

Abstract
Functional neuroimaging studies investigating sex differences in visuospatial processing traditionally focus on mental rotation tasks in adults, as it is a consistently robust finding, with a limited number of studies examining tasks tapping visuospatial skills at a more basic level. Furthermore, fewer studies have examined this issue in conjunction with investigating whether differences exist in younger populations. Therefore, functional neuroimaging was used to examine whether sex-based differences exist and/or develop during childhood. Thirty-two participants, matched on performance, participated in this study. Overall, both groups showed overlapping activation in bilateral superior parietal lobe, extrastriate cortex, and cerebellum; differences between the sexes showed that males had significantly greater activation in right lingual gyrus and cerebellum. Formal comparisons between age groups revealed that older males show engagement of left hemisphere regions, while females show greater bilateral (R > L) engagement of regions traditionally associated with visuospatial processing. Together, these results suggest that older males, as compared to younger males, may engage regions that are associated with a visuomotor network, whereas females utilize areas indicated in spatial attention and working memory. Furthermore, these results could also suggest that there may be differences in strategy use that are evident early on and may continue to develop over time evident by differential engagement of networks associated with visuospatial processing. Our data provide evidence for sex-based differences in the neural basis of visuospatial processing.
Keywords: Visuospatial processing, Sex differences, JLO, Judgment of line orientation
Sex differences in processing and performance within the visuospatial domain have been of great interest. Generally, tasks involving spatial transformations (i.e., mental rotation) produce the most robust sex-based differences, with males outperforming females; however, differences have also been reported on spatial perception/visualization tasks [14]. Most studies have typically addressed the issue of identifying and understanding the neurobiology of sex-based differences using mental rotation tasks as these tend to produce robust results [2,6-9,12,15]. Additionally, a limited number of studies have utilized more basic perceptual tasks, such as spatial line orientation tasks [3-5], to assess whether differences are evident at a more basic level of processing and not only during greater cognitive demands. Typically, studies have examined this issue in adults [2,6-8,12,15], while only a few have investigated younger populations [9,11]. However, examining sex-based differences in visuospatial processing in younger populations (e.g., children) could provide further insights into the maturation of important regions involved in visuospatial processing as well as to identify potential differences in strategies used, and the development of these strategies, during task completion.
Across all adult studies investigating visuospatial processing, a network of activation within the frontal, occipital, and parietal lobes is reported [2,3,6-8,12,13,15]. While most studies specifically designed to assess sex-based differences have found that differences exist, the nature of the precise difference has yielded inconsistent results [2-5,7,8,15]. Additionally, a limited number of studies have failed to show any sex difference, attributing such differences to performance-related factors [13]. A few studies have primarily focused attention on the lateralization of activation [3-5], via a region of interest (ROI) analysis. ROIs within the inferior parietal lobe have shown that males exhibit a bilateral representation of activation while females show greater right lateralization; these studies were the only ones to utilize a more basic perceptual task such as spatial line orientation. The remaining studies have all utilized some variant of a mental rotation task. Weiss et al. (2003) showed a distinction between inferior and superior parietal lobe, with greater activation in inferior parietal lobe for males and greater activation in superior parietal lobe for females. Other studies have shown that in addition to parietal lobe activation, females also tend to show additional activation in right frontal regions when compared to males [7,15].
In addition to looking at differences in lateralization and recruitment of parietal and frontal lobe regions, other studies examining sex-based differences have focused attention on how strategy-related differences may be linked to functional-related differences in visuospatial processing. In 2002, Jordan and colleagues showed that females had greater activation than males in bilateral inferior temporal gyrus and premotor regions as well as right superior parietal and left inferior parietal lobe, whereas males had greater activation than females in bilateral primary motor, left posterior intraparietal sulcus, and right parieto-occipital sulcus. It was concluded that differences in activation were due to different strategies used to complete the task with females utilizing a verbal-analytic (“piecemeal”) strategy and males a visuomotor (“gestalt”) strategy. Furthermore, Butler et al. (2006) showed that females had greater activation than males in bilateral dorsomedial prefrontal regions, right inferior temporal gyrus and left inferior occipital lobe, whereas males had greater activation in bilateral postcentral gyrus and left hemisphere regions to include paracentral gyrus, precuneus, basal ganglia. However, Butler et al. concluded that the results indicated that females engaged areas important for spatial working memory suggesting more effortful “top–down” processing while males utilized a visuomotor network reflecting more efficient “bottom–up” processing.
