

Abstract
Change-detection tasks reveal that infants’ ability to bind color to location in visual short-term memory (VSTM) develops rapidly: Seven-month-old infants, but not 6-month-old infants, detect that successive arrays of 3 objects are different if they contain the same colors in different locations (Oakes et al., 2006). Here we test a counterintuitive consequence of the hypothesis that six-month-old infants are unable to bind colors to locations: When comparing two successive stimulus arrays, these infants will often compare noncorresponding items, making it impossible for them to distinguish between identical arrays and nonidentical arrays. As a result, they will not show a preference for changing arrays over nonchanging arrays even when all of the items change. We tested this prediction by presenting 6- and 7-month-old infants (N = 36) with nonchanging displays of three items and changing displays in which all three items simultaneoulsy changed colors. As predicted, 7-month-old infants, but not 6-month-old infants, responded to the difference between these changing and nonchanging displays, providing additional evidence that the ability to bind colors to locations develops rapidly across this age range.
Understanding the early development of visual short-term memory (VSTM) is important for several reasons. For example, infants must encode and maintain information in VSTM for daily tasks such as comparing objects located on opposite sides of a room or recognizing the continuity of objects that go in and out of view. In addition, experimental procedures used to assess infants’ visual cognition often involve momentarily occluding some or all of the objects in the display (e.g., Baillargeon & Graber, 1988; Wilcox & Chapa, 2004; F. Xu, 2003). Therefore, we can best understand infants’ performance on such tasks by considering their emerging abilities to store and maintain information in short-term memory. Finally, studies documenting developmental changes in memory abilities can constrain theories about the neural and cognitive systems underlying visual short-term memory. For example, if the ability to store bound features in visual short-term memory has the same developmental time course as the ability to store unbound features, then this will suggest that both involve the same mechanism (see Vogel, Woodman, & Luck, 2001).
We use the phrase short-term memory rather than working memory because our experiments involve infant subjects, making it difficult to determine whether the memory system we are studying is used in the service of other tasks, which is the defining feature of a working memory system (Baddeley, 1986). However, the memory system we study is almost certainly a short-term memory system because our experimental paradigm uses conditions that are known to promote the use of short-term memory and to minimize the use of long-term memory (e.g., short encoding durations, brief retention intervals, and high levels of proactive interference). Moreover, this paradigm is quite similar to the change detection paradigm that has been used extensively to study VSTM in adults (Vogel et al., 2001).
Using the paradigm illustrated in Figure 1, we have documented clear developmental changes in VSTM over the first 12 months. In this paradigm, infants are simultaneously presented with two stimulus streams and their looking at each one is recorded. Each stream contains a specific number of objects, which cycle on and off over a period of many seconds (as in the flicker version of the change detection paradigm – see Simons & Rensink, 2005). In our streams, the array of objects is on for 500 ms, off for 250–300 ms, on for 500 ms, and so on. On one monitor infants see a changing stream, in which some aspect of the array changes from cycle to cycle; for example, a randomly chosen item might change color between cycle N and cycle N+1 (as in Figure 1C). On another monitor, infants see a nonchanging stream in which the arrays are identical from cycle to cycle (as in Figure 1A). Using this general paradigm, we can determine the conditions under which infants prefer to look at a changing stream more than to a simultaneously presented nonchanging stream. It is well established that infants prefer (i.e., look longer at) visual arrays that are more complex, or contain more elements (Cohen, 1991), and that they prefer dynamic stimuli to static ones (Shaddy & Colombo, 2004). Consequently, they will prefer a changing stream to a nonchanging stream if they can detect that the changing stream is changing and that the nonchanging stream is not. Given that a gap of 250–300 ms separates consecutive arrays within a stream, the infants will be able to distinguish between the changing and nonchanging streams only if they are able to store some of the information from a given array in VSTM and compare it with the corresponding information from the subsequent array. The combination of the 250–300 ms gap and the large transient caused by the onset of each array make it impossible to use iconic memory to solve this task. That is, even though the iconic memory of a given array will not have completely faded after 250–300 ms, the transient caused by the next array will eliminate this iconic memory so that it cannot be used to detect changes. Indeed, Rensink, O’Regan, and Clark (1997) found that iconic memory could not be used to perform change detection even when the delay was only 80 ms (see also Becker, Pashler, & Anstis, 2000). In addition, because the stimuli are brief and change frequently, infants cannot easily use long-term memory representations to detect the changes in the changing stream (see Ross-Sheehy, Oakes, & Luck, 2003).
