

Abstract
Random disturbances of signals, termed ‘noise’, pose a fundamental problem for information processing and affect all aspects of nervous-system function. However, the nature, amount and impact of noise in the nervous system have only recently been addressed in a quantitative manner. Experimental and computational methods have shown that multiple noise sources contribute to cellular and behavioural trial-to-trial variability. We review the sources of noise in the nervous system, from the molecular to the behavioural level, and show how noise contributes to trial-to-trial variability. We highlight how noise affects neuronal networks and the principles the nervous system applies to counter detrimental effects of noise, and briefly discuss noise's potential benefits.
Variability is a prominent feature of behaviour. Variability in perception and action is observed even when external conditions, such as the sensory input or task goal, are kept as constant as possible. Such variability is also observed at the neuronal level1-4. What are the sources of this variability? Here, a linguistic problem arises, as each field has developed its own interpretation of terms such as variability, fluctuation and noise. In this Review, we use the term variability to refer to changes in some measurable quantity, such as spike timing or movement duration. Importantly, the term variability does not indicate that a particular mechanism has generated the variability, and does not suggest whether the variability is beneficial or detrimental. Trial-to-trial variability can arise from two distinct sources. The first source is the deterministic properties of the system. For example, the initial state of the neural circuitry will vary at the start of each trial, leading to different neuronal and behavioural responses. The variability in the response will be exacerbated if the system's dynamics are highly sensitive to the initial conditions. The second source of variability is noise, which is defined in the Oxford English Dictionary as “random or irregular fluctuations or disturbances which are not part of a signal […] or which interfere with or obscure a signal or more generally any distortions or additions which interfere with the transfer of information”.
Whereas previous reviews have focused on neuronal variability in general, we focus here on work directly relating to noise. Noise permeates every level of the nervous system, from the perception of sensory signals to the generation of motor responses, and poses a fundamental problem for information processing5,6. In recent years the extent to which noise is present and how noise shapes the structure and function of nervous systems have been studied. In this Review, we begin by considering the nature, amount and effects of noise in the CNS. As the brain's purpose is to receive and process information and act in response to that information [FIG. 1], we then examine how noise affects motor behaviour, considering the contribution of noise to variability at each level of the behavioural loop. Finally, we discuss the strategies that the nervous system uses to counter, compensate for or account for noise in perception, decision making and motor behaviour. Given the many levels and systems that are spanned, we cannot provide a comprehensive Review, but instead we pick out specific examples that reflect in a more general manner the constraints and limitations that noise sets in the CNS; the benefits of noise are discussed in BOX 1.
Figure 1. Overvie of the behavioural loop and the stages at which noise is present in the nervous system.

a | Sources of sensory noise include the transduction of signals. This is exemplified here by a photoreceptor and its signal-amplification cascade. b | Sources of cellular noise include the ion channels of excitable membranes, synaptic transmission and network interactions (see BOX 2). c | Sources of motor noise include motor neurons and muscle. In the behavioural task shown (catching a ball), the nervous system has to act in the presence of noise in sensing, information processing and movement.
Box 1. Benefits of noise.
Noise is not only a problem for neurons: it can also be a solution to other information-processing issues. Several strategies have been adopted to use noise in this fashion. For example, stochastic resonance is a process by which the ability of threshold-like systems to detect and transmit weak (periodic) signals can be enhanced by the presence of a certain level of noise87,175. At low noise levels, the sensory signal does not cause the system to cross the threshold and few signals are detected. For large noise levels, the response is dominated by the noise. For intermediate noise intensities, however, the noise allows the signal to reach the threshold but does not swamp it. For stochastic resonance to be useful, positive detection of a sub-threshold input must be more desirable than a failure to detect a supra-threshold input. Since its first discovery in cat visual neurons176, stochastic-resonance-type effects have been demonstrated in a range of sensory systems. These include crayfish mechanoreceptors177, shark multimodal sensory cells178, cricket cercal sensory neurons179 and human muscle spindles180. The behavioural impact of stochastic resonance has been directly demonstrated and manipulated in passive electrosensing paddlefish181 and in human balance control182.
In addition, in spike-generating neurons, sub-threshold signals have no effect on the output of the system. Noise can transform such threshold nonlinearities by making sub-threshold inputs more likely to cross the threshold, and this becomes more likely the closer the inputs are to the threshold. Thus, when it is averaged over time, this noise produces an effectively smoothed nonlinea56. This facilitates spike initiation and can improve neural-network behaviour, as was shown in studies of contrast invariance of orientation tuning in the primary visual cortex183. Moreover, neuronal networks that have formed in the presence of noise will be more robust and explore more states, which will facilitate learning and adaptation to the changing demands of a dynamic environment184,185.
Sensory noise
External sensory stimuli are intrinsically noisy because they are either thermodynamic or quantum mechanical in nature. For example, all forms of chemical sensing (including smell and gustation) are affected by thermodynamic noise because molecules arrive at the receptor at random rates owing to diffusion and because receptor proteins are limited in their ability to accurately count the number of signalling molecules7,8. Similarly, vision involves the absorption of photons that arrive at the photoreceptor at a rate governed by a Poisson process. This places a physical limit on contrast sensitivity in vision, which is reduced at low light levels — when fewer photons arrive at the photoreceptor9.
At the first stage of perception (FIG.1a), energy in the sensory stimulus is converted into a chemical signal (through photon absorption or ligand-binding of odour molecules) or a mechanical signal (such as the movement of hair cells in hearing). The subsequent transduction process amplifies the sensory signal and converts it into an electrical one, either directly or indirectly through second-messenger cascades. Any sensory noise that is already present or that is generated during the amplification process (transducer noise10) wil will increase trial-to-trial variability. Therefore, noise levels set perceptual threshold for later stages of information processing, as signals that are weaker than the noise cannot be distinguished from it after amplification11. This is rigorously underpinned by the data-processing inequality theorem12, which states that subsequent stages of processing (even if they are noise free) cannot extract more information than is present at earlier stages. Therefore, to reduce noise, organisms often pay a high metabolic and structural price at the first stage of processing (the sensory stage). For example, a fly's photoreceptors account for 10% of its resting metabolic consumption and its eye's optics make up over 20% of the flight payload13.
Cellular noise
If neurons are driven with identical time-varying stimuli over repeated trials, the timing of the resultant action potentials (APs) varies across the trials3,14-19. This variability is on the order of milliseconds or lower14,15,20-25, but because cortical neurons can detect the coincident arrival of APs on millisecond timescales26,27, the variability might well be physiologically relevant. Indeed, the precision of single-neuron AP timing on the milli- and sub-millisecond scale has been shown to be behaviourally relevant in perceptionn28,29 and movement30. To what extent this neuronal variability contributes to meaningful processing (as opposed to being meaningless noise) is the fundamental question of neural coding4,19,31-33. A key issue is that neuronal activity might look random without actually being random.
Neuronal variability (both in and across trials) can exhibit statistical characteristics (such as the mean and variance) that match those of random processes. However, even when neuronal-firing statistics match those of a random process, it does not necessarily follow that the firing is generated by random processes. In fact, we know from Shannon's theory of information5,12 that when optimal information encoding is used to maximize information transmission, neural signals might look random 31. Furthermore, neuronal variability is not equal in all neurons. The Fano factor is a simple measure of variability that ignores temporal structure and higher-order statistics. Neural responses without variability have Fano factors of zero, whereas Poisson processes (which are highly variable) have a Fano factor of one. Some cortical neurons are highly variable, with Fano factors of one or greater2,3,34,35, whereas others have Fano factors that are closer to zero24,36,37. Similarly, there is a range of variability in neurons in the mammalian24,38 and invertebrate18,39 visual pathways. Moreover, high- and low-variability neurons are often observed in the same region, and a single neuron can respond with different amounts of variability depending on the stimulus conditions18,39.
Multiple factors contribute to neuronal trial-to-trial variability. These include changes in the internal states of neurons and networks, and random processes inside neurons and neuronal networks40,41. To what extent each of these factors contributes to the total observed trial-to-trial variability remains unclear, especially as network [BOX 2] and other effects might reduce variability despite the presence of noise. In general, the impact of noise on cellular function will inescapably increase neuronal variability (but see BOX 1), and thus we can compare the amount of variability that is produced by noise with the total observed variability to give us an idea of the relative contribution of noise to trial-to-trial variability.
Box 2. Noise build-up in networks.
How can neural networks maintain stable activity in the presence of noise6? There are several ways in which networks can affect overall noise levels. The figure illustrates this with three simple examples in which graded-potential neurons linearly sum inputs. Part a shows convergence of signals onto a single neuron. If the incoming signals have independent noise, then noise levels in the postsynaptic neuron will scale in proportion to the square root of the number of signals (N), whereas the signal scales in proportion to N. If the noise in the signals is perfectly correlated, then the noise in the neuron will also scale in proportion to N. Part b shows the passage of signals through a series of neurons. In this case, noise levels increase in proportion to the square root of the number of successive neurons. By contrast, parallel connections (not shown) do not augment noise through network interactions. Part c shows that recurrence in networks results in the build-up of correlated noise. 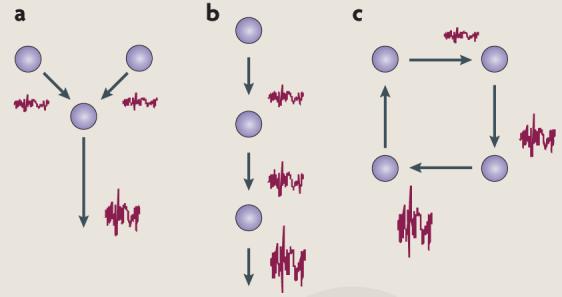
Other computational operations in each neuron can alter the build-up of network noise. The linear operation of amplification leaves the signal-to-noise ratio unchanged. Nonlinear operations, such as multiplication and thresholding, affect noise build-up differently. In general, multiplication operations increase the coefficient of variation (CV) of the output, whereas thresholding decreases the CV. Several studies have examined how noise acts in neuron-like nonlinear systems186,187. The highly parallel and distributed yet compact structure of the CNS might help to limit the amount of noise that builds up.
Experimental evidence suggests that average neuronal activity levels are maintained by homeostatic plasticity mechanisms that dynamically set synaptic strengths188, ion-channel expression157 or the release of neuromodulators25. This in turn suggests that networks of neurons can dynamically adjust to attenuate noise effects. Moreover, these networks might be wired so that large variations in the response properties of individual neurons have little effect on network behaviour189.
Furthermore, in many spiking neurons190,191, doubling the input results in less then twice the output. This suggests that presynaptic noise and intracellular noise are attenuated as the signal passes through the neuron. The fact that the noise remains so small suggests that neuronal networks can be organized in a way that prevents local noise accumulating as neural signals propagate through them41. Thus, the analogue (membrane-potential based) nature of local neural computation (computation within neurons) and the more digital (action-potential based) nature of global information transmission192,193 might be essential ingredients in building noise-robust computational circuits194. Figure modified, with permission, from REF. 195 © (2001) Wiley.