Moreover, a limited number of studies have investigated the neurobiology of sex differences as related to development in visuospatial processing in younger populations. Kucian and colleagues (2007) conducted a study comparing activation associated with a mental rotation task in children and adults. Both groups had activation in right fusiform gyrus and bilateral parietal lobe—superior parietal for the child group (R > L) and inferior parietal for the adult group; adults had additional activation in bilateral extrastriate cortex. The child group showed additional activation when compared to adults in bilateral posterior cingulate and precuneus while adults had greater activation than children did in left intraparietal sulcus. There were no sex differences in the child group while adult females showed stronger activation in right middle temporal gyrus, right inferior frontal gyrus, and left primary motor. Thus, this study shows that there is a developmental shift in the intraparietal sulcus from stronger right lateralization in younger groups to a bilateral representation in adults. Schweinsburg et al. (2005) conducted a study investigating the impact of age and sex on spatial working memory. They showed that there was a significant positive correlation with age in a left prefrontal region and bilateral inferior posterior parietal lobe; a significant negative correlation with age was seen in bilateral superior parietal cortex. Moreover, females demonstrated a diminished response in anterior cingulate activation while males demonstrated a greater response in frontopolor cortex compared to females. Schweinsburg et al. concluded that frontal and parietal neural networks that are involved in spatial working memory change during adolescences, influenced by sex, which possibly reflects the development of sex-specific strategies used for task completion.
The objective of the present study was to test whether there are differences in brain activation between males and females during a more basic visuospatial processing task, rather than tasks requiring greater cognitive demands, while also using a cross-sectional design across mid-childhood into adolescence to assess for possible developmental differences. Specifically, we hypothesized that with increasing age, males would develop a bilateral representation of visuospatial processing in parietal regions shown by the utilization of right hemisphere regions in younger-aged participants and the engagement of left hemisphere regions later in adolescent males. For females, we predicted that we would see a consistent right hemisphere network in frontal and parietal regions regardless of age. We used a spatial line orientation task, previously shown to elicit sex-based differences [3-5], that is less cognitively demanding than mental rotation [10]. More specifically, this task was chosen as previously research in our lab [3] and others [4,5] has revealed differences between the sexes in lateralization, and we wanted to know if these same differences in basic visuospatial processing were present in children and adolescents. Additionally, because the task was less demanding than mental rotation tasks, this allowed us to easily control for performance; thus, any difference in activation patterns would not be confounded and attributed to differing performance levels, ensuring that the results could be attributed to sex-based differences, and not performance. Furthermore, utilization of a population of children with varying ages, in conjunction with a more basic visuospatial task, could pave the way for understanding the development of areas important for visuospatial processing, as well as the identification of potential differences in strategies used during task completion. Additionally, both groups were equated on age.
Thirty-two children participated in the study between 7 and 15 years of age (16 males: 10.150 ± 2.551, 16 females: 10.350 ± 2.597). Before entering the study, parents were briefly interviewed to ensure that the following criteria were met: (1) were native English speakers, (2) had normal hearing and normal/corrected-to-normal vision, (3) had no history of a developmental disability, major psychiatric illness, and/or neurological disorder, (4) were not taking medications, and (5) had no contraindication to the MRI environment. For those participants who met criteria, written consent was obtained (children signed an additional assent statement) and procedures were carried out in accordance with the Johns Hopkins Medical Institutional Review Board.
