has been achieved for only a limited number of axotomized RGCs. Recent results from our laboratory (9, 10), however, have shown that the vast majority of RGCs (approximately 71,000 cells: more than 70% of the total RGC number) survive optic nerve axotomy for at least 3.5 months in adult transgenic mice (line NSE73a) overexpressing the human oncoprotein Bcl-2 (11). This protein is a powerful inhibitor of apoptotic cell death (12–14). In control wild-type mice, only few RGCs survived 2 months after optic nerve transection (approximately 2200 cells: 5% of the total RGC number) (9, 10).
Long-term survival of a large fraction of RGCs after optic nerve injury is a necessary condition for any attempt to promote regeneration and eventually restore visual function. To this aim, however, essential condition is that RGCs respond normally to light (i.e., display the same response properties of RGCs of intact mice to a wide range of visual stimuli). We have addressed this point by evaluating the response characteristics of RGCs in transgenic Bcl-2 mice, as well as in wild-type mice, in control conditions and at different times after unilateral optic nerve section.
The visually driven electrical activity of RGCs has been recorded noninvasively by means of the electroretinogram in response to gratings of different spatial frequencies, temporally modulated in contrast [pattern electroretinogram (P-ERG)]. In all mammals studied so far, including human, the P-ERG has been shown to depend on RGC activity, and it is abolished in retrograde RGC degeneration (15–19). It is worth noticing that P-ERG losses may precede anatomical losses of RGCs (20, 21), thereby signaling early retinal dysfunction that eventually leads to cell death.
We report evidence that, in transgenic Bcl-2 mice, most RGCs survive 3.5 months after axotomy. In the same animals the visual response of RGCs is still recordable with normal characteristics. By contrast, in wild-type mice, RGCs are massively degenerated 2 months after axotomy and the visual response of RGCs is abolished.
MATERIALS AND METHODS
Transgenic Mice.
We have used line NSE73a of the transgenic mice expressing human Bcl-2 (11). In this line, human Bcl-2 protein is expressed under the control of the neuron-specific enolase promoter. In the retina, Bcl-2 expression has been detected in the majority of cells of the inner nuclear layer and in all the cells of the ganglion cell layer. Bcl-2 expression is not detectable in the photoreceptor layer. The colony, started from one albino founder, was fostered by crossing males heterozygous for the transgene with C57BL/6J females. Albino offsprings were used for reproduction, and the experiments were carried out on pigmented mice 6–8 months of age. The presence of the Bcl-2 transgene has been checked by PCR on tail tissue of all animals [data shown elsewhere (10)].
Optic Nerve Section.
Mice were anesthetized with Avertin (tribromoethanol in amylene hydrate, 20 μl/g of body weight administered intraperitoneally). After partial craniotomy, the optic nerve was exposed by suctioning the overlaying tissue and completely severed between the optic foramen and the chiasm (approximately 2 mm from the papilla), in such a way that the two stumps were clearly separated from each other. This approach avoids possible damages to the ophthalmic artery.
Electrophysiological Recordings.