What are the sources of noise in neurons? In each neuron, noise accumulates owing to randomness in the cellular machinery that processes information42 [FIG. 1b] and can further increase as a result of nonlinear computations and network interactions [BOX 2]. At the biochemical and biophysical level there are many stochastic processes at work in neurons. These include protein production and degradation, the opening and closing of ion channels, the fusing of synaptic vesicles and the diffusion and binding of signalling molecules to receptors. It is often implicitly assumed that averaging large numbers of such stochastic elements effectively eliminates the randomness of individual elements. However, this assumption requires reassessment. Neurons perform highly nonlinear operations that involve high gain amplification and positive feedback. Therefore, small biochemical and electrochemical fluctuations (when considering systems at the molecular level we use the term fluctuation interchangeably with noise) can significantly alter whole-cell responses. For example, when the membrane potential is near the firing threshold, the generation of an AP becomes highly sensitive to noise43,44. Large neuronal structures, such as the squid giant axon (which can measure up to 1 mm in diameter), have been used extensively to investigate neural mechanisms42,45-48. Given the scale of these structures, they appear to function deterministically, because large numbers of signalling molecules are involved and random fluctuations are indeed averaged out. However, many neurons are tiny: cerebellar parallel fibres have an average diameter of 0.2 μm; C-fibres, which are involved in sensory and pain transmission, range between 0.1 and 0.2 μm in diameter; and the unmyelinated pyramidal-cell axon collaterals, which form the vast majority of local cortico–cortical connections, have an average diameter of 0.3 μm. Similarly, most (spiny- or bouton-type) CNS synapses have sub-micrometer dimensions. At these small length scales the numbers of molecules involved are small and the influence of noise is dramatically increased. Here we review the main sources of noise in the nervous system at the cellular level and the consequences for neuronal function.
Electrical noise and action potentials
The membrane potential is used both for local computation and to carry APs. Although variability in resting membrane potential49,50 (membrane-potential fluctuations) and AP threshold51 have been studied for a long time, the mechanisms that underlie these fluctuations have only recently gained attention. Electrical noise in neurons causes membrane-potential fluctuations even in the absence of synaptic inputs. The most dominant source of such electrical noise is channel noise52-54 (FIG. 1b) — electrical currents produced by the random opening and closing of voltage- or ligand-gated ion channels. Stochastic models have shown that channel noise can account for variability in the AP threshold at nodes of Ranvier55 and the reliability of AP initiation in membrane patches43,56,57. Furthermore, patch-clamp experiments in vitro show that channel noise in the dendrites and in the soma produces membrane-potential fluctuations that are large enough to affect AP timing58-61. Both the initiation and the propagation of APs can be affected by channel noise.
At the site of AP initiation — the soma or the axon hillock — channel noise can affect the timing of APs (despite the comparatively large number of ion channels that are present at these sites)44,55. Stochastic simulations have shown that it is not the number of ion channels that are open at the peak of the AP that determines its timing precision, but the much smaller number of ion channels that are open at the AP threshold. The resulting variability in spike timing is larger for weaker driving signals, for which the likelihood of the membrane potential reaching the AP threshold is more affected by channel noise54,62. The effects of channel noise also increase dramatically as neurons become smaller63, because the opening of an ion channel affects the membrane potential in proportion to the membrane's input resistance, which increases rapidly with decreasing diameter64. In axons of less than 0.3 μm diameter, the input resistance is large enough that spontaneous opening of single Na+ channels at the resting potential can produce ‘Na+ sparks’ that can trigger APs in the absence of any other inputs. These ‘rogue’ APs become exponentially more frequent as axon diameter decreases, rendering axons below 0.08–0.10 μm diameter useless for communication. This lower limit matches the smallest diameters of axons across species. Analogously, noise sets the lower limit for the diameter of excitable cell bodies to ~3 μm. Thus, thermodynamic noise in individual ion-channel proteins sets an upper limit to the wiring densities of the whole brain63.
Channel noise also affects AP propagation in axons, producing trial-to-trial variability in AP timing. This variability occurs whenever the input resistance of an axon is large enough that small numbers of ion channels can support AP conduction63. Using biophysical theory and stochastic simulations, it was shown that in CNS axons of 0.1–0.5 μm diameter, channel noise introduces significant jitter in AP propagation65 (FIG. 2a). Thus, the variability in postsynaptic responses that results from axonal channel noise will increase the longer and thinner the presynaptic axon. Moreover, populations of ion channels can retain a memory of axonal activity for several hundred milliseconds, owing to a complex interaction between the internal states of ion-channel populations and the membrane potential. This history dependency results in some patterns of spikes (such as bursts) being less affected by noise than others65. Such ‘message-dependent’ noise has been observed in mammalian neurons66,67; however, this effect is missed when models use stochastic approximations (for example, the Langevin or Fokker–Planck models) or ignore spatial interactions. Despite the evidence for the importance of variability in AP propagation for trial-to-trial variability, this has tended to be overlooked by most experimental studies (except those of AP conduction failure47), with postsynaptic variability being mainly attributed to synapses.
Figure 2. Examples amples of cellular noise.

a | Channel noise as a source of trial-to-trial variability in action potential (AP) propagation. Stochastic simulations of the response of a 0.2 μm diameter CNS axon (comparable with a cerebellar parallel fibre) in response to repeated identical current stimuli and initial conditions are shown. The only source of variability is the stochastic opening and closing of a million individually simulated ion channels. Spike trains were triggered by a time-varying current stimulus (top plot). Spike raster plots for each measurement site are shown, from the soma (second-from-top plot) down to the most distal part (the axon; bottom plot). In each raster plot, the precise timing of spikes is marked by dots, which are stacked over each other for each repeated trial (there were 60 trials). The shift of the overall spike pattern across rows reflects the average propagation speed of the APs. The raster plot of the somatic measurement reflects spike-time variability from AP initiation. Owing to channel noise, the spike-time variability rapidl increases the further the AP propagates, and it eventually reaches millisecond orders. b | In vitro paired patch-clamp recordings demonstrate the trial-to-trial variability of synaptic transmission in rat somatosensory cortex slices. Six consecutive postsynaptic responses (black traces) to an identical presynaptic-stimulation pattern (top trace) are shown, along with the ensemble mean response (grey trace) from over 50 trials. Part a modified from REF. 65. Part b modified, with permission, from REEF. 77 © (2006) American Physical Society.
Why is AP propagation so sensitive to noise, contrary to previous claims42,44,47,68,69? Detailed stochastic modelling has shown65 that the leading edge of AP propagation is driven by a relatively small — and thus noisy — ionic current flowing inside the axon. This causes jitter in the speed of the propagation of the AP and thus results in variability in AP timing. By contrast, the current following the leading edge is large and therefore conduction failures owing to channel noise are unlikely, even in very thin axons (where <3% of all APs fail). Thus, axonal channel noise cannot account for the failure rates that have been reported in much larger CNS axons (where 5–80% of APs fail70), and conduction failures that have been observed in the nervous system are more likely to be due to computational mechanisms that allow ‘editing’ of spike trains71 than to noise65.
Other electrical-noise sources include Johnson noise and shot noise, which are three orders of magnitude smaller than channel noise in CNS neurons72,73. Moreover, variations in the activity of nearby neurons could produce ‘cross-talk noise’ in the confined spaces of the CNS. Such cross-talk can arise through ephaptic coupling70, large changes of extracellular ion concentration after electrical signalling74, and spillover of neurotransmitters75 between unrelated synapses.
Synaptic noise
If a presynaptic cell is driven repeatedly with identical stimuli, there is trial-to-trial variability in the postsynaptic response76,77 (FIG. 2b). This variability could arise from noise42,47 or from a deterministic process that is too complex to grasp and thus appears random77,78. Here we discuss evidence for the considerable contribution of noise to synaptic variability.
Many neocortical cells receive an intense synaptic bombardment from thousands of synapses79-81, which is often referred to as ‘synaptic background noise’ (REFS 82,83). However, the rich set of dendritic mechanisms that allow individual synapses to interact suggests that this ‘background’ activity is unlikely to be composed only of noise26,27,84,85. Indeed, experimental evidence and computational arguments suggest that the synaptic background activity contains meaningful structure16,85-87. Nevertheless, there are microscopic sources of true noise present at each synapse that are also likely to contribute to this synaptic background variability and influence neuronal firing42,47.
The classic manifestation of synaptic noise is the spontaneous miniature postsynaptic current (mPSC) that can be recorded in the absence of presynaptic input. Katz and collaborators interpreted mPSCs as being the result of spontaneously released neurotransmitter vesicles, thus establishing the quantal nature of synaptic transmission46. This work remains an exquisite example of how taking noise into account informs our understanding of neural mechanisms.
Several sources of noise at synapses can influence information transmission and induce variability (FIG 1b). mPSCs are caused by random events in the synaptic-transmission machinery, such as the spontaneous opening of intracellular Ca2+ stores88,89, synaptic Ca2+-channel noise, spontaneous triggering of the vesicle-release pathway48 or spontaneous fusion of a vesicle with the membrane. Once vesicles are released they induce a postsynaptic current, the amplitude of which shows considerable trial-to-trial variability (the coefficient of variation on (CV) being typically >0.2 (REFS 90,91)). To what extent can this variability in the postsynaptic response be attributed to noise? First, the same stochastic processes that produce spontaneous mPSCs are also present during normal synaptic transmission, and will alter the amplitude of the postsynaptic current. Second, the width (duration of channel opening) of the presynaptic AP determines the size of the Ca2+ signal that drives vesicle release and governs the number of vesicles that are released as well as the as the probability of release and AP width variability can result from axonal channel noise, which becomes significant for CNS synapses that are innervated by neurons with thin axons65.
Several additional factors have been shown to affect postsynaptic-response amplitude, each of which relies on noisy biochemical mechanisms and involves small numbers of molecules and is therefore subject to considerable thermodynamic noise. First, variability in the number of neurotransmitter molecules released per vesicle (~2000) arises owing to variations in vesicle size92 and vesicular neurotransmitter concentration93. Second, there is variability owing to the randomness of the diffusion of a relatively small number of molecules (CV = 0.16 (REF. 94)). Third, the location of vesicle release in the synaptic cleft has an impact on the postsynaptic response (CV = 0.37 (REF. 94)). Vesicles are distributed over the synaptic active zone and, as each AP will trigger the release of only one of them, the location varies from event to event. Fourth, synaptic-receptor channel noise increases the variability, especially if only a small number of receptors are involved95. Fifth, the number96 and density97 of receptor proteins at any synapse might stochastically vary over time, as the expression and degradation of proteins is limited by thermodynamic noise98.
