

Abstract
Bone marrow derived progenitor cells were reported to be involved in the pathogenesis of pterygium and have been suggested to be important in angiogenesis and the repair process after tissue damage. In order to investigate the involvement of these cells in wound healing after a pterygium excision, immunohistochemical staining was performed with a temporary amniotic membrane, applied to the bare sclera, after a pterygium excision using various progenitor cell markers, including CD34, c-kit, STRO-1, and AC133, to determine the expression levels of the participating cells. CD34-positive cells were observed along with some round or spindle-shaped mononuclear cells on the stromal side of the amniotic membrane. Some CD34-positive, large, and round or spindle-shaped cells formed clusters resembling small vessels in some regions of the amniotic membrane. c-kit was expressed in the epithelium that had grown over the amniotic membrane and in the spindle-shaped or round mononuclear cells in the stroma. Many stellate- to spindle-shaped fibroblast like cells expressed STRO-1, and AC133 was expressed in some round and ovoid cells. Overall, these results suggest that adult bone marrow-derived progenitor cells, such as endothelial progenitor cells and mesenchymal stem cells, are involved in the wound healing process post-excision in patients with pterygium.
Keywords: Amniotic membrane, progenitor cells, pterygium, wound healing
INTRODUCTION
Pterygium is a chronic condition, characterized by the invasion of an altered ocular surface tissue into the normal cornea.1 Pterygium tissue consists of a superficial conjunctival epithelial layer and an underlying fibrovascular component of hypertrophic and elastotic, degenerated connective tissue.2 The pathogenesis of pterygium is not well-understood. Many causative environmental factors may induce ocular inflammation, and various types of cytokines may cause abnormal cell changes of the ocular surface.3 Recent reports indicate that stem cell-related growth factors, such as TGF-β, VEGF, and stem cell factor (SCF), are expressed at abnormally high levels in patients with pterygium.4,5 In addition, we previously reported that bone marrow-derived stem and progenitor cells are involved in the pathogenesis of pterygium.6
In addition to being involved in the pathogenesis of pterygium, these stem cells may also promote the wound healing process either directly or indirectly by activating many growth factors and cytokines. Several studies have provided evidence that bone marrow-derived progenitor cells are crictically required for angiogenesis and repair processes after tissue damage, such as in patients with lung or in ischemic heart disease.7,8 Revascularization during healing does not rely exclusively on the sprouting of pre-existing vessels but also involves circulating endothelial cells, derived from bone marrow, called endothelial progenitor cells.9 Likewise, human hematopoietic and mesenchymal progenitor cells can transdifferentiate into multiple lineages of tissue and display structural and functional activity.10 Initial injury and acute inflammatory response can result in the emigration of these bone marrow-derived progenitor cells into the circulation, their migration to the damaged site, and subsequent differentiation. Therefore, we hypothesize that these cells could be involved in wound healing after pterygium removal.
Numerous studies have demonstrated that the human hematopoietic progenitor cell antigen, CD34, is synthesized and expressed by certain cells of the hematopoietic lineage and is a marker of endothelial precursor tissue and differentiated endothelium.11,12 c-kit, used as a cell marker, has been identified as a member of the receptor-tyrosine kinase family. The c-kit receptor binds to human stem cell factor and is involved in the development of several lineages of stem cells, such as germ cells and hematopoietic precursor cells, as well as in erythropoiesis and mast cell proliferation.13 AC133 is a stem cell antigen that may be useful for the selection of hematopoietic cells and vascular endothelium and may also be used as a positive marker for the characterization of the trophoblast cell line.14 The majority of cells reactive to AC133 antibodies have been found to co-express CD34 and c-kit.15 STRO-1 antigen is expressed by human bone marrow fibroblast-like cells and by various nonhematopoietic progenitor cells.16 On the basis of these reports, we performed immunohistochemical staining with a temporary amniotic membrane, applied to the bare sclera after pterygium excision, using various progenitor cell markers to investigate the involvement of bone marrow-derived progenitor cells in the wound healing process.