Mice under Avertin anesthesia were leaned on a plastic head holder. The P-ERG was recorded by means of corneoscleral Ag–AgCl rings (3.5 mm in diameter and 0.2 mm thick) inserted below the eyelids. Recording was monocular, and the electrode placed on the contralateral black patched eye served as reference. For most experiments, visual stimuli were horizontal gratings with a sinusoidal luminance profile of different spatial frequency (0.1–0.7 c/degree) and fixed contrast (90%), temporally modulated in counterphase (sinusoidally) at 6 Hz (12 contrast reversals per s) at constant mean luminance (10 cd/m2). In other experiments, either the contrast was varied (20–90%) at fixed spatial (0.1 c/degree) and temporal frequency (6 Hz) or the temporal frequency was varied (1–24 Hz) at fixed spatial frequency (0.1 c/degree) and contrast (90%). Visual stimulus was also a uniform field (10 cd/m2 luminance) flickering sinusoidally at 6 Hz (90% modulation depth). Stimuli were generated by a specialized grating generator (Cambridge VSG2/2), displayed on a high-resolution monitor (Barco CCID 7751: 24 × 26 cm display), centered on the pupil and presented at 20-cm distance (subtending a field of 69 × 74 degrees) against a background luminance of 1 c/m2. Retinal signals were amplified 50,000-fold, filtered between 1 and 100 Hz, digitized at 1 kHz with 12-bit resolution, and averaged in synchrony with the stimulus period over 500 stimulus cycles. Retinal signals were also averaged asynchronously at 1.1 times the stimulus period to have an estimation of the intrinsic noisiness of the signal. For both the P-ERG and the noise waveforms, the response component corresponding to the contrast reversal rate (12 Hz: second harmonic of the stimulus frequency) was evaluated by discrete Fourier transform. The 12-Hz response component was also evaluated for partial estimates (50 sweeps each) of the total average to have an index of response consistency. No pupillary dilatory agents or dioptric correction was used. It is known that the eye of the pigmented mouse is emmetropic (22) and with large depth of focus due to the pinhole pupil. In addition, previous controls obtained by recording the P-ERG as a function of dioptric power of trial lenses placed before the eye, indicated zero dioptric power as suitable correction to obtain responses of maximal amplitude.
Statistics.
Group averages for both wild-type and Bcl-2 mice were statistically evaluated by two-way ANOVAs on log10 transform of raw amplitude data to approximate (P < 0.05) normal distribution/equal variances assumptions. Relevant factors were axotomy (operated vs. unoperated eyes) and interaction between axotomy and spatial frequency.
RESULTS
The visual response of RGCs has been recorded in 6 wild-type mice and 12 transgenic Bcl-2 mice. Three wild-type mice and 6 Bcl-2 mice served as intact controls, whereas the others received a unilateral optic nerve transection between optic foramen and the chiasm. Hystological analysis of whole-mounted retinae of optic nerve transected animals (10) indicated massive ganglion cell loss in axotomized wild-type mice within 2 months. By contrast, in Bcl-2 mice more than 70% of RGCs still survived 3.5 months after axotomy.
Control Mice.
A typical example of P-ERG and noise recorded in a wild-type mouse in response to gratings of 0.1 c/degree counterphased at 6 Hz, is shown in Fig. 1. The P-ERG and the noise have been averaged over 500 stimulus presentations. The P-ERG (bold-line waveform) has a sinusoidal-like waveform, whose period corresponds to the stimulus contrast reversal (12 Hz). Thus, the 12-Hz Fourier component of the P-ERG represents the main response modulation well. The polar diagram to the lower left shows the 12-Hz component of the P-ERG (vector) and that of partial averages (50 sums each) of the response (open symbols). The clustering of partial estimates indicates that the response is reliable (23) in spite of its small amplitude. Analysis of the noise response (dotted-line waveform) is shown in the polar plot to the lower right. Data points group themselves around the origin with a scattered phase, indicating that the averaged intrinsic noise (vector) is much smaller than the P-ERG. The amplitude of this asynchronous noise is comparable to that obtained by recording the P-ERG with the stimulus occluded (data not shown).
Figure 1.
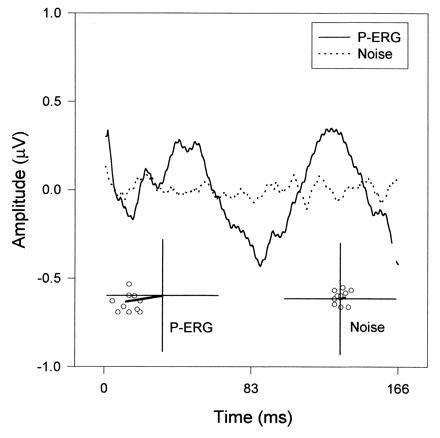
Example of P-ERG (bold line waveform) in response to a 0.1 c/degree grating counterphased at 6 Hz. The superimposed dotted-line waveform represent the asynchronous noise. Note that the P-ERG has a strong modulation at twice the stimulus period (12 Hz). The polar plots represent the Fourier 12-Hz component for either waveform (vector) as well as for their partial averages (open symbols). For each estimate, the distance from origin represents amplitude (axes are calibrated to 1 μV) and the angle represents phase. Grouping of estimates indicates response reliability.