In addition to variability in response amplitude, some CNS synapses release either one or no vesicles in response to an AP. The vesicle-release probability at small and bouton-type central synapses is typically low and is controlled by plasticity and adaptation mechanisms91. The probability of release itself could constitute a signal for information processing99. Therefore, the accuracy with which vesicle-release probability can be controlled might be computationally important; however, this has not been adequately quantified.
Summing up the effect on postsynaptic variability from the above synaptic noise sources, we note that the total observed synaptic trial-to-trial variability in many synapses (CV >0.2) can be fully accounted for by noise. However, there might also be biochemical mechanisms that reduce noise100.
Motor noise
We interact with the environment through movements, which are inherently variable from trial-to-trial. To generate movement the signals from the CNS have to be converted into mechanical forces in the muscle fibres (FIG 1c). The force that a single motor neuron can command is directly proportional to the number of muscle fibres that it innervates. When small forces are generated, motor neurons that innervate a small number of muscle fibres are active. When larger forces are generated, additional motor neurons that innervate a larger number of muscle fibres are also active. This is known as Henneman's size principle101. Moreover, as whole-muscle force increases, the firing rates of the active motor neurons increase, such that those that innervate a small number of muscle fibres have the highest firing rate.
The variability in the force that is produced by a whole human skeletal muscle is proportional to the average force that is produced by that muscle102,103. This has been attributed to the physiological organization of the pool of motor neurons and their muscle fibres103-106: each AP arriving at the muscle fibre induces a ‘twitch’. At low firing rates these twitches are separated in time, but as firing rates increase the twitches fuse into one smooth contraction. Whole-muscle force is determined by the number of active motor neurons and the firing rates of these neurons. The motor neuron that innervates the most fibres will have the lowest firing rate and will therefore induce unfused twitches in the muscle fibres that it innervates. Thus, any variability in the force that is generated by the muscle fibres that are innervated by this motor neuron will contribute most to whole-muscle force variability.
Three mechanisms contribute to the variability in the force that is generated by muscle fibres. First, even if a motor neuron fires perfectly periodically, there will be ‘ripples’ in the force that is generated by its muscle fibres, owing to unfused twitches. This effect is further enhanced by the synchronization of motor neurons through common mechanosensory feedback107. Second, motor neurons are subject to the same sources of cellular noise as any other neuron, making noise appreciable in AP timing in myelinated motor axons of 10 μm diameter108 and at the neuromuscular junction109. The resulting AP timing variability will reduce the periodicity of the force and thus increase its variability. Both of these factors will contribute to overall muscle-force variability110. Third, each twitch triggered by a single AP might also show trial-to-trial variability in its amplitude and duration, owing to noise in the biochemical cascade that generates the twitch force. However, to our knowledge this has not been quantified. In addition, as in thin axons, Ca2+-channel noise in muscle fibres111 or stochastic processes in energy release and transport could also produce random twitches. Furthermore, noise might result from unrelated electrical cross-talk between motor neurons112,113 or muscle fibres114, which could recruit other muscle fibres by ephaptic coupling.
Our present knowledge of force variability is based on isometric contractions (in which muscle length does not change), and it is unclear how this translates to variability during movement. The effect of single motor-neuron spikes on muscle movement has been measured only in invertebrate systems, in which it was shown that variability in spike timing (on the order of milliseconds) and in the number of spikes (±1) produces variability in muscle length of up to 10%30,115-117. These invertebrate muscles are comparable in scale to the human laryngeal muscles that control speech production, which have to operate with millisecond precision. However, little is known about the characteristics, activation and reliability of such muscles118.
Human motor behaviour — from eye movements119-121 to hand trajectories119,122,123 — can be explained by optimal control models that generate movements in a way that minimizes the impact of motor noise. It remains unclear how much of the observed trial-to-trial movement variability is due to motor-neuron and muscle noise and how much is due to other sources of variability in the (spinal) motor commands121,123.
Principles of how the CNS manages noise
In general, noise cannot be removed from a signal once it has been added. Furthermore, it is important to note that in some cases it is not always desirable to remove noise, as noise can have beneficial consequences for information processing (BOX 1). However, there are several principles that can be used to minimize the negative consequences of noise. We now review two key principles — averaging and prior knowledge — that the CNS applies at multiple levels.
The principle of averaging can be applied whenever redundant information is present in the sensory inputs to the CNS or is generated by the CNS. Averaging can counter noise if several units (such as receptor molecules, neurons or muscles) carry the same signal and each unit is affected by independent sources of noise (FIG. 2b). Averaging is seen at the very first stage of sensory processing. For example, the stereocilia of auditory hair cells capture sound vibrations and open mechanically gated ion channels. These stereocilia are mechanically coupled and so they move together, averaging random fluctuations in the movement of individual stereocilia124. Similarly, visual inputs are typically averaged over photoreceptors with adjacent or overlapping receptive fields69. Moreover, hair cells and photoreceptors are graded-potential neurons that do not use APs but instead communicate their varying membrane potential through graded synapses. This makes noise removal through averaging a straightforward operation for their postsynaptic membranes125.
Counterintuitively, divergence (one neuron synapsing onto many) can also support averaging. When signals are sent over long distances through noisy axons, rather than using a single axon it can be beneficial to send the same signal redundantly over multiple axons and then combine these signals at the destination. Crucially, for such a mechanism to reduce noise the initial divergence of one signal into many must be highly reliable. Such divergence is seen in auditory inner hair cells, which provide a divergent input to 10–30 ganglion cells through a specialized ‘ribbon synapse’ (REF. 126).
Prior knowledge can also be used to counter noise. If the structure of the signal and/or noise is known it can be used to distinguish the signal from the noise. This principle is especially helpful in dealing with sensory signals that, in the natural world, are highly structured redundant127-129. By using prior knowledge about the expected structure, sensory processing can compensate for noise. This is manifest in the notion that a neuron's receptive field tells us what message the neuron is conveying130. Signal-detection theory shows that the optimal signal detector, subject to additive noise, is obtained by matching all parameters of the detector to those of the signal to be detected131: in neuroscience this is termed the matched-filter principle132. Thus, the structures of receptive fields embody prior knowledge about the expected inputs and thereby allow neurons to attenuate the impact of noise.
Simple averaging works best when each signal source is corrupted by a similar amount of noise. Therefore, principles of averaging and prior knowledge are often combined in the nervous system when the sources are affected by different amounts of noise. Prior knowledge about the amount of noise for a given source allows for weighted averaging. In general, the less noisy (more reliable) inputs should contribute more to the averaging process than more noisy (less reliable) inputs. This has been demonstrated in several behavioural studies in which subjects were required to integrate inputs from different pairs of sensory sources133-140. The studies showed that the weight given to each source was proportional to its reliability (the inverse of the variance of the source), demonstrating that the nervous system has prior knowledge about the variability of its senses. Moreover, the observed behavioural variability in estimation tasks involving multiple sensory sources could be predicted from the behavioural variability to the individual sources if the source variability was mathematically treated as independent noise133-137. This strongly suggests that most of the behavioural variability in such sensory tasks arises from noise rather than from deterministic sources of variability. The motor system also applies a weighted averaging mechanism of this type to reduce the consequences of noise. For example, when redundant muscles can rotate a joint, the muscles are co-activated in a way that minimizes total movement variability141.
Averaging is used in many neural systems in which information is encoded as patterns of activity across a population of neurons that all subserve a similar function (for example, see REFS 142,143): these are termed neural population codes. A distributed representation of information of this type is more robust to the effects of noise. Many sensory systems form a spatially-ordered population — that is, a map — in which neighbouring neurons encode stimuli that share closely related features. Such spatially ordered populations support two basic goals of neural computation: first, a transformation between different maps (such as the direction of sounds into neck rotation) and, second, the combination of information from multiple sources (such as visual- and auditory-cue combination)144. The information capacity of a population of neurons is greatest when the noise sources across the population are not correlated. Noise correlations, which are often observed in populations of higher-order neurons, limit information capacity145,146 and have led to the development of population-coding strategies that account for the effects of correlations147.
The principles of averaging and prior knowledge can be placed into a larger mathematical framework of optimal statistical estimation and decision theory, known as Bayesian inference148. Bayesian inference assigns probabilities to propositions about the world (beliefs). These beliefs are calculated by combining prior knowledge (for example, that an animal is a predator) and noisy observations (for example, the heading of animal) to infer the probability of propositions (for example, animal attacks). Psychophysical experiments have confirmed that humans use these Bayesian inferences to allow them to cope with noise (and, more generally, with uncertainty) in both perception and action149,150. However, the neural mechanisms that are involved in Bayesian computations are unknown. One idea is that neurons encode probabilities or beliefs about the state of the world151,152, and this concept has been incorporated into Bayesian models of neuronal population codes144,153,154.
The above discussion has focused on the processing of information arriving simultaneously from multiple neurons or sensory modalities. However, information is often acquired over time, and in this case temporal averaging to be used to remove noise. For example, in signal-transduction systems, biochemical reaction time-constants could be set to make the duration of the reactions longer than the duration of the noise events — this would average-out random fluctuations155. Averaging over time can take place at the cellular level because of the temporal-integration properties of the membrane. These properties can be tuned by an appropriate choice of neuronal geometry156 and ion channels157 so that the characteristic bandwidth of the noise is strongly attenuated whereas the signal is not. Electrophysiological studies in the monkey have shown that behaviourally relevant signals are averaged not only across neuronal populations but also over time in the formation of a behavioural decision158.
In behaviour, temporal averaging is important when we need to estimate the current state or configuration of our limbs. Both the motor commands acting on our body and the sensory feedback containing information about the configuration of our body are noisy. Knowing the motor command allows us to predict the expected body configuration using an internal forward (predictive) model. However, such a prediction would deviate over time if sensory feedback were not available. The Kalman filter159 is an algorithm that combines noisy sensory feedback and the prediction from forward models to estimate the current configuration of our body over time. Kalman filtering in the CNS was demonstrated in behavioural studies of hand position160 and posture161.
In many cases the CNS has to choose a strategy by which it will achieve a goal through interaction with the environment. For example, in reaching the motor system has to specify a sequence of muscle activations to achieve a goal. However, there are many possible strategies to achieve a goal, and each might have a different associated cost (error, energy or time). Finding efficient strategies involves optimizing a cost function. For example, it has been proposed that we choose to move in a way that reduces the detrimental consequences of noise119. Stochastic optimal-control theory162 has emerged as a framework by which to study sensorimotor control. This theory makes several predictions that have been experimentally verified. For example, rather than specify a desired hand trajectory and use feedback to keep you on that trajectory, this theory proposes that optimal feedback control on task-relevant parameters is used: by allowing variation in parameters that do not affect the task, the system can behave in a more optimal manner. Stochastic optimal-feedback control is a beautiful example of how the principles of prior knowledge and averaging in motor behaviour are put to use biologically. This framework has been able to explain quantitative data from human and primate movements162-165. However, the neuronal substrate and mechanisms of such optimal controllers remain unknown.