MATERIALS AND METHODS
Patient and specimen collection
Thirty patients with primary or recurrent pterygium were selected for this study (primary pterygium, n = 15, recurrent pterygium, n = 15, mean age 62.7 ± 5.8 and 63.6 ± 4.9, respectively). All subjects underwent a detailed bio-microscopic examination to exclude ocular surface disorders, and all operations were performed by the same surgeon. Briefly, after conventional bare sclera excision of the pterygium, a preserved human amniotic membrane was properly designed and trimmed with Westcott scissors, and applied stromal-side-down. To prevent the amniotic membrane from being dislodged by nictitating movement, the patch margins were placed underneath the free edges of the conjunctiva. The amniotic membrane was then firmly attached to the exposed sclera, using a needle holder. Four days postoperatively, the amniotic membrane patches were removed, using a fine tissue forceps and formalin embedded or snap-frozen. Serial cross sections were made along the longitudinal axis or the membrane was flat-mounted. All procedures followed the tenets of the Helsinki declaration, and written informed consent was obtained from all patients.
Immunohistochemical study
Serially sectioned (2-4 um) specimens were evaluated by hematoxylin and eosin staining for routine analysis and then processed immunohistologically, as previously described.17 Briefly, paraffin sections were deparaffinized in xylene, rehydrated, and quenched for endogenous peroxidase. Cryostat sections were placed on gelatinized slides and fixed in cold acetone. Tissue sections were equilibrated in Tris-buffered saline (TBS) and blocked in non-immuno serum (Histostain-plus kit, Zymed Laboratories, South San Francisco, CA, USA). Sections were then incubated with polyclonal goat anti-human CD34 antibody (N19, Santa Cruz Biotechnology, Santa Cruz, CA, USA), polyclonal goat anti-human c-kit antibody (C-14, Santa Cruz Biotechnology, Santa Cruz, CA, USA), and monoclonal mouse anti-human antibodies AC133(Clone AC133 epitope1, Miltenyi Biotech, Bergisch Gladbach, Germany) and STRO-1(Developmental Studies Hybridoma Bank, Iowa, IA, USA) overnight at 4℃. Sections were extensively washed in TBS before adding a biotinylated secondary antibody. Then the sections were rewashed and incubated for 1 hour with peroxidase-conjugated streptavidin. The presence of peroxidase was revealed by adding a substrate-chromogen (3-amino-9-ethycarbazole) solution. The sections were then counterstained with haematoxylin.
Each section was examined under a microscope (Carl Zeiss, Jena, Germany) and photographed. The number of CD34-, c-kit-, AC133-, and STRO-1-positive cells per high power (40 × objective) field (HPF) was counted from three different regions of each sample, and counts were averaged among regions for each sample. Statistical analysis was performed, using SPSS version 11.0 for Windows (SPSS Inc, Chicago, IL, USA).
RESULTS
On the first postoperative day after pterygium excision, using a bare sclera technique with a temporary amniotic membrane patch, we observed firm attachment of the amniotic membrane to the sclera (Fig. 1). On the fourth postoperative day, a small degree of conjunctival growth was observed over the amniotic membrane.
Fig. 1.
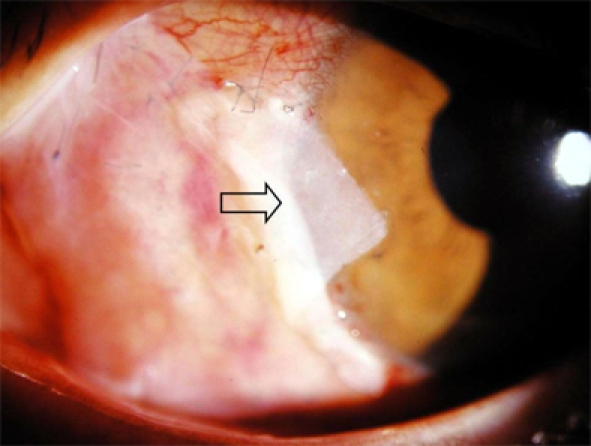
One day after pterygium excision, the amniotic membrane (arrow) was firmly attached to the sclera.
Routine histological sections of the amniotic membrane patches with H & E staining revealed one or two layers of uniformly small, cuboidal epithelial cells over a partially dissolved amniotic membrane (Fig. 2A). Significant inflammatory infiltration by cells, including polymorphonuclear leukocytes and some eosinophils and monocytes, was observed on the stromal side. On flat mounts, many round or spindle-shaped, infiltrating mononuclear cells, suggestive of primitive cells, were also observed (Fig. 2B).
Fig. 2.