Fig. 2 summarizes the results obtained in unoperated control mice. The P-ERG amplitude depends on the spatial frequency of the visual stimulus. In both wild-type (Fig. 2A) and Bcl-2 (Fig. 2B) mice, the P-ERG amplitude decrease with increasing spatial frequencies to reach the noise level at about 0.7 c/degree. This value is an estimate of spatial resolution of the retinal output (P-ERG acuity). The P-ERG acuity has been shown in different mammals to correspond well to the behavioral visual acuity (24–26). A resolution of the order 0.7 c/degree is in keeping with the behavioral acuity reported for mice (27).
Figure 2.
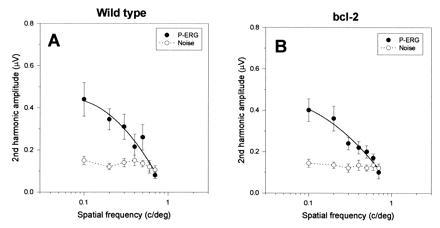
Average variation (±SEM) of the P-ERG amplitude with spatial frequency for wild-type mice (A) (n = 3) and Bcl-2 mice (B) (n = 6). In both kinds of mice the P-ERG amplitude (•) is above the noise level (○) for spatial frequencies lower than 0.7 c/degree.
Optic-Nerve-Transected Mice.
Representative waveforms of P-ERGs recorded in both wild-type and Bcl-2 mice after unilateral optic nerve section are depicted in Fig. 3. In the wild-type mouse, the P-ERG is no longer recordable in the operated eye 2 months after axotomy. This result extends to mice previous data obtained in other mammals [human (16, 19), monkey (28), cat (15, 25), and rat (26)] showing that the P-ERG depends on the normal function of RGCs, being selectively depressed in dysfunction of RGCs and abolished when they are degenerated. Contrary to wild-type mice, in the Bcl-2 mouse, the P-ERG is still recordable from the operated eye 3.5 months after axotomy, with an amplitude comparable to that of the unoperated eye.
Figure 3.
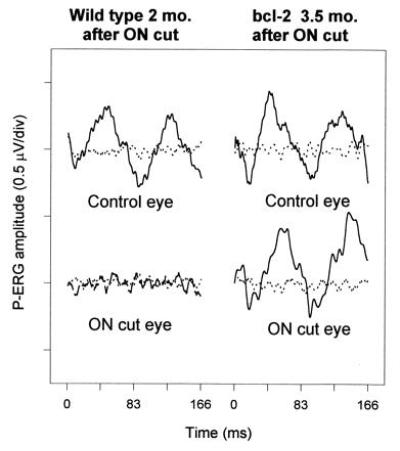
Representative examples of P-ERGs in response to gratings of 0.1 c/degree recorded in both wild-type and Bcl-2 mice after unilateral optic nerve section. Note that the response is abolished in the operated eye of the wild-type, but not Bcl-2, mouse. The ganglion cell number, evaluated from whole-mounted retinae as described (10), was for the control eyes (upper traces) 45,600 (wild type), 108,200 (bcl-2) and for the axotomized eyes (lower traces) 2100 (wild type), 75,200 (bcl-2).