The task was an adapted version of the judgment of line orientation (JLO) [1], consisting of a fan of 11 lines displayed at the bottom of the screen. All of the lines were displayed in blue except for two lines that were yellow. Displayed above the fan was a pair of yellow lines that were oriented in either the same or different position as the two yellow lines in the fan. Participants pushed a button with their right index finger if the top and bottom yellow lines were oriented in the same position and pushed a button with their left index finger if the lines were oriented in different positions. The control task (color) was a color discrimination task, controlling for basic visual discrimination abilities. During the color task, participants were asked to decide if the lines above the fan, either displayed as blue or yellow, were the same color as the fan of 11 lines, which were always displayed as blue. When the colors were the same, participants pressed the button with their right index finger, and when they were not, with their left.
For the task, each block (JLO or control) was approximately 50 s, consisting of 10 stimuli with each stimulus on the screen for 4 s and had an ISI of 1 s. A 3 s instruction screen and a 2 s rest were also included between each block. The timing parameters were the same for both tasks. Two runs were presented, with each run lasting 5 min, 40 s and consisting of six alternating blocks of each stimulus type (three JLO in each run). An LCD projector was used to project all stimuli onto a rear-projection screen that participants viewed via a mirror fixed atop the MRI head coil. The paradigm was computer-controlled with E-Prime (Psychology Software Tools, Pittsburgh, PA, USA), which presented the task and recorded the timing of both stimulus presentation and participant response.
Scanning was carried out in a 1.5T ACS-NT Powertrack 6000 MRI scanner (Philips Medical Systems, Inc.) using body coil transmission and quadrature end-capped head coil reception. Functional MRI data were acquired using single shot echo planar imaging with an echo time = 40 ms, repetition time = 2.6 s, 64 × 64 matrix, and 230 mm field of view. Across the 128 images, 40 4.0 mm thick coronal slices with 0.5 mm inter-slice gaps provided whole brain coverage.
Image analysis was performed using statistical parametric mapping (SPM2; http://www.fil.ion.ucl.ac.uk/spm/) on Matlab 6.5 (Mathworks, Inc., Natick, MA, USA). For each participant, images from the scanner were converted to Analyze format, time corrected to adjust for within volume time of acquisition differences, realigned, spatially normalized to Montreal Neurological Institute (MNI)-labeled space, re-sampled into (2 mm)3 voxels, and smoothed using a 8 mm3 Gaussian kernel. At the individual level, task associated activation was assessed using a block design and SPMs were created corresponding with the time-course for the JLO task contrasted to the color task. Voxel-wise t-maps were constructed for each of the subjects as a first-level analysis and were taken to a second-level analysis with subjects as a random factor. Both groups were broken down into two separate age groups (division at 10 years of age). Data were entered into a 2 (sex) × 2 (age) ANOVA to test not only for the main effect of sex, but also to test for significant effects of age between males and females separately. Lastly, simple correlations with age as the regressor of interest were run for each sex separately. All images, presented in radiological format, had a cluster-based threshold of uncorrected threshold of P < 0.005, with an extent threshold of 150 voxels, which is equivalent to a false positive rate of P < 0.05 over the whole brain based on Monte Carlo simulations. The locations of significant voxels were summarized by their local maxima, separated by 4 mm, converted from MNI to Talairach coordinate space, and then assigned neuroanatomic and cytoarchitectonic labels using the Talairach Daemon (http://ric.uthscsa.edu/projects/talairachdaemon.html).
Mean response time and task accuracy (in percentage correct) were measured1. Overall, there was a high level of performance for males (86.893 ± 8.381) and females (86.785 ± 5.390), with roughly equivalent reaction times (males: 1678.762 ± 202.437; females: 1551.246 ± 293.279). Thus, sex differences in performance (task accuracy: P = 0.967; response time: P = 0.178) were not found.
Males and females showed overlapping areas of activation in bilateral superior parietal lobe (BA 7), cerebellum, lingual gyrus (BA 17/18), and middle occipital gyrus (BA 18/19/37); additional activation seen in right middle/inferior frontal gyri (BA 9/46 males only), right cuneus/precuneus (BA 19/7, respectively), and left fusiform and inferior occipital gyri (Fig. 1).