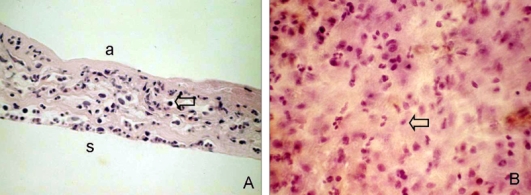
Histological features of a temporary amniotic membrane patch removed 4 days after pterygium excision. Amniotic membrane was sectioned and stained with H&E. (A) In a cross section, one or two layers of epithelium were observed over the partially dissolved amniotic membrane with marked mononuclear cell infiltration (arrow). (B) In a flat mount, numerous round or spindle-shaped mononuclear cells (arrow) had infiltrated and a small number of polymorphonuclear leukocytes were found on the stromal side. a, amnion side; s, amniotic membrane stroma. Original magnification: A 200×; B 400×.
Strong positive immunoreactivities to CD34, c-kit, AC133, and STRO-1 were detected in the stromal side of the amniotic membranes. CD34-positive cells were observed in some round or spindle-shaped mononuclear cells. Epithelial cells that had grown over the amniotic membrane did not show immunoreactivity to CD34 upon cross sectioning (Fig. 3A). Some CD34-positive, large, and round or spindle-shaped cells formed clusters resembling small vessels in some regions of the amniotic membrane (Fig. 3B, inset). In contrast to CD34, c-kit was differentially expressed within amniotic membrane patches. Of note, c-kit staining was found specifically associated with epithelium growing above the amniotic membrane (Fig. 3C). Immunoreactivity of c-kit was also observed in a heterogeneous cell population, comprised of spindle-shaped cells and small round mononuclear cells, resembling mast cells (Fig. 3D). STRO-1 was expressed by many stellate to spindle-shaped fibroblast-like cells (Fig. 3E). AC133 immunoreactivity was found in small, round cells or long, slender cells, which were scattered all amniotic membrane fields (Fig. 3F). No staining was found in normal amniotic membranes (micrographs not shown).
Fig. 3.
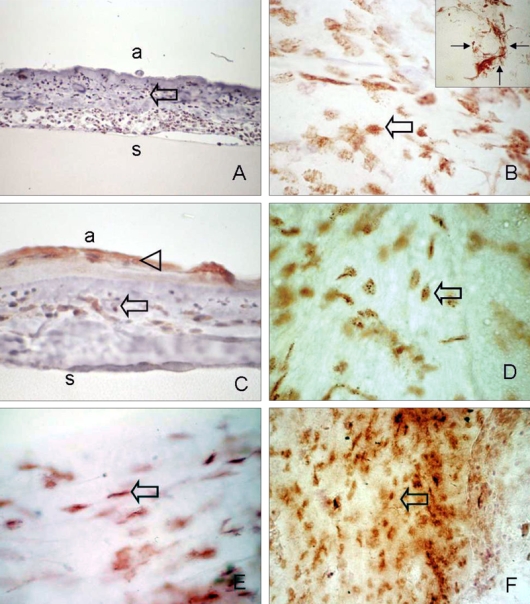
Immunohistochemical analysis of infiltrated stem cells in removed amniotic membrane. (A, B) Abundant CD34-positive cells, which were mainly composed of round or spindle-shaped mononuclear cells, were observed on the stromal side of the amniotic membrane (arrow). Occasionally, some CD34-positive cells formed clusters resembling small vessels (B; small arrow, inset). (C, D) c-kit immunoreactivity was found in grown epithelium (arrow head), spindle or round-shaped monocytes on the amniotic membrane (arrow). (E) SRTO-1-positive spindled-shaped cells resembling fibroblasts were observed (arrow). (F) Small round or long slender shaped AC133 positive cells (arrow). Sections were counterstained with hematoxylin. a, amnion side; s, amniotic membrane stroma. Original magnification: A, C 200×; B, B inset, D, E 400×. A, C: cross-section, B, D, E, F: flat mount.
The intensity of the CD34-, c-kit-, AC133-, and STRO-1-positive cells was variable among the amniotic membrane specimens. In cases which recurred, excessive infiltration by CD34-, c-kit-, AC133-, and STRO-1-positive cells was observed, but there was no obvious correlation between recurrence and the expression of any other antigen.