Group averages are shown in Fig. 4. In the operated eyes of wild-type mice (Fig. 4A), the P-ERG is at noise level 2 months after axotomy at any spatial frequency in the range 0.1–0.5 c/degree, whereas the P-ERG recorded from the unoperated eyes is in the range of that of control wild-type mice [two-way ANOVA, effect of axotomy: F(1, 41) = 50.3, P < 0.0001]. As shown in Fig. 4B, 2 months after axotomy the P-ERG recorded in Bcl-2 mice has comparable amplitude in operated and unoperated eyes as a function of spatial frequency [two-way ANOVA; effect of axotomy, F(1, 35) = 0.088, P = 0.77; axotomy by spatial frequency, F(5, 35) = 0.66, P = 0.65]. In Bcl-2 mice, 3.5 months after axotomy (Fig. 4C), the P-ERG recorded in operated eyes is still comparable in amplitude to that of unoperated eyes in all the range of spatial frequency tested [two-way ANOVA; axotomy, F(1, 29) = 0.087, P = 0.77; axotomy by spatial frequency, F(4, 29) = 0.77, P = 0.55].
Figure 4.

Group averages (±SEM) of the P-ERG amplitude as a function of spatial frequency for both the operated eyes (○) and unoperated eyes (•) of wild-type mice 2 months after axotomy (n = 3) (A) and of Bcl-2 mice 2 months after axotomy (n = 3) (B) and 3.5 months after axotomy (n = 3) (C). In the operated eyes of wild-type mice (A), the P-ERG is at noise level (dashed lines) 2 months after axotomy at any spatial frequency in the range of 0.1–0.5 c/degree. At the same spatial frequencies, the P-ERG recorded in Bcl-2 mice 2 months (B) and 3.5 months (C) after unilateral axotomy has comparable amplitude in the two eyes.
In addition to the systematic recording of the P-ERG as a function of spatial frequency, in some cases the effect of temporal frequency and contrast has been also evaluated. Overall, responses of control wild-type, control Bcl-2, and axotomized Bcl-2 mice were rather comparable over temporal frequency and contrast. As an illustrative example, we report in Fig. 5 the P-ERG raw data as a function of stimulus contrast (the spatial frequency and temporal frequency were fixed at 0.1 c/degree and 6 Hz, respectively) obtained in one intact wild-type mouse, one intact Bcl-2 mouse, and one Bcl-2 mouse 3.5 months after optic nerve transection. In control mice [either wild-type (Fig. 5A) or Bcl-2 (Fig. 5B)], the responses decrease monotonically in amplitude with decreasing contrast to reach the noise level (dashed lines) at about 20% contrast. The amplitude of the P-ERG recorded from the axotomized eye of the Bcl-2 mouse (Fig. 5C) is in the range of that of control mice over the whole range of contrast, including the contrast threshold.
Figure 5.
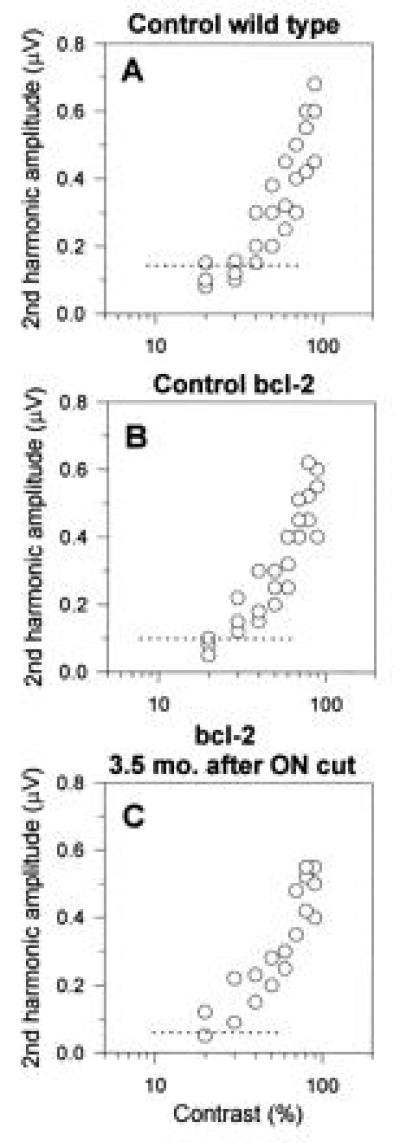
Raw amplitude data of the P-ERG recorded, at different stimulus contrasts, in one intact wild-type mouse (A), one intact Bcl-2 mouse (B), and from the eye of one Bcl-2 mouse axotomized 3.5 months before (C). The spatial frequency and temporal frequency were fixed at 0.1 c/degree and 6 Hz, respectively. In all cases, the P-ERG amplitude decreases with decreasing contrast and approaches the noise level (dotted lines) at about 20% contrast. The ganglion cell number, evaluated from whole mounted retinae as described (10), was 46,200 (A), 112,300 (B), and 70,900 (C).