Figure 1.

Sequence of events in several different types of stimulus streams: A. Nonchanging streams in which the array is identical from cycle to cycle; B. One-change streams at set size one, in which the arrays contain only one object and that one object changes color from cycle to cycle (Ross-Sheehy et al., 2003); C. One-change streams at set size 3, in which arrays contain three differently colored objects, and the color of a different randomly chosen object changes on each cycle (Ross-Sheehy et al., 2003); D. Binding change streams, in which arrays contain three differently colored objects, and the color-location bindings (but not the particular colors or the particular locations) change from cycle to cycle (Oakes et al., 2006); E. Homogeneous change cycles, in which arrays contain three identically colored objects, and the colors of all the objects change from cycle to cycles (Oakes et al., 2006); and F. All colors change streams, in which arrays contain three differently colored objects, and the colors of all the objects change from cycle to cycle. In addition, to the right of each cycle, we present what we know about whether infants prefer each type of changing stream when it is paired with a non-changing stream (illustrated in A. for each of the comparisons except for the homogeneous change streams in which were paired with corresponding homogeneous no-change streams).
Using this procedure, Ross-Sheehy, Oakes, and Luck (2003) found that 6-month-old infants detected a change (as evidenced by a preference for changing streams) when there was 1 item in each array (Figure 1B), but they did not detect a change (i.e., they showed no preference for the changing stream) when there were 2 or more items on each array (Figure 1C). Thus, although these infants exhibited the ability to remember the color of a single item in VSTM, they appeared to be unable to remember the colors of multiple objects. Ten-month-old infants, in contrast, significantly preferred the streams in which one randomly chosen item changed color to nonchanging streams when there were 1, 2, 3, or 4 items in each stream (Figure 1B,C, Ross-Sheehy et al., 2003), but they showed no significant preference at set size 6. Thus, VSTM abilities appear to undergo dramatic change in the first year of life—infants’ ability to detect changes in multi-object arrays develops between 6- and 10-months of age, perhaps reaching adult-like level of 3–4 objects by 10 months of age (other work has lead to similar estimates of the number of objects infants at this age can store, e.g. Cheries, Wynn, & Scholl, 2006; Feigenson, 2007), although it is difficult to determine their precise capacity with confidence using our procedure (see Cowan et al., 2005 for an extensive discussion of the development of VSTM capacity across development).
The failure of the 6-month-old infants to detect change in multi-object arrays is particularly noteworthy because infants could, in principle, notice changes in an array of two or more objects even if they stored only one of the objects in VSTM. That is, if they remembered 1 of the N items in the array, and if the color of 1 randomly chosen item changes on each cycle, they should be able to detect 1/N of the changes in the changing stream. If infants did adopt this strategy, however, their preference would be small at set size 3 because they would detect only 1/3 of the changes, and their preference may not be statistically significant. The 6-month-old infants observed by Ross-Sheehy et al. (2003) did not show a gradually decreasing preference as a function of set size (as predicted by their having detected 1/N changes); rather their preference was robust at set size one and near chance for both set sizes 2 and 3. Thus 6-month-old infants’ failure to show a significant preference for changing arrays containing two or more objects may not solely reflect a limited storage capacity, but rather may reflect some other difficulty that arises when multiple objects are present. In particular, infants may have difficulty binding object features to specific locations in the array. That is, they may represent the object features and the object locations, but without any link between the features and their locations.