DISCUSSION
The reepithelialization and revascularization processes of denuded sclera after pterygium excision are not well-known. Some studies have implicated the centripetal migration of undifferentiated conjunctival cells and possible conjunctival epithelial stem cell involvement, but the exact roles of these cells and the mechanisms modulating the migration and differentiation of cells in the repair process are controversial.18 In addition, the aggressive recurrence of pterygium after surgical excision suggests the involvement of an unknown cellular mechanism in the pathogenesis and wound healing processes. In a previous study, we conclusively demonstrated the involvement of bone marrow-derived cells in the pathogenesis of pterygium.6
This study shows that CD34, c-kit, AC133, and STRO-1 are strongly expressed in amniotic membrane applied to bare sclera after pterygium removal. The leukocyte antigen, CD34, is known to be expressed on the surfaces of hematopoietic progenitor cells, vascular endothelial cells, and some fibroblasts.12 During the repair process after pterygium excision, round or spindle-shaped, CD34-positive mononuclear cells were observed on the stromal side of the amniotic membrane. Cells that stained positively for CD34 clustered in small, vessel-like shapes in some regions of amniotic membrane. Whether this vessel formation was due to the vasculogenic transformation of progenitor cells or to the angiogenesis of existing vessels is unclear. A recent study on staining patterns in vessels indicated that CD34 is expressed on endothelial cells in tumors and fetal tissue, but not during the angiogenic microvascular process in wounded tissue.19 This result suggests that CD34-positive endothelial cells involved in wound healing originate from bone marrow. Therefore, our findings provide evidence of bone marrow-derived progenitor cell involvement in the healing process.
AC133 is a marker for endothelial progenitor and neuronal stem cells and is rapidly downregulated during cell differentiation.20 Abundant AC133-positive cells were observed on removed amniotic membranes by morphological analysis, and were round to spindle-shaped, similar to CD34-positive cells. The specific roles of these cells are uncertain; however it is possible that these cells may differentiate into endothelial cells, since a previous study showed that a subset of CD34-positive cells express AC133 as a true functional marker of endothelial progenitor cells.21
In cross sections, we found that amniotic membranes were covered by one or two layers of small cuboidal epithelial cells that were morphologically similar to conjunctival basal epithelial cells. Additionally, aberrant c-kit expression was localized within the epithelium that had grown over the amniotic membrane. These findings are consistent with a report that corneal basal epithelial cells immunostain for SCF.5 These regenerated epithelial cells are an important source of a variety of cytokines, and chemotactic and growth factors, including SCF.22 Thus, our results provide more evidence that regenerated epithelial cells, indeed, contribute to the healing process. On a flat mount, c-kit-positive mononuclear cells appeared round or spindle-shaped and resembled mast cells or tissue macrophages. These findings are consistent with reports that mast cells are expressed in pterygial tissue and that their numbers significantly increase during the wound healing process.23,24 These cells may participate in the scarring caused by fibroblast proliferation or in angiogenesis by providing vascular endothelial growth factor.25
The results of our study suggest that bone marrow-derived progenitor cells are involved in healing and recurrence after pterygium excision. The basic mechanism of how bone marrow-derived progenitor cells migrate from the bone marrow into injured sites at a restricted vessel is unclear. A recent study indicated that tissue hypoxia might fundamentally govern progenitor cell recruitment and retention.26 Vascular disruption due to surgical excision is likely to cause a hypoxic zone on the ocular surface, in which the activation of trafficking-related factors, such as hypoxia inducible factor-1α (HIF-1α), facilitate progenitor cell recruitment in ischemic tissue requiring repair.
In addition to hypoxia, several other mechanisms may lead to progenitor cell release in response to tissue damage. Surgical injury might induce a rapid influx of not only inflammatory cells, but also of abundant growth factors and chemokines, such as IL-6 and matrix metalloprotease-9.27 These may stimulate bone marrow and enhance the migration of stem cells into the damaged ocular surface to repair damaged lesions via a rich vascular arcade of limbus. In addition, substance p, induced by surgical pain, may play a role in progenitor cell recruitment.28
Bone marrow-derived cells may be important for the renewal of resident stem cells, required for the maintenance of an organ or for the repair of tissue damage. Moreover, migrated bone marrow-derived cells would be activated by chemotactic factors released by certain inflammatory cells and keratocytes and gain the ability to differentiate into vascular endothelial cells, fibroblasts, and epithelial cells to recover the damaged surface.