Humans also use strategies that appear to increase noise. Confronted with higher movement-accuracy constraints (for example, when asked to rapidly point at small targets), people co-contract their muscles166, which increases joint stiffness105,167. However, greater activation of the muscles results in higher neuromuscular noise levels and is expected to produce larger movement variability. The reason that this is not the case lies in the dynamic properties of the muscles. In fact, movement variability decreases overall because the positive stabilizing effect of enhanced stiffness exceeds the negative effects of the increased force variability of the individual muscles105,167-169. Thus, human sensorimotor control takes account of noise to increase behavioural precision.
Conclusion
Noise has recently emerged as a key component of a wide range of biological systems — from gene expression98 to heart function89. In neuroscience, we have shown how noise is introduced at all stages of the sensorimotor loop, from the level of a single signalling protein to that of body movement. Noise has direct behavioural consequences, from setting perceptual thresholds to affecting movement precision. Although there has been an awareness of sensory noise for over half a century, cellular and motor noise have only recently received significant attention.
The question of the extent to which noise generates variability in the CNS is likely to require both experimental studies and stochastic modelling (in which each source of variability can be controlled for). We are beginning to develop a bottom-up understanding of how noise that is present at the molecular level (channel noise in membranes and biochemical noise at synapses) affects information processing at macroscopic levels (whole neurons, neuronal networks and behaviour). At all of these levels a key advance has been the use of stochastic models that can explain the experimentally observed variability and enable mechanisms to be characterized in a more detailed and often simpler manner than deterministic models (for example, see REFS 46,63,105,119,150). It has often been convenient to approximate noise by some additive random processes, such as Poisson or Gaussian processes, in which higher-order statistics beyond the mean (and the variance), as well as temporal structure, are ignored. This simple approach often forms the best assumption when data is lacking, and it simplifies the mathematical manipulation. However, it can result in noise levels being underestimated by several orders of magnitude in many small structures of the CNS63. Owing to the discrete nature of molecular noise and the nonlinearities that are present, noise can have a complex temporal structure, such as abrupt and large changes in noise level63, and spatial interactions can produce unexpected effects65,94. Advances in stochastic (Monte Carlo) simulations have made it possible to investigate in silico the nature and effects of noise in well known but previously deterministically described mechanisms (for example, see REFS 63,65,94,119).
The amount of noise that can be tolerated for a task depends on the required internal (such as long-term stability of memories) and behavioural (such as movement accuracy) performance. Noise levels set both hard limits on the CNS, such as the degree of miniaturization of the brain's circuits63, and soft constraints, such as the metabolic cost or the amount of time that is required to complete a task. For example, APs are noisy but also metabolically costly (mean neuronal firing rates in the cortex appear to be limited by energy supply170). Therefore, although neuronal communication becomes more reliable by using more APs, it also becomes more expensive. This trade-off has been observed in mammalian visual systems171,172. Another trade-off involves noise and time. For example, in pointing tasks, movement speed and pointing accuracy are inversely related (Fitt's law173), as faster movements require greater muscle forces, which are more noisy119. Therefore, noise is an integral part of the tradeoff between CNS resources (mass, size, time delays, et cetera) and performance which might ultimately determine evolutionary fitness.
Noise is an inescapable consequence of brains operating with molecular components at the nanometer scale, sensors that are sensitive to individual quanta and complex networks of noisy neurons that generate behaviour. The presence of noise in nervous systems has profound implications for their computational power174. Yet, despite significant noise levels our brain appears to function reliably, presumably because it has evolved under the constraints that are imposed by noise. Therefore, to understand the nervous system we have to distinguish variability from noise by accounting for its sources and appreciate the way in which it influences the brain's structure and function.
Glossary
- Fluctuation
A variation in a quantity from its spatial or temporal average.
- Noise
Random or unpredictable fluctuations and disturbances that are not part of a signal.
- Spike
An action potential interpreted as a unitary pulse signal (that is, it either is or is not present), the timing of which determines its information content. Other properties of the action potential, such as its shape or depolarization levels, are ignored.
- Trial-to-trial variability
The differences between responses that are observed when the same experiment is repeated in the same specimen (for example, in the same neuron or in the same subject).
- Poisson process
A random process that generates binary (yes or no) events for which the probability of occurrence for any small time interval is low. The rate at which events occur completely determines the statistics of the events. Poisson processes have a Fano factor of 1.
- Muscle spindle
Mechanosensory organ relaying muscle length and velocity.
- Fano factor
The ratio of the variance of a variable quantity to its mean.
- Stochastic process (random process)
A process that generates a series of random events.
- Positive feedback
Feedback that responds to a perturbation in the same direction as the perturbation, thereby amplifying its effect.
- Nodes of Ranvier
Regularly spaced gaps in the myelin sheath that surrounds a myelinated axon or nerve fibre. They expose the axonal membrane to the extracellular fluid and contain large numbers of voltage-gated ion channels and thus enable conduction of the action potential.
- Patch-clamp technique
An electrophysiological method that allows the study of the flow of current through a very small patch of cell membrane, which can contain just a single ion channel.
- Signal-to-noise ratio
The ratio of how much power is contained in the signal over the power of the noise, often measured as the variance of the signal divided by the variance of the noise.
- Axon hillock
The anatomical part of a (cortical) neuron that connects the cell body to the axon. Axon hillocks are the postulated primary site of action-potential initiation.
- Johnson noise (thermal noise , Johnson–Nyquist noise or Nyquist noise)
The electronic noise that is generated by the thermal agitation of the charge carriers (electrons and ions) inside an electrical conductor at equilibrium, which happens regardless of any applied of any applied voltage. Johnson noise is distinguished from shot noise, which consists of additional current fluctuations that occur when a voltage is applied to a resistance and a macroscopic current starts to flow.
- Shot noise
A type of noise that occurs at occurs when the finite number of particles that carry energy in a system, such as electrons or ions in an electrical circuit or photons in an optical device such as the photoreceptor, is small enough to give rise to detectable statistical fluctuations in a measurement.
- Ephaptic coupling
The coupling of very close or touching neurons, mediated by the electrical fields the neurons generate during electrical activity.
- Coefficient of variation(CV)
The ratio of the standard deviation of a variable quantity to its mean.
- Release probability
The probability of a vesicle being released during a synaptic-transmission event.
- Redundancy
The incorporation of surplus or duplicate information in a message to reduce the likelihood that noise can corrupt the message.
- Gaussian noise process
A random process that generates continuous events distributed with a Gaussian distribution. The mean and variance completely determines the statistics of events and there is no temporal correlation between events. Also known as white noise.
Footnotes
Competing interests statement
The authors declare no competing financial interests.
References
- 1.Adrian ED, Zotterman Y. The impulses produced by sensory nerve endings. II. The responses of a single end-organ. J. Physiol. 1926;61:151–171. doi: 10.1113/jphysiol.1926.sp002281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Tomko GJ, Crapper DR. Neuronal variability: non-stationary responses to identical visual stimuli. Brain Res. 1974;79:405–418. doi: 10.1016/0006-8993(74)90438-7. [DOI] [PubMed] [Google Scholar]
- 3.Tolhurst DJ, Movshon JA, Dean AF. The statistical reliability of signals in single neurons in cat and monkey visual cortex. Vision Res. 1983;23:775–785. doi: 10.1016/0042-6989(83)90200-6. [DOI] [PubMed] [Google Scholar]
- 4.Shadlen MN, Newsome WT. The variable discharge of cortical neurons: implications for connectivity, computation, and information coding. J. Neurosci. 1998;18:3870–3896. doi: 10.1523/JNEUROSCI.18-10-03870.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Shannon CE. A mathematical theory of communication. Bell Syst. Tech. J. 1948;27:373–423. 623–656. [Google Scholar]
- 6.von Neumann J. Probabilistic logics and the synthesis of reliable organisms from unreliable components. Automata Studies. 1956;34:43–99. [Google Scholar]