A failure of object-location binding would make it difficult for infants to differentiate between changing and nonchanging streams of multi-element stimulus arrays because of the correspondence problem that must be solved in this task. Specifically, determining whether or not a given item changed between array N and array N+1 requires that the infant is able to determine which objects in array N correspond to which objects in array N+1 so that the corresponding objects can be compared. If infants are unable to bind color and location they may compare noncorresponding objects and perceive a change even when no change occurred.
Consider, for example, the nonchanging stream in Figure 1A. An infant who remembers the red item from cycle N-1 but does not remember the location of this item may compare the remembered red color with the visible blue item in cycle N (i.e., comparing noncorresponding objects), thus perceiving a change when there was no change. Thus, infants who are unable to bind object identity to location will have difficulty differentiating changing from non-changing arrays at set sizes of 2 or greater, even if they could in principle store multiple object identities in VSTM.
Oakes, Ross-Sheehy, and Luck (2006) demonstrated rapid developmental changes in infants’ ability to bind information in VSTM. In this study, the changing stream contained the same colors and same locations on each cycle, but the locations of the colors swapped from cycle to cycle (Figure 1D). Six-month-old infants failed to differentiate changing from nonchanging streams in this experiment, whereas 7-month-old infants strongly preferred the changing side. Kaldy and Leslie (2003; 2005) reported converging results for developmental changes in binding in this same age range. Thus the ability to bind color and location in VSTM develops rapidly around 7 months of age. One might be concerned that in our procedure the 6-month-olds could detect the color-location binding changes but that these changes were too subtle to have an impact on looking time. However, in this experiment all three items in the changing stream swapped locations on every cycle, and the changes should have been very salient if the colors were bound to the locations. Thus, the 6-month-old infants showed no preference for changing streams even though every location in the changing stream changed color each time the array reappeared. If the infants had been able to remember just one color at one location, they would have been able to detect the change on every cycle. These results indicate that the 6-month-old infants’ lack of preference for the changing side was not solely a result of limited VSTM storage capacity. Instead, they were unable to bind the color of an object to its location in VSTM. In contrast, 7-month-old and 12-month-old infants showed a significant preference for the changing arrays, indicating that they were able to bind color and location in VSTM.
Two control experiments demonstrated that 6-month-old infants are not simply overwhelmed by the presence of multiple items. In one experiment, all the items in a given array of three items were the same color, but the color of these three items changed from cycle to cycle in the changing stream (Figure 1E). Six-month-old infants showed a strong preference for the changing side in this experiment, demonstrating that they can use VSTM in the face of multiple-object stimulus arrays when no color-location binding is required to detect a change. In a second control experiment, each array in one stream contained three identical colors and each array in the other stream contained three different colors. Six-month-old infants preferred the arrays containing three different colors, demonstrating that they could discriminate the color differences. Moreover, in tasks with exposure times of tens of seconds (or more), by 6 months infants can recognize color-location bindings (e.g., Catherwood, Skoien, Green, & Holt, 1996) suggesting that they both can perceive such relations and that they can store them in long-term memory. Thus, the failure of 6-month-old infants to show a preference for streams in which all color-location bindings are swapped on every trial is best explained by an inability to bind color and location together in VSTM, and the success of 7-month-old infants in this task indicates that the ability to bind color and location develops rapidly between 6 and 8 months of age.