Fig. 1.

SPM maps showing areas significantly activated by the JLO task for females (A) and males (B).
For all between group comparisons, see Fig. 2. There was a significant main effect of sex in right lingual gyrus (BA 18) and cerebellum; subsequent analyses showed that males, irrespective of age, had significantly greater activation than females in these regions (Fig. 2A). For the comparisons between the younger and older participants for each sex separately, results showed there were no significant differences in activation patterns for younger as compared to older participants. However, there were significant differences seen, for both sexes, in the older versus the younger participants. For males, greater activation was seen in frontal regions including bilateral middle frontal gyrus and left precentral gyrus, as well as posterior regions including bilateral lingual gyrus, right cuneus (BA 17), and left inferior occipital gyrus (Fig. 2C). For females, greater activation was seen in parietal regions: bilateral inferior parietal lobe (BA 40) and left superior parietal lobe (BA 7), left cerebellum, and additional right posterior regions: fusiform gyrus (BA 37), middle occipital gyrus, and middle temporal gyrus (Fig. 2B).
Fig. 2.

SPM map showing significant differences between males and females regardless of age (A); between older and younger females (B); and between older and younger males (C).
For males, age had significant positive correlations within left hemisphere regions only (P < .001). There were several posterior regions including inferior occipital gyrus (BA 17), cerebellum, inferior parietal lobe (BA 40), superior parietal lobe including precuneus (BA 7; see Fig. 3), and middle frontal gyrus (BA 6). Moreover, males showed negative correlations within right hemisphere regions (P < .001) including postcentral gyrus, middle/superior temporal gyri (BA 38–superior), and inferior parietal lobe.
Fig. 3.
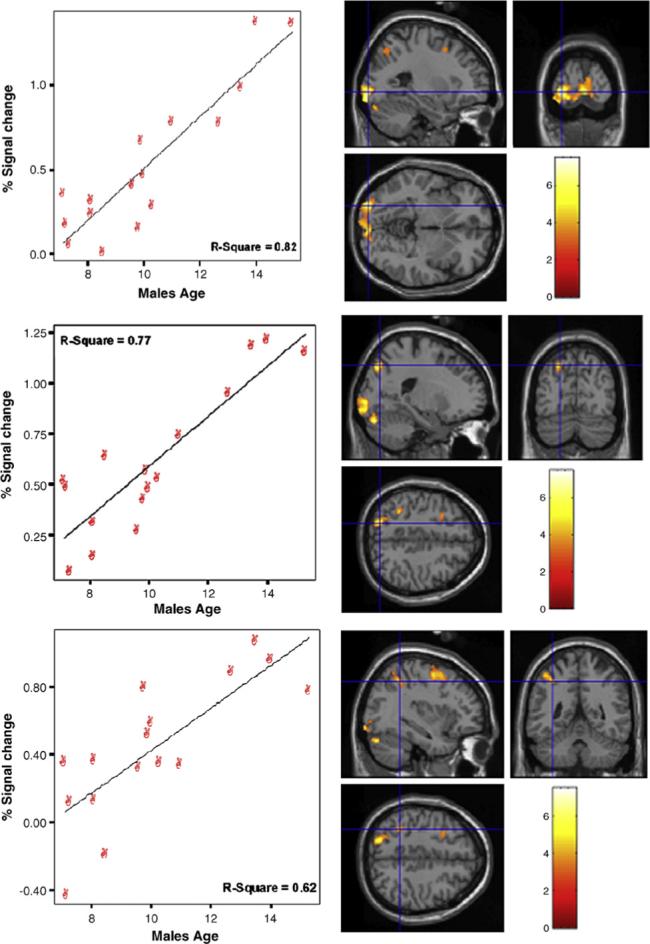
Areas indicating a significant positive correlation with age for males within left posterior cortex. The top panel shows areas within the lingual gyrus (BA 17) and cerebellum; middle panel includes inferior parietal lobe (BA 40); bottom panel includes superior parietal lobe (BA 7).