Our findings also explain the features of recurrence after pterygium removal. After surgical excision, if abnormal microenvironments affected the wound healing process, then infiltrating progenitor cells would differentiate and proliferate more, thus contributing to alterations of fibroblasts and limbal stem cells. In addition, we observed many STRO-1-positive cells (a fibroblast characteristic) on the amniotic membrane. The inappropriate proliferation of bone marrow-derived circulating fibroblasts contributes to the genesis of subepithelial fibrosis and, thus, accelerates recurrence. This hypothesis is compatible with a finding of our previous study, which is that much stronger immunoreactivity to the progenitor cell markers is present in recurrent pterygium.6 Thus, stem cells may play a role as indicators of pterygium recurrence, and it is reasonable to believe that the more stem cells present, the higher the pterygium recurrence rate will be after removal.
The application of amniotic membrane for ocular surface reconstruction was found to be a useful therapeutic method with various advantages.29,30 It provides a healthy basement membrane for epithelial proliferation and contains various proteins that inhibit proteinases, which are destructive to tissue after the initial injury. The reported clinical outcomes after amniotic membrane graft for pterygia have demonstrated good results, although some studies reported high recurrence rate.30-33 The reasons for this variability are unclear, but many kinds of growth factors in amniotic membrane might induce excessive fibrovascular proliferation and pterygium recurrence. In a previous study, alkali-burned corneas, treated with amniotic membrane coverage for three days, showed enhanced wound healing, compared with those treated with coverage for seven days.34 Therefore, we reasoned that covering the bare sclera with the temporary amniotic membrane patch may alleviate possible adverse effects caused by permanent amniotic membrane grafting. Additionally, when amniotic membrane covers nerves exposed in the cornea, the application of amniotic membrane relieves pain. Based on these findings, we applied the amniotic membrane stromal-surface-down to the exposed sclera and observed many infiltrating cells, including bone marrow-derived progenitor cells and inflammatory cells, over the amniotic membrane. Thus, a temporary amniotic membrane patch could effectively prevent recurrence by absorbing excessively infiltrating stem cells and relieving pain by means of a biological wound-dressing effect.
To the best of our knowledge, our findings demonstrate, for the first time, the possibility that bone-marrow-derived cells are involved in the process of wound healing and in the recurrence of pterygium after surgical excision. Additional experiments are required to identify the factors that mediate recruitment, proliferation, and differentiation of progenitor cells to reveal the pathogenesis of pterygium and the wound healing process after pterygium removal. We hope that the clinical applications of this new concept will accelerate the healing process in a controlled fashion in the treatment of this disease.
Footnotes
This work was supported by the Strategic Research Program of Chung-Ang University, and in part by the Stem Cell Research Center of the 21C Frontier program funded by the Ministry of Science and Technology, Seoul, Korea, in 2003.
References
- 1.Coroneo MT, Di Girolamo N, Wakefield D. The pathogenesis of pterygia. Curr Opin Ophthalmol. 1999;10:282–288. doi: 10.1097/00055735-199908000-00011. [DOI] [PubMed] [Google Scholar]
- 2.Kwok LS, Coroneo MT. A model for pterygium formation. Cornea. 1994;13:219–224. doi: 10.1097/00003226-199405000-00005. [DOI] [PubMed] [Google Scholar]