The present study was designed to extend these previous findings and test a counterintuitive prediction that arises from our hypothesis that the ability to bind objects to locations develops over this period. Specifically, we hypothesized that an inability to bind color and location would lead to an inability to differentiate between changing and nonchanging streams even if every item in the changing stream changed to a completely new color on each cycle (see Figure 1F). This prediction is counterintuitive because 6-month-old infants have a VSTM capacity of at least one item, which should make it trivial to detect a change when every item changes to a color that was absent in the previous cycle. Such a failure would be remarkable given that memory for a single item should be sufficient to perform this task. However, memory for a single item will be useful only if it contains location information so that this memory can be compared to the appropriate item. Without the ability to bind color and location, infants will be unable to determine which item in the currently visible array should be compared with any items they have stored in VSTM from the previous array. As a result, even if infants retain one color in VSTM they are likely to see changes even in the nonchanging stream and it should be extremely difficult for them to distinguish between changing and nonchanging arrays containing multiple items, even if all the feature values vary from cycle to cycle in the changing stream.
Because 7-month-old infants, in contrast, can bind color and location, they should be able to determine that the nonchanging arrays are in fact nonchanging, and they should find it trivial to determine that the changing arrays are changing given that every item changes to a new color on every cycle. Thus, we predicted that 7-month-olds would prefer the changing stream but 6-month-old infants would not. Such a finding would provide converging evidence for rapid developmental change in VSTM binding, which would suggest that changes in neural structures in this period underlie the ability to form color-location bindings in VSTM.
Method
Participants
Thirty-six healthy infants with no history of birth complications, vision problems, or risk of colorblindness were tested. They included 18 6.5-month-old infants (M = 193 days, SD = 5.43) and 18 7.5-month-olds (M = 231 days, SD = 3.93) (19 boys and 17 girls). The majority of the infants were from White, middle-class families and were recruited using our usual procedures (Ross-Sheehy et al., 2003). An additional 4 infants were tested but were excluded from the final analysis because of experimenter error (1 7.5-month-old), equipment failure (1 6.5-month-old), or extreme side bias (1 6.5-month-old, 1 7.5-month-old).
Apparatus
A Macintosh G3 computer was used to present the stimuli on two 17-in. ViewSonic monitors, each with viewable surface of 18.26 degrees (w) by 13.5 degrees (h) at a distance of 100 cm. The two monitors were positioned side by side with a 24 cm (13.5 degrees) gap between them. The total width of the two displays and gap was approximately 88 cm (47.5 degrees). With this setup, infants could detect that there was a stimulus on the non-fixated display while attending to a fixated monitor, but they could not perceive the details of the non-fixated display due to the >30° distance between the centers of the monitors, which would have led to extensive crowding within the non-fixated display (Pelli, Palomares, & Majaj, 2004). Thus, when performing this task, the infants are focused on one monitor at any given moment rather than attending to both stimulus streams at the same time.
Stimuli
Each array contained three colored squares (2.9 × 2.9 degrees) on a gray background. Each of the three squares was a different color, selected from a set of eight highly discriminable values (red, green, cyan, white, yellow, black, brown, and blue). The colors within a display were always different from each other. During each 20-s trial, the array of colored squares on each monitor continually appeared and disappeared (visible for 500 ms, off for 300 ms, visible for 500 ms, etc.). For the changing stream, the colors of all three squares changed between disappearing and reappearing; the new values were selected at random on each cycle from the colors that were not present in the previous cycle. For the nonchanging stream, the colors of all three squares remained the same.
Procedure
Infants sat on a parent’s lap 100 cm in front of a large black curtain that obscured their view of the experimental apparatus. Holes were cut in the curtain for the 2 computer monitors, a small black box positioned between the monitors, and the lens of a video camera below the black box. To direct infants’ attention between the two monitors at the start of each trial, and thereby reduce side bias, the black box produced a flashing light and beeping tone between trials.
Infants received six 20-s trials in which a changing stream was presented on one monitor and a nonchanging stream was presented on the other. The left-right position of the changing stream was counterbalanced across trials (i.e., all infants received 3 trials with the changing stream on the right and three trials with the changing stream on the left). A trained observer, unaware of the location of the changing stream, recorded the looking time to each monitor online, and a second trained observer recoded the looking time off-line for a randomly chosen 25% of the infants. The average correlation between the two observers’ looking times was high (r = .98), with an average difference of 0.42 s.