- 7.Berg HC, Purcell EM. Physics of chemoreception. Biophys. J. 1977;20:193–219. doi: 10.1016/S0006-3495(77)85544-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Bialek W, Setayeshgar S. Physical limits to biochemical signaling. Proc. Natl Acad. Sci. USA. 2005;102:10040–10045. doi: 10.1073/pnas.0504321102. This paper uses a first-principle approach to show that there are noise limits to biochemical signalling, which should apply to olfaction and intracellular signalling and at the synapse. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Bialek W. Physical limits to sensation and perception. Annu. Rev. Biophys. Biophys. Chem. 1987;16:455–478. doi: 10.1146/annurev.bb.16.060187.002323. [DOI] [PubMed] [Google Scholar]
- 10.Lillywhite PG, Laughlin SB. Transducer noise in a photoreceptor. Nature. 1979;277:569–572. doi: 10.1038/277569a0. [DOI] [PubMed] [Google Scholar]
- 11.Barlow HB, Levick WR, Yoon M. Responses to single quanta of light in retinal ganglion cells of the cat. Vision Res. 1971;11:87–101. doi: 10.1016/0042-6989(71)90033-2. [DOI] [PubMed] [Google Scholar]
- 12.Cover TM, Thomas JA. Elements of Information Theory. Wiley-Interscience; New York: 1991. [Google Scholar]
- 13.Laughlin SBA, John C, O'Carroll DC, de Ruyter van Steveninck RR. In: Information Theory and the Brain. Baddeley RHR, Foldiak R, editors. Cambridge Univ. Press; 2000. [Google Scholar]
- 14.Bryant HL, Segundo JP. Spike initiation by transmembrane current: a white-noise analysis. J. Physiol. 1976;260:279–314. doi: 10.1113/jphysiol.1976.sp011516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Mainen ZF, Sejnowski TJ. Reliability of spike timing in neocortical neurons. Science. 1995;268:1503–1506. doi: 10.1126/science.7770778. [DOI] [PubMed] [Google Scholar]
- 16.Harsch A, Robinson HPC. Postsynaptic variability of firing in rat cortical neurons: the roles of input synchronization and synaptic NMDA receptor conductance. J. Neurosci. 2000;20:6181–6192. doi: 10.1523/JNEUROSCI.20-16-06181.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Schreiber S, Fellous J-M, Tiesinga P, Sejnowski TJ. Influence of ionic conductances on spike timing reliability of cortical neurons for suprathreshold rhythmic inputs. J. Neurophysiol. 2004;91:194–205. doi: 10.1152/jn.00556.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.de Ruyter van Steveninck RR, Lewen GD, Strong SP, Koberle R, Bialek W. Reproducibility and variability in neural spike trains. Science. 1997;275:1805–1808. doi: 10.1126/science.275.5307.1805. [DOI] [PubMed] [Google Scholar]
- 19.Berry MJ, Warland DK, Meister M. The structure and precision of retinal spike trains. Proc. Natl Acad. Sci. USA. 1997;94:5411–5416. doi: 10.1073/pnas.94.10.5411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Rieke F, Warland D, van Steveninck R, Bialek W. MIT Press; Cambridge, Massachusetts: 1997. [Google Scholar]
- 21.Bair W. Spike timing in the mammalian visual system. Curr. Opin. Neurobiol. 1999;9:447–453. doi: 10.1016/S0959-4388(99)80067-1. [DOI] [PubMed] [Google Scholar]
- 22.Keat J, Reinagel P, Reid RC, Meister M. Predicting every spike: a model for the responses of visual neurons. Neuron. 2001;30:803–817. doi: 10.1016/s0896-6273(01)00322-1. [DOI] [PubMed] [Google Scholar]
- 23.Panzeri S, Petersen RS, Schultz SR, Lebedev M, Diamond ME. The role of spike timing in the coding of stimulus location in rat somatosensory cortex. Neuron. 2001;29:769–777. doi: 10.1016/s0896-6273(01)00251-3. [DOI] [PubMed] [Google Scholar]
- 24.Kara P, Reinagel P, Reid RC. Low response variability in simultaneously recorded retinal, thalamic and cortical neurons. Neuron. 2000;27:635–646. doi: 10.1016/s0896-6273(00)00072-6. [DOI] [PubMed] [Google Scholar]
- 25.Billimoria CP, DiCaprio RA, Birmingham JT, Abbott LF, Marder E. Neuromodulation of spike-timing precision in sensory neurons. J. Neurosci. 2006;26:5910–5919. doi: 10.1523/JNEUROSCI.4659-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hausser M, Spruston N, Stuart GJ. Diversity and dynamics of dendritic signaling. Science. 2000;290:739–744. doi: 10.1126/science.290.5492.739. [DOI] [PubMed] [Google Scholar]
- 27.Stuart G, Hausser M. Dendritic coincidence detection of EPSPs and action potentials. Nature Neurosci. 2001;4:63–71. doi: 10.1038/82910. [DOI] [PubMed] [Google Scholar]
- 28.Carr CE, Konishi M. A circuit for detection of interaural time differences in the brain stem of the barn owl. J. Neurosci. 1990;10:3227–3246. doi: 10.1523/JNEUROSCI.10-10-03227.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Fairhall AL, Lewen GD, Bialek W, de Ruyter van Steveninck RR. Efficiency and ambiguity in an adaptive neural code. Nature. 2001;412:787–792. doi: 10.1038/35090500. [DOI] [PubMed] [Google Scholar]
- 30.Zhurov Y, Brezina V. Variability of motor neuron spike timing maintains and shapes contractions of the accessory radula closer muscle of aplysia. J. Neurosci. 2006;26:7056–7070. doi: 10.1523/JNEUROSCI.5277-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Bialek W, Rieke F, de Ruyter van Steveninck RR, Warland D. Reading a neural code. Science. 1991;252:1854–1857. doi: 10.1126/science.2063199. [DOI] [PubMed] [Google Scholar]
- 32.Softky WR, Koch C. The highly irregular firing of cortical cells is inconsistent with temporal integration of random EPSPs. J. Neurosci. 1993;13:334–350. doi: 10.1523/JNEUROSCI.13-01-00334.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Strong SP, Koberle R, de Ruyter van Steveninck RR, Bialek W. Entropy and information in neural spike trains. Phys. Rev. Lett. 1998;80:197–200. [Google Scholar]
- 34.Heggelund P, Albus K. Response variability and orientation discrimination of single cells in striate cortex of cat. Exp. Brain Res. 1978;32:197–211. doi: 10.1007/BF00239727. [DOI] [PubMed] [Google Scholar]
- 35.Shadlen MN, Newsome WT. The variable discharge of cortical neurons: implications for connectivity, computation, and information coding. J. Neurosci. 1998;18:3870–3896. doi: 10.1523/JNEUROSCI.18-10-03870.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Gur M, Beylin A, Snodderly DM. Response variability of neurons in primary visual cortex (V1) of alert monkeys. J. Neurosci. 1997;17:2914–2920. doi: 10.1523/JNEUROSCI.17-08-02914.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.DeWeese MR, Wehr M, Zador AM. Binary spiking in auditory cortex. J. Neurosci. 2003;23:7940–7949. doi: 10.1523/JNEUROSCI.23-21-07940.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Berry MJ, Meister M. Refractoriness and neural precision. J. Neurosci. 1998;18:2200–2211. doi: 10.1523/JNEUROSCI.18-06-02200.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Warzecha A-K, Egelhaaf M. Variability in spike trains during constant and dynamic stimulation. Science. 1999;283:1927–1930. doi: 10.1126/science.283.5409.1927. [DOI] [PubMed] [Google Scholar]
- 40.Azouz R, Gray CM. Cellular mechanisms contributing to response variability of cortical neurons in vivo. J. Neurosci. 1999;19:2209–2223. doi: 10.1523/JNEUROSCI.19-06-02209.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Deweese MR, Zador AM. Shared and private variability in the auditory cortex. J. Neurophysiol. 2004;92:1840–1855. doi: 10.1152/jn.00197.2004. [DOI] [PubMed] [Google Scholar]
- 42.Calvin WH, Stevens CF. Synaptic noise and other sources of randomness in motoneuron interspike intervals. J. Neurophysiol. 1968;31:574–587. doi: 10.1152/jn.1968.31.4.574. [DOI] [PubMed] [Google Scholar]
- 43.Strassberg AF, DeFelice LJ. Limitation of the Hodgkin-Huxley formalism: effects of single channel kinetics on transmembrane voltage dynamics. Neural Comput. 1993;5:843–855. [Google Scholar]
- 44.Schneidman E, Freedman B, Segev I. Ion channel stochasticity may be critical in determining the reliability and precision of spike timing. Neural Comput. 1998;10:1679–1703. doi: 10.1162/089976698300017089. This paper shows that even when large numbers of stochastic ion channels are present in a neuron, fluctuations can become critical near the AP threshold. [DOI] [PubMed] [Google Scholar]
- 45.Hodgkin AL, Huxley AF. Quantitative description of membrane current and its application to conduction and excitation in nerve. J. Physiol. 1952;117:500–544. doi: 10.1113/jphysiol.1952.sp004764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Fatt P, Katz B. Some observations on biological noise. Nature. 1950;166:597–598. doi: 10.1038/166597a0. [DOI] [PubMed] [Google Scholar]
- 47.Calvin WH, Stevens CF. Synaptic noise as a source of variability in the interval between action potentials. Science. 1967;155:842–844. doi: 10.1126/science.155.3764.842. [DOI] [PubMed] [Google Scholar]
- 48.Lou X, Scheuss V, Schneggenburger R. Allosteric modulation of the presynaptic Ca2+ sensor for vesicle fusion. Nature. 2005;435:497–501. doi: 10.1038/nature03568. [DOI] [PubMed] [Google Scholar]
- 49.Derksen HE, Verveen AA. Fluctuations of resting neural membrane potential. Science. 1966;151:1388–1389. doi: 10.1126/science.151.3716.1388. [DOI] [PubMed] [Google Scholar]
- 50.Verveen AA, Derksen HE, Schick KL. Voltage fluctuations of neural membrane. Nature. 1967;216:588–589. doi: 10.1038/216588a0. [DOI] [PubMed] [Google Scholar]
- 51.Blair EA, Erlanger J. A comparison of the characteristics of axons through their individual electric responses. Am. J. Physiol. 1933;106:524–564. [Google Scholar]
- 52.Steinmetz PN, Manwani A, Koch C, London M, Segev I. Subthreshold voltage noise due to channel fluctuations in active neuronal membranes. J. Comput. Neurosci. 2000;9:133–148. doi: 10.1023/a:1008967807741. [DOI] [PubMed] [Google Scholar]
- 53.White JA, Rubinstein JT, Kay AR. Channel noise in neurons. Trends Neurosci. 2000;23:131–137. doi: 10.1016/s0166-2236(99)01521-0. [DOI] [PubMed] [Google Scholar]
- 54.van Rossum MC, O'Brien BJ, Smith RG. Effects of noise on the spike timing precision of retinal ganglion cells. J. Neurophysiol. 2003;89:2406–2419. doi: 10.1152/jn.01106.2002. [DOI] [PubMed] [Google Scholar]
- 55.Rubinstein JT. Threshold fluctuations in an N sodium channel model of the node of Ranvier. Biophys. J. 1995;68:779–785. doi: 10.1016/S0006-3495(95)80252-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Skaugen E, Wallow L. Firing behaviour in a stochastic nerve membrane model based upon the Hodgkin-Huxley equations. Acta Physiol. Scand. 1979;107:343–363. doi: 10.1111/j.1748-1716.1979.tb06486.x. [DOI] [PubMed] [Google Scholar]