- 3.Kennedy M, Kim KH, Harten B, Brown J, Planck S, Meshul C, et al. Ultraviolet irradiation induces the production of multiple cytokines by human corneal cells. Invest Ophthalmol Vis Sci. 1997;38:2483–2491. [PubMed] [Google Scholar]
- 4.Marcovich AL, Morad Y, Sandbank J, Huszar M, Rosner M, Pollack A, et al. Angiogenesis in pterygium: morphometric and immunohistochemical study. Curr Eye Res. 2002;25:17–22. doi: 10.1076/ceyr.25.1.17.9959. [DOI] [PubMed] [Google Scholar]
- 5.Nakagami T, Watanabe I, Murakami A, Okisaka S, Ebihara N. Expression of stem cell factor in pterygium. Jpn J Ophthalmol. 2000;44:193–197. doi: 10.1016/s0021-5155(99)00214-2. [DOI] [PubMed] [Google Scholar]
- 6.Ye J, Song YS, Kang SH, Yao K, Kim JC. Involvement of bone marrow-derived stem and progenitor cells in the pathogenesis of pterygium. Eye. 2004;18:839–843. doi: 10.1038/sj.eye.6701346. [DOI] [PubMed] [Google Scholar]
- 7.Orlic D, Kajstura J, Chimenti S, Jakoniuk I, Anderson SM, Li B, et al. Bone marrow cells regenerate infarcted myocardium. Nature. 2001;410:701–705. doi: 10.1038/35070587. [DOI] [PubMed] [Google Scholar]
- 8.Yamada M, Kubo H, Kobayashi S, Ishizawa K, Numasaki M, Ueda S, et al. Bone marrow-derived progenitor cells are important for lung repair after lipopolysaccharide-induced lung injury. J Immunol. 2004;172:1266–1272. doi: 10.4049/jimmunol.172.2.1266. [DOI] [PubMed] [Google Scholar]
- 9.Asahara T, Masuda H, Takahashi T, Kalka C, Pastore C, Silver M, et al. Bone marrow origin of endothelial progenitor cells responsible for postnatal vasculogenesis in physiological and pathological neovascularization. Circ Res. 1999;85:221–228. doi: 10.1161/01.res.85.3.221. [DOI] [PubMed] [Google Scholar]
- 10.Makino S, Fukuda K, Miyoshi S, Konishi F, Kodama H, Pan J, et al. Cardiomyocytes can be generated from marrow stromal cells in vitro. J Clin Invest. 1999;103:697–705. doi: 10.1172/JCI5298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Boyd AW. Human leukocyte antigens: an update on structure, function, and nomenclature. Pathology. 1987;19:329–337. doi: 10.3109/00313028709103879. [DOI] [PubMed] [Google Scholar]
- 12.Krause DS, Ito T, Fackler MJ, Smith OM, Collector MI, Sharkis SJ, et al. Characterization of murine CD34, a marker for hematopoietic progenitor and stem cells. Blood. 1994;84:691–701. [PubMed] [Google Scholar]
- 13.Zsebo KM, Williams DA, Geissler EN, Broudy VC, Martin FH, Atkins HL, et al. Stem cell factor is encoded at the S1 locus of the mouse and is the ligand for the c-kit tyrosine kinase receptor. Cell. 1990;63:213–224. doi: 10.1016/0092-8674(90)90302-u. [DOI] [PubMed] [Google Scholar]
- 14.Schmeisser A, Strasser RH. Phenotypic overlap between hematopoietic cells with suggested angioblastic potential vascular endothelial cells. J Hematother Stem Cell Res. 2002;11:69–79. doi: 10.1089/152581602753448540. [DOI] [PubMed] [Google Scholar]
- 15.Majka M, Ratajczak J, Machalinski B, Carter A, Pizzini D, Wasik MA, et al. Expression, regulation and function of AC133, a putative cell surface marker of primitive human hematopoietic cells. Folia Histochem Cytobiol. 2000;38:53–63. [PubMed] [Google Scholar]
- 16.Simmons PJ, Torok-Strob B. Identification of stromal cell precursors in human bone marrow by a novel monoclonal antibody, STRO-1. Blood. 1991;78:55–62. [PubMed] [Google Scholar]
- 17.Di Girolamo N, McCluskey P, Lloyd A, Coroneo MT, Wakefield D. Expression of MMPs and TIMPs in human pterygia and cultured pterygium epithelial cells. Invest Ophthalmol Vis Sci. 2000;41:671–679. [PubMed] [Google Scholar]
- 18.Wei ZG, Wu RL, Lavker RM, Sun TT. In vitro growth and differentiation of rabbit bulbar, fornix, and palpebral conjunctival epithelia. Implications on conjunctival epithelial transdifferentiation and stem cells. Invest Ophthalmol Vis Sci. 1993;34:1814–1828. [PubMed] [Google Scholar]