Results
Our main dependent variable was the change-preference score, which was calculated by dividing the duration of looking to the changing stream by the total time spent looking at either of the two streams for a given trial (i.e., excluding time spent looking somewhere other than the two streams). The mean scores, and the mean looking scores used to derive them, are presented in Table 1.
Table 1.
Average looking times and change preference scores in s (SD in parentheses) by age.
| Age | Changing streams | Non-changing streams | Change Preference |
|---|---|---|---|
| 6 months | 4.72 (2.38) | 4.46 (1.70) | .50 (.09) |
| 7 months | 6.30 (1.87) | 4.81 (1.36) | .57 (.05) |
To determine whether infants discriminate the changing and nonchanging streams, we compared the change-preference score at each age to chance (.50) with a two-tailed t-test. If infants can determine that only the changing stream involves a change, they should prefer it and their change-preference score should be greater than chance. If infants are unable to determine that only the changing stream involves a change, they should look equivalently at the changing and nonchanging streams and their change-preference score should not be different from chance. The mean novelty preference score (with 95% confidence intervals) and individual scores for each infant are presented in Figure 2. It is clear that 7-month-old infants, but not 6-month-old infants, differentiated the two types of streams. Indeed, 7-month-old infants’ change-preference score was significantly greater than chance, t (17) = 3.37, p = .004, whereas 6-month-old infants’ change-preference score did not differ from chance, t (17) = −.17, p = .87.1
Figure 2.
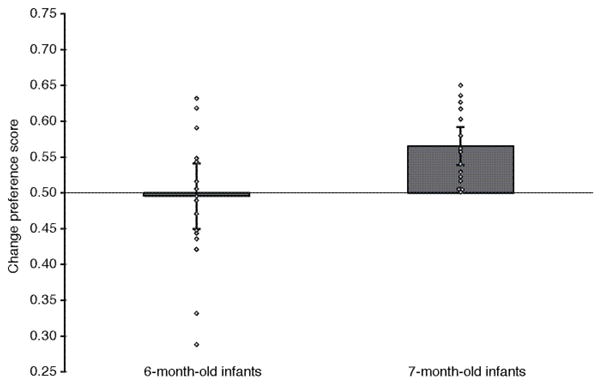
Mean change preference score (error bars represent 95% confidence intervals) and individual infants’ change preference scores by age.
To determine whether the preference score of the 7-month-old infants was significantly greater than that of the 6-month-old infants, we compared the preference scores across age. This comparison also was significant, t (34) = 2.15, p = .039, confirming that not only did 7-month-old infants have a preference score that was greater than chance, but they also preferred the changing side more than did the 6-month-old infants.
Finally, we compared the proportion of 6-month-old infants whose preference score was greater than .50 (9 of 18) to the proportion of 7-month-old infants whose preference score was greater than .50 (18 of 18). There was a significantly higher number of 7-month-old infants than 6-month-old infants who had scores greater than .50, chi-squared (df = 1) = 12.00, p = .001.
Discussion
This experiment reveals a striking pattern of findings: 6-month-old infants, but not 7-month-old infants, failed to prefer a changing stream at set size 3 even though all the items changed to a new color on every cycle. This is a remarkable failure, especially given that 6-month-old infants show a robust and replicable preference for changing streams that consist of a single object or three identically colored objects. Together, this work reveals that at 6 months infants can represent a single object—or a single object feature—but are unable to detect changes when multiple object identities must be stored and compared. After only one more month of development, however, infants could detect the changes with ease. The mean change preference score of the 7-month-old infants in the present experiment was 0.57, which means that they spent 33% more time looking toward the changing stream than toward the nonchanging stream, even though they were provided with no external incentives for doing so. This preference was highly significantly greater than what would be expected by chance, and it was also significantly greater than the preference exhibited by the 6-month-old infants, which was not significantly greater than what would be expected by chance. The timing of this difference in performance is precisely the timing of the dramatic change we previously observed in a different test of VSTM binding (Oakes et al., 2006), providing converging evidence for a key developmental change in this age range.