- 57.Chow CC, White JA. Spontaneous action potentials due to channel fluctuations. Biophys. J. 1996;71:3012–3021. doi: 10.1016/S0006-3495(96)79494-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Diba K, Lester HA, Koch C. Intrinsic noise in cultured hippocampal neurons: experiment and modeling. J. Neurosci. 2004;24:9723–9733. doi: 10.1523/JNEUROSCI.1721-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Jacobson GA, et al. Subthreshold voltage noise of rat neocortical pyramidal neurones. J. Physiol. 2005;564:145–160. doi: 10.1113/jphysiol.2004.080903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Dorval AD, Jr, White JA. Channel noise is essential for perithreshold oscillations in entorhinal stellate neurons. J. Neurosci. 2005;25:10025–10028. doi: 10.1523/JNEUROSCI.3557-05.2005. This paper uses an elegant application of the dynamic-clamp technique to show that stochastic effects are required to describe CNS behaviour in vitro. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Kole MH, Hallermann S, Stuart GJ. Single Ih channels in pyramidal neuron dendrites: properties, distribution, and impact on action potential output. J. Neurosci. 2006;26:1677–1687. doi: 10.1523/JNEUROSCI.3664-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Schneidman E, Freedman B, Segev I. Ion channel stochasticity may be critical in determining the reliability and precision of spike timing. Neural Comput. 1998;10:1679–1703. doi: 10.1162/089976698300017089. [DOI] [PubMed] [Google Scholar]
- 63.Faisal AA, White JA, Laughlin SB. Ion-channel noise places limits on the miniaturization of the brain's wiring. Curr. Biol. 2005;15:1143–1149. doi: 10.1016/j.cub.2005.05.056. This paper shows that channel noise places a universal lower limit on neuron diameter, matching anatomical data across species. [DOI] [PubMed] [Google Scholar]
- 64.Rall W. Time constants and electrotonic length of membrane cylinders and neurons. Biophys. J. 1969;9:1483–1508. doi: 10.1016/S0006-3495(69)86467-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Faisal AA, Laughlin SB. Stochastic simulations on the reliability of action potential propagation in thin axons. PLoS Comput. Biol. 2007;3:e79. doi: 10.1371/journal.pcbi.0030079. This paper uses stochastic simulations to show that the many thin axons in the CNS are a source of spike-time variability that had been overlooked experimentally. It demonstrated that noise acts in a context-dependent manner and allows for a previously unknown mode of AP conduction. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Cecchi GA, et al. Noise in neurons is message dependent. Proc. Natl Acad. Sci. USA. 2000;97:5557–5561. doi: 10.1073/pnas.100113597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Fellous J-M, Tiesinga PHE, Thomas PJ, Sejnowski TJ. Discovering spike patterns in neuronal responsess. J. Neurosci. 2004;24:2989–3001. doi: 10.1523/JNEUROSCI.4649-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Horikawa Y. Noise effects on spike propagation in the stochastic Hodgkin-Huxley models. Biol. Cybern. 1991;66:19–25. doi: 10.1007/BF00196449. [DOI] [PubMed] [Google Scholar]
- 69.Kandel ER, Schwartz JH, Jessell TM. Principles of Neural Science. McGraw–Hill/Appleton & Lange; New York: 2000. [Google Scholar]
- 70.Debanne D. Information processing in the axon. Nature Rev. Neurosci. 2004;5:304–316. doi: 10.1038/nrn1397. [DOI] [PubMed] [Google Scholar]
- 71.Debanne D, Guerineau NC, Gahwiler BH, Thompson SM. Action-potential propagation gated by an axonal IA-like K+ conductance in hippocampus. Nature. 1997;389:286–289. doi: 10.1038/38502. [DOI] [PubMed] [Google Scholar]
- 72.Lecar H, Nossal R. Theory of threshold fluctuations in nerves. II. Analysis of various sources of membrane noise. Biophys. J. 1971;11:1068–1084. doi: 10.1016/S0006-3495(71)86278-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Manwani A, Koch C. Detecting and estimating signaals in noisy cable structures. I. Neuronal noise sources. Neural Comput. 1999;11:1797–1829. doi: 10.1162/089976699300015972. [DOI] [PubMed] [Google Scholar]
- 74.Jefferys JG. Nonsynaptic modulation of neuronal activity in the brain: electric currents and extracellular ions. Physiol. Rev. 1995;75:689–723. doi: 10.1152/physrev.1995.75.4.689. [DOI] [PubMed] [Google Scholar]
- 75.Szapiro G, Barbour B. Multiple climbing fibers signal to molecular layer interneurons exclusively via glutamate spillover. Nature Neurosci. 2007;10:735–742. doi: 10.1038/nn1907. [DOI] [PubMed] [Google Scholar]
- 76.Katz B, Miledi R. Membrane noise produced by acetylcholine. Nature. 1970;226:962–963. doi: 10.1038/226962a0. [DOI] [PubMed] [Google Scholar]
- 77.Kleppe IC, Robinson HPC. Correlation entropy of synaptic input-output dynamics. Phys. Rev. E Stat. Nonlin. Soft Matter Phys. 2006;74:041909. doi: 10.1103/PhysRevE.74.041909. [DOI] [PubMed] [Google Scholar]
- 78.Faure P, Kaplan D, Korn H. Synaptic efficacy and the transmission of complex firing patterns between neurons. J. Neurophysiol. 2000;84:3010–3025. doi: 10.1152/jn.2000.84.6.3010. [DOI] [PubMed] [Google Scholar]
- 79.Steriade M, McCormick DA, Sejnowski TJ. Thalamocortical oscillations in the sleeping and aroused brain. Science. 1993;262:679–685. doi: 10.1126/science.8235588. [DOI] [PubMed] [Google Scholar]
- 80.Cowan RL, Wilson CJ. Spontaneous firing patterns and axonal projections of single corticostriatal neurons in the rat medial agranular cortex. J. Neurophysiol. 1994;71:17–32. doi: 10.1152/jn.1994.71.1.17. [DOI] [PubMed] [Google Scholar]
- 81.Metherate R, Ashe JH. Ionic flux contributions to neocortical slow waves and nucleus basalis-mediated activation: whole-cell recordings in vivo. J. Neurosci. 1993;13:5312–5323. doi: 10.1523/JNEUROSCI.13-12-05312.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Destexhe A, Rudolph M, Fellous JM, Sejnowski TJ. Fluctuating synaptic conductances recreate in vivo-like activity in neocortical neurons. Neuroscience. 2001;107:13–24. doi: 10.1016/s0306-4522(01)00344-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Fellous JM, Rudolph M, Destexhe A, Sejnowski TJ. Synaptic background noise controls the input/output characteristics of single cells in an in vitro model of in vivo activity. Neuroscience. 2003;122:811–829. doi: 10.1016/j.neuroscience.2003.08.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Stuart GJ, Sakmann B. Active propagation of somatic action potentials into neocortical pyramidal cell dendrites. Nature. 1994;367:69–72. doi: 10.1038/367069a0. [DOI] [PubMed] [Google Scholar]
- 85.Wehr M, Zador AM. Balanced inhibition underlies tuning and sharpens spike timing in auditory cortex. Nature. 2003;426:442–446. doi: 10.1038/nature02116. [DOI] [PubMed] [Google Scholar]
- 86.Stevens CF, Zador AM. Input synchrony and the irregular firing of cortical neurons. Nature Neurosci. 1998;1:210–217. doi: 10.1038/659. [DOI] [PubMed] [Google Scholar]
- 87.Shu Y, Hasenstaub A, Badoual M, Bal T, McCormick DA. Barrages of synaptic activity control the gain and sensitivity of cortical neurons. J. Neurosci. 2003;23:10388–10401. doi: 10.1523/JNEUROSCI.23-32-10388.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Conti R, Tan Y, Llano I. Action potential-evoked and ryanodine-sensitive spontaneous Ca2+ transients at the presynaptic terminal of a developing CNS inhibitory synapse. J. Neurosci. 2004;24:6946–6957. doi: 10.1523/JNEUROSCI.1397-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Wang SQ, Song LS, Lakatta EG, Cheng H. Ca2+ signalling between single L-type Ca2+ channels and ryanodine receptors in heart cells. Nature. 2001;410:592–596. doi: 10.1038/35069083. [DOI] [PubMed] [Google Scholar]
- 90.Bekkers JM, Richerson GB, Stevens CF. Origin of variability in quantal size in cultured hippocampal neurons and hippocampal slices. Proc. Natl Acad. Sci. USA. 1990;87:5359–5362. doi: 10.1073/pnas.87.14.5359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Zucker RS, Regehr WG. Short-term synaptic plasticity. Annu. Rev. Physiol. 2002;64:355–405. doi: 10.1146/annurev.physiol.64.092501.114547. [DOI] [PubMed] [Google Scholar]
- 92.Sulzer D, Edwards R. Vesicles: equal in neurotransmitter concentration but not in volume. Neuron. 2000;28:5–7. doi: 10.1016/s0896-6273(00)00077-5. [DOI] [PubMed] [Google Scholar]
- 93.Wu X-S, et al. The origin of quantal size variation: vesicular glutamate concentration plays a significant role. J. Neurosci. 2007;27:3046–3056. doi: 10.1523/JNEUROSCI.4415-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Franks KM, Stevens CF, Sejnowski TJ. Independent sources of quantal variability at single glutamatergic synapses. J. Neurosci. 2003;23:3186–3195. doi: 10.1523/JNEUROSCI.23-08-03186.2003. This paper uses stochastic simulations to model the effects of random variability in the synapse, down to the level of individual transmitter molecules. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Nimchinsky EA, Yasuda R, Oertner TG, Svoboda K. The number of glutamate receptors opened by synaptic stimulation in single hippocampal spines. J. Neurosci. 2004;24:2054–2064. doi: 10.1523/JNEUROSCI.5066-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Nusser Z, Cull-Candy S, Farrant M. Differences in synaptic GABAA receptor number underlie variation in GABA mini amplitude. Neuron. 1997;19:697–709. doi: 10.1016/s0896-6273(00)80382-7. [DOI] [PubMed] [Google Scholar]
- 97.Lim R, Alvarez FJ, Walmsley B. Quantal size is correlated with receptor cluster area at glycinergic synapses in the rat brainstem. J. Physiol. 1999;516:505–512. doi: 10.1111/j.1469-7793.1999.0505v.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Paulsson J. Summing up the noise in gene networks. Nature. 2004;427:415–418. doi: 10.1038/nature02257. [DOI] [PubMed] [Google Scholar]
- 99.Abbott LF, Regehr WG. Synaptic computation. Nature. 2004;431:796–803. doi: 10.1038/nature03010. [DOI] [PubMed] [Google Scholar]
- 100.Miller P, Zhabotinsky AM, Lisman JE, Wang XJ. The stability of a stochastic CaMKII switch: dependence on the number of enzyme molecules and protein turnover. PLoS Biol. 2005;3:e107. doi: 10.1371/journal.pbio.0030107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Henneman E. Relation between size of neurons and their susceptibility to discharge. Science. 1957;126:1345–1347. doi: 10.1126/science.126.3287.1345. [DOI] [PubMed] [Google Scholar]