However, for females, age had significant positive correlations with more diffuse areas of activation within left and right hemispheres (P < .001). Within the right hemisphere, age was significantly correlated with middle/inferior frontal gyri (BA 46 and 9, respectively), inferior temporal gyrus (BA 20/37), and inferior parietal lobe (BA 40; see Fig. 4). Age also had significant correlations with left lingual gyrus and cerebellum. Lastly, there was a bilateral area of activation, superior parietal lobe (including precuneus), which had a significant positive correlation with age (R > L). Furthermore, for females age correlated negatively (P < .001) in right anterior cingulate gyrus (BA 32) streaming into medial frontal gyrus and left angular gyrus (BA 39), precuneus (BA 19) and cingulate cortex (BA 31).
Fig. 4.
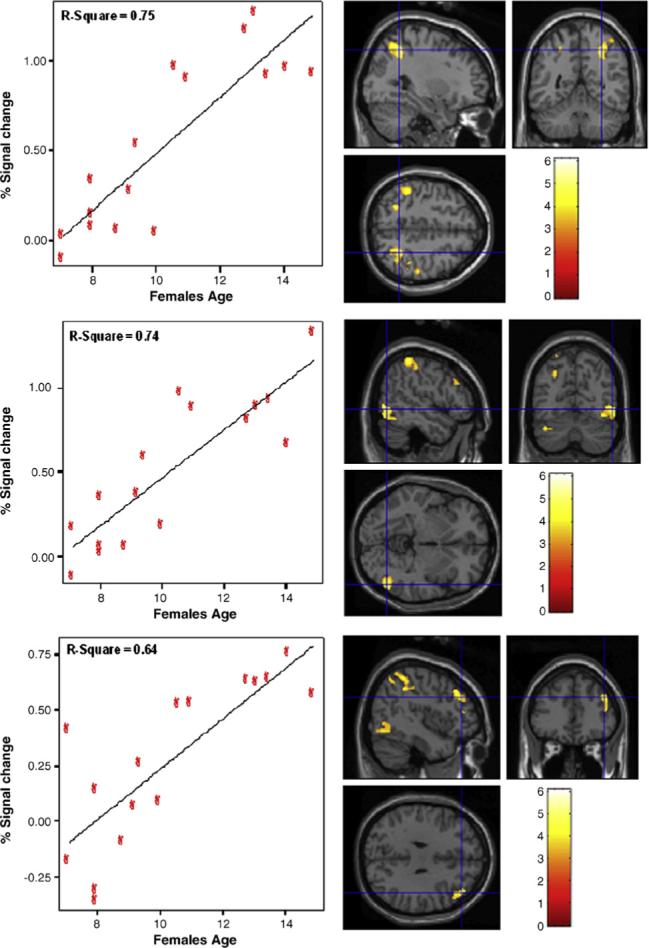
Areas indicating a significant positive correlation with age for females within right-hemisphere regions. The top panel shows areas within the inferior parietal lobe (BA 40); middle panel includes inferior temporal gyrus (BA 20/37); bottom panel includes middle and inferior frontal gyri (BA 46 and 9, respectively).
Overall, consistent with previous literature, both groups had overlapping areas of bilateral activation within superior parietal lobe, occipital lobe (inferior and middle occipital gyri), and cerebellum with additional activation seen in right dorsolateral prefrontal cortex [3-5]. However, the only between sex difference was noted in males having significantly greater activation in right lingual gyrus (BA17) and cerebellum than females. The cerebellum is a region associated with motor planning and sequencing, whereas the lingual gyrus has been associated with the encoding/decoding of spatial memories. Therefore, this finding suggests that males utilize regions to a greater degree than females implicated in visuomotor processing [7,8,12]. Between ages, it was shown that older females had greater activation than younger females in a more traditional visuospatial processing network including such regions as bilateral inferior parietal lobe, left superior parietal lobe, left cerebellum, and additional right posterior regions including fusiform gyrus and middle temporal gyrus. The additional activation in left superior parietal lobe has been found in other studies investigating sex differences (see [15]). For older males as compared to younger males, additional activation was seen in primarily in bilateral middle frontal gyrus and bilateral lingual gyrus. Together, these findings could indicate that males are more reliant upon a visuomotor network while females utilize areas consistently seen in spatial attention and spatial working memory, both of which have been associated with visuospatial processing.