Detecting changes in the present paradigm seems trivially easy—infants need only remember a single object and compare it to the corresponding object in the next array to detect the change. However, this comparison requires that infants can determine which items correspond across the arrays, a process that requires binding. That is, recognizing that the red object has become green or remained red requires that the observer can remember which object was the red one, which in turn requires remembering where it was located. The inability of 6-month-old infants to prefer the changing stream in the present study therefore indicates that they either lacked information about the color-location bindings or were unable to use this information. Infants at this age do show a preference when each stream contains a single item or three identical items, ruling out a lack of storage capacity or an inability to process multi-element arrays as explanations for this result.
It should be noted that color-location bindings are not strictly necessary, in a formal sense, to perform the present task. An observer who could remember only the colors could, in principle, determine whether a change is present or absent by comparing a representation of the statistical properties of one array (e.g., the mean color of the entire array) with those of the next array. Indeed, adult observers can extract and compare some statistical properties, such as size (Chong & Treisman, 2003, 2005), and this ability may influence VSTM performance (J. S. Johnson, Hollingworth, & Luck, in press). There were large changes in the statistical properties of the arrays in the changing streams of the present experiment on every cycle, giving the 6-month-old infants every opportunity to use representations of these statistical properties and show a preference for the changing streams. Their failure to do so indicates that they either do not form these representations or do not use them to control their looking behavior. Moreover, 7-month-old infants do not solely rely on statistical properties because they prefer changing arrays both when they involve large changes in statistical properties (as in the present experiment) and when they involve no changes in statistical properties (as in the binding-changing experiment illustrated in Figure 1D, Oakes et al., 2006). Further research is necessary to determine whether observers of any age can use statistical information to perform tasks such as this.
One might also suppose that 6-month-old infants—who appear to be able to store a single item in VSTM—would be able to detect changes in the present paradigm by simply focusing attention onto a single location, storing this item in VSTM, and comparing it with the next item that appears at that location. Their lack of a preference for the changing stream therefore might reflect a problem with attention rather than a problem with binding, per se. However, by 6 months of age, attention can effectively perform simple stimulus selection operations such as this, and there is no evidence of a rapid transition in attentional selection between 6 and 8 months (Colombo, 2001). Thus, although we cannot completely rule out an attentional explanation of our findings, it is unlikely. Moreover, the binding of features to locations appears to require attention (Hyun, Woodman, & Luck, in press), so a failure of attention may be viewed as a failure of one major component of binding.
Kaldy and Leslie (2003; 2005) also observed that the ability to bind object location to identity changes in the second half of the first year. However, Kaldy and Leslie argue that at 6 months infants can bind, but they have a limited capacity to do so—specifically, they can bind only a single object and location. Thus, for Kaldy and Leslie, binding and capacity develop somewhat independently. We argue, in contrast, that infants’ ability to detect changes in multi-object arrays emerges from their ability to bind; that is, binding allows infants to compare the corresponding items in multi-object arrays, which is a necessary prerequisite for detecting changes, and thus the ability to bind features and to represent multiple objects are intertwined whenever the task requires comparing arrays separated by a gap.
These two different explanations have different implications for the brain mechanisms involved in binding. That is, although solving Kaldy and Leslie’s task may involve working memory (i.e., infants must update and maintain a representation of an object on each trial), the encoding and retention periods may be sufficiently long to engage long-term memory representations2. Indeed, they argued that their results reflected maturation of the medial temporal lobes, which appear to play a role in binding only with delays that exceed 1 second (Olson, Page, Moore, Chatterjee, & Verfaellie, 2006). This system may be able to bind objects to locations by 6 months of age given a sufficiently long encoding period3.