- 19.Schlingemann RO, Rietveld FJ, de Waal RM, Bradley NJ, Skene AI, Davies AJ, et al. Leukocyte antigen CD34 is expressed by a subset of cultured endothelial cells and on endothelial abluminal microprocesses in the tumor stroma. Lab Invest. 1990;62:690–696. [PubMed] [Google Scholar]
- 20.Gehling UM, Ergun S, Schumacher U, Wagener C, Pantel K, Otte M, et al. In vitro differentiation of endothelial cells from AC133-positive progenitor cells. Blood. 2000;95:3106–3112. [PubMed] [Google Scholar]
- 21.Peichev M, Naiyer AJ, Pereira D, Zhu Z, Lane WJ, Williams M, et al. Expression of VEGFR-2 and AC133 by circulating human CD34+ cells identifies a population of functional endothelial precursors. Blood. 2000;95:952–958. [PubMed] [Google Scholar]
- 22.Pompen M, Smids BS, Dingemans KP, Jansen HM, Out TA, Lutter R. Lung epithelial H292 cells induce differentiation of immature human HMC-1 mast cells by interleukin-6 and stem cell factor. Clin Exp Allergy. 2000;30:1104–1112. doi: 10.1046/j.1365-2222.2000.00881.x. [DOI] [PubMed] [Google Scholar]
- 23.Sakaguchi H, Takai S, Sakaguchi M, Sugiyama T, Ishihara T, Yao Y, et al. Chymase and angiotensin converting enzyme activities in a hamster model of glaucoma filtering surgery. Curr Eye Res. 2002;24:325–331. doi: 10.1076/ceyr.24.5.325.8519. [DOI] [PubMed] [Google Scholar]
- 24.Nakagami T, Murakami A, Okisaka S, Ebihara N. Mast cells in pterygium: number and phenotype. Jpn J Ophthalmol. 1999;43:75–79. doi: 10.1016/s0021-5155(98)00069-0. [DOI] [PubMed] [Google Scholar]
- 25.Heissig B, Werb Z, Rafii S, Hattori K. Role of c-kit/Kit ligand signaling in regulating vasculogenesis. Thromb Haemost. 2003;90:570–576. doi: 10.1160/TH03-03-0188. [DOI] [PubMed] [Google Scholar]
- 26.Ceradini DJ, Kulkarni AR, Callaghan MJ, Tepper OM, Bastidas N, Kleinman ME, et al. Progenitor cell trafficking is regulated by hypoxic gradients through HIF-1 induction of SDF-1. Nat Med. 2004;10:858–864. doi: 10.1038/nm1075. [DOI] [PubMed] [Google Scholar]
- 27.Janowska-Wieczorek A, Marquez LA, Nabholtz JM, Cabuhat ML, Montano J, Chang H, et al. Growth factors and cytokines upregulate gelatinase expression in bone marrow CD34(+) cells and their transmigration through reconstituted basement membrane. Blood. 1999;93:3379–3390. [PubMed] [Google Scholar]
- 28.Pelletier L, Angonin R, Regnard J, Fellmann D, Charbord P. Human bone marrow angiogenesis: in vitro modulation by substance P and neurokinin A. Br J Haematol. 2002;119:1083–1089. doi: 10.1046/j.1365-2141.2002.03969.x. [DOI] [PubMed] [Google Scholar]
- 29.Park WC, Tseng SC. Modulation of acute inflammation and keratocyte death by suturing, blood, and amniotic membrane in PRK. Invest Ophthalmol Vis Sci. 2000;41:2906–2914. [PubMed] [Google Scholar]
- 30.Jang JH, Choi TH. The effect of amniotic membrane transplantation for pterygium excision. J Korean Ophthalmol Soc. 2005;46:597–604. [Google Scholar]
- 31.Ma DH, See LC, Hwang YS, Wang SF. Comparison of amniotic membrane graft alone or combined with intraoperative mitomycin C to prevent recurrence after excision of recurrent pterygia. Cornea. 2005;24:141–150. doi: 10.1097/01.ico.0000141237.71837.d8. [DOI] [PubMed] [Google Scholar]
- 32.Essex RW, Snibson GR, Daniell M, Tole DM. Amniotic membrane grafting in the surgical management of primary pterygium. Clin Experiment Ophthalmol. 2004;32:501–504. doi: 10.1111/j.1442-9071.2004.00884.x. [DOI] [PubMed] [Google Scholar]
- 33.Tananuvat N, Martin T. The results of amniotic membrane transplantation for primary pterygium compared with conjunctival autograft. Cornea. 2004;23:458–463. doi: 10.1097/01.ico.0000116522.57227.97. [DOI] [PubMed] [Google Scholar]
- 34.Kim JS, Kim JC, Na BK, Jeong JM, Song CY. Amniotic membrane patching promotes healing and inhibits proteinase activity on wound healing following acute corneal alkali burn. Exp Eye Res. 2000;70:329–337. doi: 10.1006/exer.1999.0794. [DOI] [PubMed] [Google Scholar]