Further differences in the development of processing visuospatial information are confirmed through the correlation data (within each group separately). With increasing age, males shift from traditional right hemisphere network (including IPL) to engagement of the left hemisphere; this is consistent with previous literature [3-5]. These findings suggest that an efficient right hemisphere network is developed at an early age for males and that with increasing age the left hemisphere is recruited and both hemispheres are utilized in a parallel fashion. Furthermore, these results could indicate that regions within the left hemisphere, for males, are part of an efficient visuospatial and/or visuomotor network. However, a different developmental pattern is seen in females. With increasing age, females shift from the engagement of bilateral medial frontal and posterior regions (e.g., medial frontal gyrus and cingulate) to more traditional areas seen in visuospatial processing (e.g., rIPL, rITG, rDLPFC, bilateral SPL) [2,15]. Findings suggest that younger females recruit regions typically seen in decision-making and possibly verbal strategy use and as females get older, greater activation is seen in right hemisphere regions. Specifically, activation in females is seen in more traditional areas associated with visuospatial processing, in particular, spatial attention and working memory. However, a weakness of this study is that a cross-sectional design was used as opposed to a longitudinal design. Additional studies are needed to investigate whether the differences seen truly represent development over time and the factors that may mediate this change in processing.
In summary, over time males show a more integrated visuomotor/visuospatial network while females show the development of a more spatial attention/working memory system. The differences in activation patterns in females could be mediated by a verbal strategy used early in development [7,8]. Additionally, differences seen in males are consistent with visually based strategy use in which it is suggested that males rely on imagery and are “more hands-on” during task completion [2,6-8,12]. Moreover, males’ reliance on a visuomotor network could account for the traditional advantage on visuospatial tasks. Lastly, the differential engagement of different networks associated with visuospatial processing between males and females seen with more complex tasks like mental rotation [2] is supported on this more basic perceptual task; thus, this could reflect a true sex-based difference in visuospatial processing.
Acknowledgements
This work was supported in part by the Johns Hopkins School of Medicine General Clinical Research Center (NIH grant M01-RR00052), U.S. Congressionally Directed Materiel and Medical Command (DAMD17−00−1−0548), the National Institute of Neurological Disorders and Stroke (NIH grant RO1 NS049096), and the F.M. Kirby Research Center (NIH/NCRR grant P41RR15241).
Footnotes
There was missing behavioral data for one male and one female due to computer error. Analyses were conducted with and without these participants and there were no changes to the data; thus, it was determined not to exclude these individuals.