In contrast, binding in our task requires rapid encoding, and infants must retain information over a very short period of time. Recent evidence suggests that VSTM representations are maintained by means of interactions between the intraparietal sulcus (IPS) and inferotemporal cortex (ITC). Both of these regions exhibit sustained activity during the delay period of change-detection tasks, but IPS activity seems to be more closely tied to storage capacity and to location binding (Todd & Marois, 2004; Vogel & Machizawa, 2004; Y. Xu & Chun, 2006). The parietal cortex has also been linked to the binding of objects and locations in multielement stimulus arrays in other contexts. For example, an fMRI study by Shafriz et al. (2002) found that parietal regions were activated when subjects were required to bind the features of multiple objects presented simultaneously. In addition, Balint’s syndrome patients, who have bilateral parietal damage, are unable to bind object identity to location (Robertson, 2003). Thus, the kind of binding required in our task seems to be more closely related to the kind of binding that engages parietal cortex.
Indeed, consistent with our behavioral findings, several studies using a variety of methods suggest increases in connectivity and activity in parietal regions until 3 months after birth, and a gradual decrease approaching asymptotic change by approximately 6 months (e.g., Chugani, 1998; M. H. Johnson, Mareschal, & Csibra, 2001). These data are obviously suggestive and should be considered with caution—establishing the developmental time course of changes in neuroanatomy in infancy is extremely difficult. Nevertheless, these conclusions are supported by behavioral findings such as those presented here that infants 6 months and younger have difficulty with tasks that appear to involve parietal cortex in adults. Thus, the developmental changes of changes in infants’ ability to bind features in VSTM before 6 months may be related to the immaturity of parietal structures that support these abilities in adults.
Acknowledgments
This research and preparation of this manuscript were made possible by NIH grant HD49840 awarded to LMO; SRS was funded by predoctoral and postdoctoral NRSAs (HD055040, MH068934). We thank Shaena Stille, Kristine Kovack-Lesh, and the undergraduate students in the Infant Perception and Cognition Laboratory at the University of Iowa for their help with this project. Correspondences should be addressed to Lisa M. Oakes, Center for Mind and Brain, 267 Cousteau Pl., University of California, Davis, CA 95618 (e-mail: lmoakes@ucdavis.edu).
Footnotes
One might expect that the individual 6-month-old infants would all have preference scores approximating .50. However, although most infants had scores near .50, a few had substantially higher or lower scores (see Figure 2). We examined the data of these infants carefully to see if we could find a clear explanation for their divergent scores, but we found no obvious differences between these infants and the infants whose scores were nearer to chance (e.g., they did not differ in age from the other infants and they generally looked at both streams on each trial). It is therefore likely that their divergent scores reflect measurement noise rather than true preferences for the changing or nonchanging streams.
Substantial evidence supports the existence of separable short-term and long-term visual memory systems, which differ in terms of both their functional characteristics and their neural implementation (reviewed in Luck & Hollingworth, in press). These systems may operate concurrently in many tasks that use relatively short retention intervals (< 10 s) but also use relatively long encoding periods (> 500 ms).
It should also be noted that the pattern of performance exhibited by the 6-month-old infants in the experiments of Kaldy and Leslie (2003; 2005) is also consistent with infants simply remembering the identity of the last item shown, with no binding of identity and location.