- 102.Schmidt R, Zelaznik H, Hawkins B, Frank J, Quinn J., Jr. Motor-output variability: a theory for the accuracy of rapid motor acts. Psychol. Rev. 1979;47:415–451. [PubMed] [Google Scholar]
- 103.Jones KE, Hamilton AF, Wolpert DM. Sources of signal-dependent noise during isometric force production. J. Neurophysiol. 2002;88:1533–1544. doi: 10.1152/jn.2002.88.3.1533. This paper provides experimental and theoretical evidence that the linear scaling of force variability (signal-dependent noise) is a natural by-product of the organization of motor neurons and muscle fibres (the size principle) [DOI] [PubMed] [Google Scholar]
- 104.Hamilton AF, Jones KE, Wolpert DM. The scaling of motor noise with muscle strength and motor unit number in humans. Exp. Brain Res. 2004;157:417–430. doi: 10.1007/s00221-004-1856-7. [DOI] [PubMed] [Google Scholar]
- 105.Selen LP, Beek PJ, van Dieen JH. Can co-activation reduce kinematic variability? A simulation study. Biol. Cybern. 2005;93:373–381. doi: 10.1007/s00422-005-0015-y. [DOI] [PubMed] [Google Scholar]
- 106.Moritz CT, Barry BK, Pascoe MA, Enoka RM. Discharge rate variability influences the variation in force fluctuations across the working range of a hand muscle. J. Neurophysiol. 2005;93:2449–2459. doi: 10.1152/jn.01122.2004. [DOI] [PubMed] [Google Scholar]
- 107.Christakos CN, Papadimitriou NA, Erimaki S. Parallel neuronal mechanisms underlying physiological force tremor in steady muscle contractions of humans. J. Neurophysiol. 2006;95:53–66. doi: 10.1152/jn.00051.2005. [DOI] [PubMed] [Google Scholar]
- 108.Moffitt MA, McIntyre CC, Grill WM. Prediction of myelinated nerve fiber stimulation thresholds: limitations of linear models. IEEE Trans. Biomed. Eng. 2004;51:229–236. doi: 10.1109/TBME.2003.820382. [DOI] [PubMed] [Google Scholar]
- 109.Mino H, Grill WM., Jr. Effects of stochastic sodium channels on extracellular excitation of myelinated nerve fibers. IEEE Trans. Biomed. Eng. 2002;49:527–532. doi: 10.1109/TBME.2002.1001966. [DOI] [PubMed] [Google Scholar]
- 110.Frank TD, Friedrich R, Beek PJ. Stochastic order parameter equation of isometric force production revealed by drift-diffusion estimates. Phys. Rev. E Stat. Nonlin. Soft Matter Phys. 2006;74:051905. doi: 10.1103/PhysRevE.74.051905. [DOI] [PubMed] [Google Scholar]
- 111.Capogrossi MC, Stern MD, Spurgeon HA, Lakatta EG. Spontaneous Ca2+ release from the sarcoplasmic reticulum limits Ca2+-dependent twitch potentiation in individual cardiac myocytes. A mechanism for maximum inotropy in the myocardium. J. Gen. Physiol. 1988;91:133–155. doi: 10.1085/jgp.91.1.133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Khatib M, Hilaire G, Monteau R. Excitatory interactions between phrenic motoneurons in the cat. Exp. Brain Res. 1986;62:273–280. doi: 10.1007/BF00238846. [DOI] [PubMed] [Google Scholar]
- 113.Chen H, Tourtellotte W, Frank E. Muscle spindle-derived neurotrophin 3 regulates synaptic connectivity between muscle sensory and motor neurons. J. Neurosci. 2002;22:3512–3519. doi: 10.1523/JNEUROSCI.22-09-03512.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Furness J. The excitatory input to a single smooth muscle cell. Pflugers Arch. 1970;314:1–13. doi: 10.1007/BF00587042. [DOI] [PubMed] [Google Scholar]
- 115.Morris LG, Hooper SL. Muscle response to changing neuronal input in the lobster (Panulirus interruptus) stomatogastric system: spike number-versus spike frequency-dependent domains. J. Neurosci. 1997;17:5956–5971. doi: 10.1523/JNEUROSCI.17-15-05956.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Lum CS, Zhurov Y, Cropper EC, Weiss KR, Brezina V. Variability of swallowing performance in intact, freely feeding aplysia. J. Neurophysiol. 2005;94:2427–2446. doi: 10.1152/jn.00280.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Hooper SL, Guschlbauer C, von Uckermann G, Buschges A. Natural neural output that produces highly variable locomotory movements. J. Neurophysiol. 2006;96:2072–2088. doi: 10.1152/jn.00366.2006. [DOI] [PubMed] [Google Scholar]
- 118.Ludlow CL. Central nervous system control of the laryngeal muscles in humans. Respir. Physiol. Neurobiol. 2005;147:205–222. doi: 10.1016/j.resp.2005.04.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Harris CM, Wolpert DM. Signal-dependent noise determines motor planning. Nature. 1998;394:780–784. doi: 10.1038/29528. This paper assumes that there is signal-dependent noise on the outgoing motor command and predicts the trajectories of goal-directed movements. The model suggests that subjects move in a way that minimizes the negative consequences of this noise. [DOI] [PubMed] [Google Scholar]
- 120.Harris CM, Wolpert DM. The main sequence of saccades optimizes speed-accuracy trade-off. Biol. Cybern. 2006;95:21–29. doi: 10.1007/s00422-006-0064-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.van Beers RJ. The sources of variability in saccadic eye movements. J. Neurosci. 2007;27:8757–8. doi: 10.1523/JNEUROSCI.2311-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Hamilton AF, Wolpert DM. Controlling statistics of action: obstacle avoidance. J. Neurophysiol. 2002;87:2434–2440. doi: 10.1152/jn.2002.87.5.2434. [DOI] [PubMed] [Google Scholar]
- 123.van Beers RJ, Haggard P, Wolpert DM. The role of execution noise in movement variability. J. Neurophysiol. 2004;91:1050–1063. doi: 10.1152/jn.00652.2003. [DOI] [PubMed] [Google Scholar]
- 124.Kozlov AS, Risler T, Hudspeth AJ. Coherent motion of stereocilia assures the concerted gating of hair-cell transduction channels. Nature Neurosci. 2007;10:87–92. doi: 10.1038/nn1818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.de Ruyter van Steveninck RR, Laughlin SB. The rate of information-transfer at graded-potential synapses. Nature. 1996;379:642–645. [Google Scholar]
- 126.Glowatzki E, Fuchs PA. Transmitter release at the hair cell ribbon synapse. Nature Neurosci. 2002;5:147–154. doi: 10.1038/nn796. [DOI] [PubMed] [Google Scholar]
- 127.Barlow H. In: Sensory Communication. Rosenblith WA, editor. MIT Press; Cambridge, Massachusetts: 1961. pp. 217–234. [Google Scholar]
- 128.Laughlin S. A simple coding procedure enhances a neuron's information capacity. Z. Naturforsch. [C] 1981;36:910–912. [PubMed] [Google Scholar]
- 129.Field D. Relations between the statistics of natural images and the response properties of cortical cells. J. Opt. Soc. Am. A. 1987;4:2379–2394. doi: 10.1364/josaa.4.002379. [DOI] [PubMed] [Google Scholar]
- 130.Adelman TL, Bialek W, Olberg RM. The information content of receptive fields. Neuron. 2003;40:823–833. doi: 10.1016/s0896-6273(03)00680-9. [DOI] [PubMed] [Google Scholar]
- 131.Turin G. An introduction to matched filters. IEEE Trans. Information Theory. 1960;6:311–329. [Google Scholar]
- 132.Barlow HB. The neurologic of matching filters. J. Opt. Technol. 1999;66:776–781. [Google Scholar]
- 133.Ernst MO, Banks MS. Humans integrate visual and haptic information in a statistically optimal fashion. Nature. 2002;415:429–433. doi: 10.1038/415429a. [DOI] [PubMed] [Google Scholar]
- 134.Alais D, Burr D. The ventriloquist effect results from near-optimal bimodal integration. Curr. Biol. 2004;14:257–262. doi: 10.1016/j.cub.2004.01.029. [DOI] [PubMed] [Google Scholar]
- 135.Jacobs RA. Optimal integration of texture and motion cues to depth. Vision Res. 1999;39:3621–3629. doi: 10.1016/s0042-6989(99)00088-7. This was one of the first studies to demonstrate statistically optimal weighting (weighted averaging) of sensory cues. [DOI] [PubMed] [Google Scholar]
- 136.Knill DC. Mixture models and the probabilistic structure of depth cues. Vision Res. 2003;43:831–854. doi: 10.1016/s0042-6989(03)00003-8. [DOI] [PubMed] [Google Scholar]
- 137.Hillis J, Watt S, Landy M, Banks M. Slant from texture and disparity cues: optimal cue combination. J. Vis. 2004;4:967–992. doi: 10.1167/4.12.1. [DOI] [PubMed] [Google Scholar]
- 138.Beers RJ, Sittig AC, Denier JJ. How humans combine simultaneous proprioceptive and visual position information. Exp. Brain Res. 1996;111:253–261. doi: 10.1007/BF00227302. [DOI] [PubMed] [Google Scholar]
- 139.van Beers RJ, Sittig AC, Denier van der Gon JJ. Integration of proprioceptive and visual position-information: an experimentally supported model. J. Neuurophysiol. 1999;81:1355–1364. doi: 10.1152/jn.1999.81.3.1355. This paper provided the first evidence of a strategy for the optimal integration of multi-dimensional cues. When integrating proprioception and visual information in the plane (two-dimensional cues), subjects use knowledge of direction-dependent variability to generate an optimal estimate. [DOI] [PubMed] [Google Scholar]
- 140.Sober S, Sabes P. Flexible strategies for sensory integration during motor planning. Nature Neurosci. 2005;8:490–497. doi: 10.1038/nn1427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Haruno M, Wolpert DM. Optimal control of redundant muscles in step-tracking wrist movements. J. Neurophysiol. 2005;94:4244–4255. doi: 10.1152/jn.00404.2005. [DOI] [PubMed] [Google Scholar]
- 142.Georgopoulos A, Schwartz A, Kettner R. Neuronal population coding of movement direction. Science. 1986;233:1416–1419. doi: 10.1126/science.3749885. [DOI] [PubMed] [Google Scholar]
- 143.Lee C, Rohrer WH, Sparks DL. Population coding of saccadic eye movements by neurons in the superior colliculus. Nature. 1988;332:357–360. doi: 10.1038/332357a0. [DOI] [PubMed] [Google Scholar]
- 144.Deneve S, Latham P, Pouget A. Efficient computation and cue integration with noisy population codes. Nature Neurosci. 2001;4:826–831. doi: 10.1038/90541. [DOI] [PubMed] [Google Scholar]
- 145.Zohary E, Shadlen MN, Newsome WT. Correlated neuronal discharge rate and its implications for psychophysical performance. 1994;370:140–143. doi: 10.1038/370140a0. [DOI] [PubMed] [Google Scholar]
- 146.Osborne LC, Bialek W, Lisberger SG. Time course of information about motion direction in visual area MT of macaque monkeys. J. Neurosci. 2004;24:3210–3222. doi: 10.1523/JNEUROSCI.5305-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Romo R, Hernandez A, Zainos A, Salinas E. Correlated neuronal discharges that increase coding efficiency during perceptual discrimination. Neuron. 2003;38:649–657. doi: 10.1016/s0896-6273(03)00287-3. [DOI] [PubMed] [Google Scholar]