References
- 1.Benton AL, Hamsher KD, Varney NR, Spreen O. Contributions to Neuropsychological Assessment. Oxford University Press; New York: 1983. Judgment of line orientation. [Google Scholar]
- 2.Butler T, Imperato-McGinley J, Pan H, Voyer D, Cordero J, Zhu Y-S, Stern E, Silbersweig D. Sex differences in mental rotation: Top-down versus bottom-up processing. Neuroimage. 2006;32:445–456. doi: 10.1016/j.neuroimage.2006.03.030. [DOI] [PubMed] [Google Scholar]
- 3.Clements AM, Rimrodt SL, Abel JR, Blankner JG, Mostofsky SH, Pekar JJ, Denckla MB, Cutting LE. Sex differences in cerebral laterality of language and visuospatial processing. Brain Lang. 2006;98:150–158. doi: 10.1016/j.bandl.2006.04.007. [DOI] [PubMed] [Google Scholar]
- 4.Gur RC, Alsop D, Glahn D, Petty R, Swanson CL, Maldjian JA, Turetsky BI, Detre JA, Gee J, Gur RE. An fMRI study of sex differences in regional activation to a verbal and a spatial task. Brain Lang. 2000;74:157–170. doi: 10.1006/brln.2000.2325. [DOI] [PubMed] [Google Scholar]
- 5.Gur RC, Raglan JD, Resnick SM, Skolnick BE, Jaggi J, Muenz L, Gur RE. Lateralized increases in cerebral blood flow during performance of verbal and spatial tasks: relationship with performance level. Brain Cogn. 1991;24:244–258. doi: 10.1006/brcg.1994.1013. [DOI] [PubMed] [Google Scholar]
- 6.Halari R, Sharma T, Hines M, Andrew C, Simmons A, Kumari V. Comparable fMRI activity with differential behavioural performance in mental rotation and overt verbal fluency tasks in healthy men and women. Exp. Brain Res. 2006;169:1–14. doi: 10.1007/s00221-005-0118-7. [DOI] [PubMed] [Google Scholar]
- 7.Hugdahl K, Thomsen T, Ersland L. Sex differences in visuo-spatial processing: an fMRI study of mental rotation. Neuropsychologia. 2006;44:1575–1583. doi: 10.1016/j.neuropsychologia.2006.01.026. [DOI] [PubMed] [Google Scholar]
- 8.Jordan K, Wüstenberg T, Heinze HJ, Peters M, Jäncke L. Women and men exhibit different cortical activation patterns during mental rotation tasks. Neuropsychologia. 2002;40:2397–2408. doi: 10.1016/s0028-3932(02)00076-3. [DOI] [PubMed] [Google Scholar]
- 9.Kucian K, von Aster M, Loenneker T, Dietrich T, Mast FW, Martin E. Brain activation during mental rotation in school children and adults. J. Neural Transm. 2007;114:675–686. doi: 10.1007/s00702-006-0604-5. [DOI] [PubMed] [Google Scholar]
- 10.Ng VW, Bullmore ET, de Zubicaray GI, Cooper A, Suckling J, Williams SCR. Identifying rate-limiting nodes in large-scale cortical networks for visuospatial processing: an illustration using fMRI. J. Cogn. Neurosci. 2001;13:537–545. doi: 10.1162/08989290152001943. [DOI] [PubMed] [Google Scholar]
- 11.Schweinsburg AD, Nagel BJ, Tapert SF. fMRI reveals alteration of spatial working memory networks across adolescence. J. Int. Neuropsychol. Soc. 2005;11:631–644. doi: 10.1017/S1355617705050757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Thomsen T, Hugdahl K, Ersland L, Barndon R, Lundervold A, Smievoll AI, Roscher BE, Sundberg H. Functional magnetic resonance imaging (fMRI) study of sex differences in a mental rotation task. Med. Sci.Monit. 2000;6:1186–1196. [PubMed] [Google Scholar]
- 13.Unterrainer J, Wranek U, Staffen W, Gruber T, Ladurner G. Lateralized cognitive visuospatial processing: is it primarily gender-related or due to quality of performance? A HMPAO-SPECT study. Neuropsychobiology. 2000;41:95–101. doi: 10.1159/000026639. [DOI] [PubMed] [Google Scholar]
- 14.Voyer D, Voyer S, Bryden MP. Magnitude of sex differences in spatial abilities: A meta-analysis and consideration of critical variables. Psychol. Bull. 1995;117:250–270. doi: 10.1037/0033-2909.117.2.250. [DOI] [PubMed] [Google Scholar]
- 15.Weiss E, Siedentopf CM, Hofer A, Deisenhammer EA, Hoptman MJ, Kremser C, Goalszewski S, Felber S, Fleischhacker WW, Delazer M. Sex differences in brain activation pattern during a visuospatial cognitive task: A functional magnetic resonance imaging study in healthy volunteers. Neurosci. Lett. 2003;344:169–172. doi: 10.1016/s0304-3940(03)00406-3. [DOI] [PubMed] [Google Scholar]