A question to ask was whether the surgical procedure for optic nerve transection caused unspecific retinal dysfunction. Dysfunction of the outer retina would necessarily alter visual responses of the inner retina. To address this question, we systematically recorded, in wild-type and Bcl-2 mice, the ERG in response to an uniform field flickering at 6 Hz (flicker ERG). This response is known to originate from preganglionic retinal sources (18, 29). Flicker ERGs had typically a very simple sinusoidal waveform, with a period corresponding to the flicker rate. Response amplitudes were of the order of 5 μV (6-Hz component: data not shown) and were comparable between unoperated and operated eyes. This result indicates that our surgical prodedure did not cause unspecific retinal dysfunction to the outer retina, in agreement with the notion that preganglionic neurons such as amacrine cells and bipolar cells are not altered by optic nerve transection (e.g., ref. 30).
DISCUSSION
This study shows that Bcl-2 overexpression preserves for a long time the normal physiological response of RGCs after axotomy, which spared more than 70% of them from death (10). Contrary to wild type, in which the visually driven activity of RGCs was abolished within 2 months, in Bcl-2 mice RGC activity was still well recordable at least up to 3.5 months (recent preliminary results in Bcl-2 mice indicate that the P-ERG is recordable even 6 months after optic nerve crush). The responses were similar to those of unoperated eyes as a function of spatial frequency, and there were strong indications that responses were also comparable as a function of contrast and temporal frequency.
In both control and axotomized Bcl-2 mice, the visually driven response of RGCs was comparable to that recorded in control wild-type mice. The retinal visual acuity (which predicts to some extent the behavioral visual acuity) was also in the normal range, supporting further the finding of good physiological conditions of RGCs a long time after axotomy. The retinal acuity being of the same order in Bcl-2 and wild-type mice may be of some interest. Because of Bcl-2 overexpression during the period of naturally occurring cell death, the retina of the adult transgenic mouse has more neurons than normal (10, 31). One may wonder whether this difference does not translate in a different spatial resolution of the retinal output. It worths noticing, however, that Bcl-2 overexpression differs among retinal neuronal subpopulations (31). In particular, Bcl-2 immunoreactivity has been found in the majority of cells in the inner nuclear layer and in all cells of the ganglion cell layer. No expression has been detected in the photoreceptors. Consequently, the increase in the number of neurons occurs mainly in the inner retina (31). It is conceivable that wild-type and transgenic mice have different retinal connectivity so that their visual capabilities are difficult to predict on the basis of RGC number alone.
A number of previous studies exploited different approaches to protect RGCs from death after optic nerve transection. In particular, the administration of neurotrophic factors (1–3, 5), the transplant of embryonic tissue (7), or graft of peripheral nerve sheaths (8) has been used. Our results indicate that the overexpression of Bcl-2 represents by far the most successful tool. The mechanism of action of the Bcl-2 protein is not yet clear, even if several hypotheses have been advanced. One of the best documented properties of the Bcl-2 protein is its strong activity in interfering with apoptotic cell death (12–14). It is likely that the effects shown in the present study are exerted through this property, since RGCs have been shown to die by apoptosis after axotomy in both neonatal and adult rats (32, 33).