- 148.Jaynes ET. In: Probability Theory: The Logic of Science. Bretthorst GL, editor. Cambridge Univ. Press; 2003. [Google Scholar]
- 149.Kording KP, Wolpert DM. Bayesian integration in sensorimotor learning. Nature. 2004;427:244–247. doi: 10.1038/nature02169. This was the first paper to demonstrate that subjects use task statistics (prior knowledge) and knowledge of their sensory variability to produce estimates that are Bayes optimal. [DOI] [PubMed] [Google Scholar]
- 150.Stocker AA, Simoncelli EP. Noise characteristics and prior expectations in human visual speed perception. Nature Neurosci. 2006;9:578–585. doi: 10.1038/nn1669. This paper provides a framework for testing how experimental variability can be used to infer prior probabilities as well as the internal noise characteristics directly from experimental data. [DOI] [PubMed] [Google Scholar]
- 151.Barlow HB. Pattern recognition and the responses of sensory neurons. Ann. NY Acad. Sci. 1969;156:872–881. doi: 10.1111/j.1749-6632.1969.tb14019.x. [DOI] [PubMed] [Google Scholar]
- 152.Barlow H. Single units and sensation: a neuron doctrine for perceptual psychology. Perception. 1972;1:371–394. doi: 10.1068/p010371. [DOI] [PubMed] [Google Scholar]
- 153.Paradiso M. A theory for the use of visual orientation information which exploits the columnar structure of striate cortex. Biol. Cybern. 1988;58:35–49. doi: 10.1007/BF00363954. [DOI] [PubMed] [Google Scholar]
- 154.Ma WJ, Beck JM, Latham PE, Pouget A. Bayesian inference with probabilistic population codes. Nature Neurosci. 2006;9:1432–1438. doi: 10.1038/nn1790. [DOI] [PubMed] [Google Scholar]
- 155.Hartwell LH, Hopfield JJ, Leibler S, Murray AW. From molecular to modular cell biology. Nature. 1999;402:C47–C42. doi: 10.1038/35011540. [DOI] [PubMed] [Google Scholar]
- 156.van Hateren JH, Laughlin SB. Membrane parameters, signal transmission, and the design of a graded potential neuron. J. Comp. Physiol. A. 1990;166:437–448. doi: 10.1007/BF00192015. [DOI] [PubMed] [Google Scholar]
- 157.Niven JE, et al. The contribution of Shaker K+ channels to the information capacity of Drosophila photoreceptors. Nature. 2003;421:630–634. doi: 10.1038/nature01384. [DOI] [PubMed] [Google Scholar]
- 158.Gold JI, Shadlen MN. Representation of a perceptual decision in developing oculomotor commands. Nature. 2000;404:390–394. doi: 10.1038/35006062. [DOI] [PubMed] [Google Scholar]
- 159.Goodwin G, Sin K. Adaptive filtering, prediction and control. Prentice-Hall; 1985. [Google Scholar]
- 160.Wolpert DM, Ghahramani Z, Jordan MI. An internal model for sensorimotor integration. Science. 1995;269:1880–1882. doi: 10.1126/science.7569931. [DOI] [PubMed] [Google Scholar]
- 161.Kuo A. An optimal control model for analyzing human postural balance. IEEE Trans. Biomed. Eng. 1995;42:87–101. doi: 10.1109/10.362914. [DOI] [PubMed] [Google Scholar]
- 162.Todorov E, Jordan MI. Optimal feedback control as a theory of motor coordination. Nature Neurosci. 2002;5:1226–1235. doi: 10.1038/nn963. This paper shows that an optimal control theory with signal-dependent noise in both the sensory feedback and the motor commands produces a number of observed motor behaviours. [DOI] [PubMed] [Google Scholar]
- 163.Todorov E. Optimality principles in sensorimotor control. Nature Neurosci. sci. 2004;7:907–915. doi: 10.1038/nn1309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Diedrichsen J. Optimal task-dependent changes of bimanual feedback control and adaptation. Curr. Biol. 2007;17:1675–1679. doi: 10.1016/j.cub.2007.08.051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Liu D, Todorov E. Evidence for the flexible sensorimotor strategies predicted by optimal feedback control. J. Neurosci. 2007;27:9354–9368. doi: 10.1523/JNEUROSCI.1110-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Gribble PL, Mullin LI, Cothros N, Mattar A. Role of cocontraction in arm movement accuracy. J. Neurophysiol. 2003;89:2396–2405. doi: 10.1152/jn.01020.2002. [DOI] [PubMed] [Google Scholar]
- 167.Selen LP, Beek PJ, van Dieen JH. Impedance is modulated to meet accuracy demands during goal-directed arm movements. Exp. Brain Res. 2006;172:129–138. doi: 10.1007/s00221-005-0320-7. This paper provided the first experimental evidence that subjects increase joint stiffness when confronted with higher accuracy demands in goal-directed movements. The increase in joint stiffness results in less kinematic variability. [DOI] [PubMed] [Google Scholar]
- 168.Selen LP, van Dieen JH, Beek PJ. Impedance modulation and feedback corrections in tracking targets of variable size and frequency. J. Neurophysiol. 2006;96:2750–2759. doi: 10.1152/jn.00552.2006. [DOI] [PubMed] [Google Scholar]
- 169.Lametti DR, Houle G, Ostry DJ. Control of movement variability and the regulation of limb impedance. J. Neurophysiol. 2007;98:3516–3524. doi: 10.1152/jn.00970.2007. [DOI] [PubMed] [Google Scholar]
- 170.Attwell D, Laughlin SB. An energy budget for signaling in the grey matter of the brain. J. Cereb. Blood Flow Metab. 2001;21:1133–1145. doi: 10.1097/00004647-200110000-00001. [DOI] [PubMed] [Google Scholar]
- 171.Balasubramanian V, Kimber D, Berry MJ. Metabolically efficient information processing. Neural Comput. 2001;13:799–815. doi: 10.1162/089976601300014358. [DOI] [PubMed] [Google Scholar]
- 172.de Polavieja GG. Errors drive the evolution of biological signalling to costly codes. J. Theor. Biol. 2002;214:657–664. doi: 10.1006/jtbi.2001.2498. [DOI] [PubMed] [Google Scholar]
- 173.Fitts PM. The information capacity of the human motor system in controlling the amplitude of movement. J. Exp. Psychol. 1954;47:381–391. [PubMed] [Google Scholar]
- 174.Maass W, Orponen P. On the effect of analog noise in discrete-time analog computations. Neural Comp. 1998;10:1071–1095. [Google Scholar]
- 175.Benzi R, Sutera A, Vulpiani A. The mechanism of stochastic resonance. J. Phys. A Math. Gen. 1981;14:L453–L457. [Google Scholar]
- 176.Longtin A, Bulsara A, Moss F. Time-interval sequences in bistable systems and the noise-induced transmission of information by sensory neurons. Phys. Rev. Lett. 1991;67:656–659. doi: 10.1103/PhysRevLett.67.656. [DOI] [PubMed] [Google Scholar]
- 177.Douglass JK, Wilkens L, Pantazelou E, Moss F. Noise enhancement of information transfer in crayfish mechanoreceptors by stochastic resonance. 1993;365:337–340. doi: 10.1038/365337a0. [DOI] [PubMed] [Google Scholar]
- 178.Braun HA, Wissing H, Schafer K, Hirsch MC. Oscillation and noise determine signal transduction in shark multimodal sensory cells. 1994;367:270–273. doi: 10.1038/367270a0. [DOI] [PubMed] [Google Scholar]
- 179.Levin JE, Miller JP. Broadband neural encoding in the cricket cereal sensory system enhanced by stochastic resonance. 1996;380:165–168. doi: 10.1038/380165a0. [DOI] [PubMed] [Google Scholar]
- 180.Cordo P, et al. Noise in human muscle spindles. 1996;383:769–770. doi: 10.1038/383769a0. [DOI] [PubMed] [Google Scholar]
- 181.Russell DF, Wilkens LA, Moss F. Use of behavioural stochastic resonance by paddle fish for feeding. 1999;402:291–294. doi: 10.1038/46279. This paper provided the first demonstration of stochastic resonance effects at the behavioural level. [DOI] [PubMed] [Google Scholar]
- 182.Priplata AA, Niemi JB, Harry JD, Lipsitz LA, Collins JJ. Vibrating insoles and balance control in elderly people. Lancet. 2003;362:1123–1124. doi: 10.1016/S0140-6736(03)14470-4. [DOI] [PubMed] [Google Scholar]
- 183.Anderson JS, Lampl I, Gillespie DC, Ferster D. The contribution of noise to contrast invariance of orientation tuning in cat visual cortex. Science. 2000;290:1968–1972. doi: 10.1126/science.290.5498.1968. [DOI] [PubMed] [Google Scholar]
- 184.Krogh A, Hertz JA. Generalization in a linear perceptron in the presence of noise. J. Phys. A Math. Gen. 1992;25:1135–1147. [Google Scholar]
- 185.Kirkpatrick S, Gelatt CD, Jr, Vecchi MP. Optimization by simulated annealing. Science. 1983;220:671–680. doi: 10.1126/science.220.4598.671. [DOI] [PubMed] [Google Scholar]
- 186.Tuckwell HC, Rodriguez R. Analytical and simulation results for stochastic Fitzhugh-Nagumo neurons and neural networks. J. Comput. Neurosci. 1998;5:91–113. doi: 10.1023/a:1008811814446. [DOI] [PubMed] [Google Scholar]
- 187.Lindner B, Garcia-Ojalvo J, Neiman A, Schimansky-Geier L. Effects of noise in excitable systems. Phys. Rep. 2004;392:321–424. [Google Scholar]
- 188.Turrigiano GG, Leslie KR, Desai NS, Rutherford LC, Nelson SB. Activity-dependent scaling of quantal amplitude in neocortical neurons. Nature. 1998;391:892–896. doi: 10.1038/36103. [DOI] [PubMed] [Google Scholar]
- 189.Prinz A, Bucher D, Marder E. Similar network activity from disparate circuit parameters. Nature Neurosci. 2004;7:1345–1352. doi: 10.1038/nn1352. [DOI] [PubMed] [Google Scholar]
- 190.Marsalek P, Koch C, Maunsell J. On the relationship between synaptic input and spike output jitter in individual neurons. Proc. Natl Acad. Sci. USA. 1997;94:735–740. doi: 10.1073/pnas.94.2.735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Poliakov AV, Powers RK, Binder MD. Functional identification of the input-output transforms of motoneurones in the rat and cat. J. Physiol. 1997;504:401–424. doi: 10.1111/j.1469-7793.1997.401be.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Sarpeshkar R. Analog versus digital: extrapolating from electronics to neurobiology. Neural Comp. 1998;9:1601–1638. doi: 10.1162/089976698300017052. [DOI] [PubMed] [Google Scholar]
- 193.Laughlin S, van Steveninck RR, Anderson J. The metabolic cost of neural information. Nature Neurosci. 1998;1:36–41. doi: 10.1038/236. [DOI] [PubMed] [Google Scholar]
- 194.Laughlin SB, Sejnowski TJ. Communication in neuronal networks. Science. 2003;301:1870–1874. doi: 10.1126/science.1089662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Laughlin SB. In: Complexity in Biological Information Processing. Bock G, Good J, editors. Wiley; Chichester: 2001. pp. 177–187. [Google Scholar]