The importance of the present findings for regeneration studies can be seen under two possible perspectives: (i) The very high survival of neurons with normal physiological response in Bcl-2 transgenic mice offers the opportunity of studying regeneration in a system where cell death is not a limiting factor. (ii) If we were successful in developing a strategy to increase the expression of Bcl-2 (or mimic its functions) in the central nervous system, then we would gain an effective tool to counteract trauma-induced cell death. This strategy could of course benefit from additional treatments such as supply of neurotrophic factors (1–3, 5), transplant of embryonic tissue (7), and neutralization of myelin-associated neurite growth inhibitors (34, 35).
Acknowledgments
This work was supported in part by the International Institute for Paraplegia through Grant P24 and by the European Economic Community through biotech contract BIO4-CT96 0774.
Footnotes
Abbreviations: RGC, retinal ganglion cell; P-ERG, pattern electroretinogram.
References
- 1.Maffei L, Carmignoto G, Perry V H, Candeo P, Ferrari G. Proc Natl Acad Sci USA. 1990;87:1855–1859. doi: 10.1073/pnas.87.5.1855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Carmignoto G, Maffei L, Candeo P, Canella R, Comelli C. J Neurosci. 1989;9:1263–1272. doi: 10.1523/JNEUROSCI.09-04-01263.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Mansour-Robaey S, Clarke D B, Wang Y-C, Bray G M, Aguayo A J. Proc Natl Acad Sci USA. 1994;91:1632–1636. doi: 10.1073/pnas.91.5.1632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Huxlin K R, Dreher B, Schulz M, Jervie Sefton A, Bennett M R. Eur J Neurosci. 1995;7:96–107. doi: 10.1111/j.1460-9568.1995.tb01024.x. [DOI] [PubMed] [Google Scholar]
- 5.Mey J, Thanos S. Brain Res. 1993;602:304–317. doi: 10.1016/0006-8993(93)90695-j. [DOI] [PubMed] [Google Scholar]
- 6.Thanos S, Mey J, Wild M. J Neurosci. 1993;13:455–466. doi: 10.1523/JNEUROSCI.13-02-00455.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Gravina A, Domenici L, Berardi N, Maffei L. Exp Brain Res. 1990;80:631–634. doi: 10.1007/BF00228004. [DOI] [PubMed] [Google Scholar]
- 8.Vidal-Sanz M, Bray G M, Villegas-Peres M P, Thanos S, Aguayo A J. J Neurosci. 1987;7:2894–2909. doi: 10.1523/JNEUROSCI.07-09-02894.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Cenni M C, Strettoi E, Bonfanti L, Ratto G M, Rabacchi S, Martinou J C, Maffei L. Soc Neurosci Abstr. 1995;21:1556. [Google Scholar]
- 10.Cenni M C, Bonfanti L, Martinou J C, Ratto G M, Strettoi E, Maffei L. Eur J Neurosci. 1996;8:1735–1745. doi: 10.1111/j.1460-9568.1996.tb01317.x. [DOI] [PubMed] [Google Scholar]
- 11.Dubois-Dauphin M, Frankowski H, Tsujimoto Y, Huarte J, Martinou J-C. Proc Natl Acad Sci USA. 1994;94:3309–3313. doi: 10.1073/pnas.91.8.3309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Osborne B S, Schwartz L M. Trends Cell Biol. 1994;4:394–398. doi: 10.1016/0962-8924(94)90052-3. [DOI] [PubMed] [Google Scholar]
- 13.Korsmeyer S J. Trends Genet. 1995;11:101–105. doi: 10.1016/S0168-9525(00)89010-1. [DOI] [PubMed] [Google Scholar]
- 14.Davies A M. Trends Neurosci. 1995;18:355–358. doi: 10.1016/0166-2236(95)93928-q. [DOI] [PubMed] [Google Scholar]
- 15.Maffei L, Fiorentini A. Science. 1981;211:953–955. doi: 10.1126/science.7466369. [DOI] [PubMed] [Google Scholar]
- 16.Zrenner E. Prog Retinal Res. 1990;9:427–476. [Google Scholar]
- 17.Sieving P A, Steimberg R H. J Neurophysiol. 1987;57:104–120. doi: 10.1152/jn.1987.57.1.104. [DOI] [PubMed] [Google Scholar]
- 18.Baker C L, Hess R F, Olsen B T, Zrenner E. J Physiol (London) 1988;407:155–176. doi: 10.1113/jphysiol.1988.sp017408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Berninger, T. & Arden, G. B. (1988) Eye 2, Suppl., 257–283. [DOI] [PubMed]
- 20.Kothe A C, Lovasik J V. Electroenceph Clin Neurophysiol. 1990;75:185–199. doi: 10.1016/0013-4694(90)90172-g. [DOI] [PubMed] [Google Scholar]
- 21.Colotto A, Falsini B, Porciatti V, Fadda A, Giudiceandrea A, Salgarello T. Invest Ophthalmol Visual Sci. 1995;36:S332. [Google Scholar]
- 22.Remtulla S, Hallet P E. Vision Res. 1985;25:21–31. doi: 10.1016/0042-6989(85)90076-8. [DOI] [PubMed] [Google Scholar]
- 23.Porciatti V, Burr D C, Morrone C, Fiorentini A. Vision Res. 1992;32:1199–1209. doi: 10.1016/0042-6989(92)90214-4. [DOI] [PubMed] [Google Scholar]
- 24.Fiorentini A, Pirchio M, Sandini G. Hum Neurobiol. 1984;3:939–951. [PubMed] [Google Scholar]
- 25.Maffei L, Fiorentini A. Exp Brain Res. 1982;48:327–334. doi: 10.1007/BF00238608. [DOI] [PubMed] [Google Scholar]
- 26.Berardi N, Domenici L, Gravina A, Maffei L. Exp Brain Res. 1990;79:539–546. doi: 10.1007/BF00229323. [DOI] [PubMed] [Google Scholar]
- 27.Sinex D, Burdette L J, Pearlman A L. Vision Res. 1979;19:853–858. doi: 10.1016/0042-6989(79)90018-x. [DOI] [PubMed] [Google Scholar]
- 28.Maffei L, Fiorentini A, Bisti S, Holländer H. Exp Brain Res. 1985;59:423–425. doi: 10.1007/BF00230925. [DOI] [PubMed] [Google Scholar]
- 29.Porciatti V, Falsini B. Clin Vision Sci. 1993;8:435–447. [Google Scholar]
- 30.Perry V H, Henderson Z, Linden R. J Comp Neurol. 1983;219:3546–3568. doi: 10.1002/cne.902190309. [DOI] [PubMed] [Google Scholar]
- 31.Martinou J-C, Dubois-Dauphin M, Staple J, Rodriguez I, Frankowski H, Missoten M, Albertini P, Talabot D, Catsicas S, Pietra C, Huarte J. Neuron. 1994;13:1017–1030. doi: 10.1016/0896-6273(94)90266-6. [DOI] [PubMed] [Google Scholar]
- 32.Rabacchi S A, Bonfanti L, Liu X-H, Maffei L. J Neurosci. 1994;14:5292–5301. doi: 10.1523/JNEUROSCI.14-09-05292.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Berkelaar M, Clarke D B, Wang Y-C, Bray G M, Aguayo A J. J Neurosci. 1994;14:4368–4374. doi: 10.1523/JNEUROSCI.14-07-04368.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Caroni P, Schwab M E. Neuron. 1988;1:85–96. doi: 10.1016/0896-6273(88)90212-7. [DOI] [PubMed] [Google Scholar]
- 35.Weibel D, Cadelli D, Schwab M E. Brain Res. 1994;642:259–266. doi: 10.1016/0006-8993(94)90930-x. [DOI] [PubMed] [Google Scholar]