References
- Baddeley AD. Working Memory. Oxford: Clarendon; 1986. [Google Scholar]
- Baillargeon R, Graber M. Evidence of location memory in 8-month-old infants in a nonsearch AB task. Developmental Psychology. 1988;24:502–511. [Google Scholar]
- Becker MW, Pashler H, Anstis SM. The role of iconic memory in change-detection tasks. Perception. 2000;29:273–286. doi: 10.1068/p3035. [DOI] [PubMed] [Google Scholar]
- Catherwood D, Skoien P, Green V, Holt C. Assessing the primary moments of infant encoding of compound visual stimuli. Infant Behavior & Development. 1996;19:1–11. [Google Scholar]
- Cheries EW, Wynn K, Scholl BJ. Interrupting infants’ persisting object representations: An object-based limit? Developmental Science. 2006;9:F50–F58. doi: 10.1111/j.1467-7687.2006.00521.x. [DOI] [PubMed] [Google Scholar]
- Chong SC, Treisman A. Representation of statistical properties. Vision Research. 2003;43:393–404. doi: 10.1016/s0042-6989(02)00596-5. [DOI] [PubMed] [Google Scholar]
- Chong SC, Treisman A. Statistical processing: Computing the average size in perceptual groups. Vision Research. 2005;45:891–900. doi: 10.1016/j.visres.2004.10.004. [DOI] [PubMed] [Google Scholar]
- Chugani HT. A critical period of brain development: studies of cerebral glucose utilization with PET. Preventative Medicine. 1998;27:184–188. doi: 10.1006/pmed.1998.0274. [DOI] [PubMed] [Google Scholar]
- Cohen LB. Infant attention: An information processing approach. In: Weiss MJ, Zelazo PR, editors. Newborn attention: Biological constraints and the influence of experience. Norwood, NJ: Ablex; 1991. pp. 1–21. [Google Scholar]
- Colombo J. The development of visual attention in infancy. Annual Review of Psychology. 2001;52:337–367. doi: 10.1146/annurev.psych.52.1.337. [DOI] [PubMed] [Google Scholar]
- Cowan N, Elliott EM, Saults JS, Morey CC, Mattox S, Hismjatullina A, et al. On the capacity of attention: Its estimation and its role in working memory and cognitive aptitudes. Cognitive Psychology. 2005;51:42–100. doi: 10.1016/j.cogpsych.2004.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Currie C, McConkie G, Carlson-Radvansky LA, Irwin DE. The role of the saccade target object in the perception of a visual stable world. Perception & Psychophysics. 2000;62:673–683. doi: 10.3758/bf03206914. [DOI] [PubMed] [Google Scholar]
- Feigenson L. Continuity of format and computation in short-term memory development. In: Oakes LM, Bauer PJ, editors. Short- and long-term memory in infancy: Taking the first steps toward remembering. New York: Oxford University Press; 2007. pp. 51–74. [Google Scholar]
- Hollingworth A, Richard AM, Luck SJ. Understanding the function of visual short-term memory: Transsaccadic memory, object correspondence, and gaze correction. Journal of Experimental Psychology: General. doi: 10.1037/0096-3445.137.1.163. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hyun J-S, Woodman GF, Luck SJ. The role of attention in the binding of surface features to locations. Visual Cognition. doi: 10.1080/13506280802113894. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson JS, Hollingworth A, Luck SJ. The role of attention in the maintenance of feature bindings in visual short-term memory. Journal of Experimental Psychology: Human Perception and Performance. doi: 10.1037/0096-1523.34.1.41. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson MH, Mareschal D, Csibra G. The functional development and integration of the dorsal and ventral visual pathways: A neurocomputational approach. In: Nelson CA, Luciana M, editors. The Handbook of Developmental Cognitive Neuroscience. Cambridge, MA: MIT Press; 2001. pp. 339–351. [Google Scholar]
- Kaldy Z, Leslie AM. Identification of objects in 9-month-old infants: Integrating ‘what’ and ‘where’ information. Developmental Science. 2003;6:360–373. [Google Scholar]
- Kaldy Z, Leslie AM. A memory span of one? Object identification in 6.5-month-old infants. Cognition. 2005;97:153–177. doi: 10.1016/j.cognition.2004.09.009. [DOI] [PubMed] [Google Scholar]
- Kovack-Lesh KA, Oakes LM. Hold your horses: How exposure to different items influences infant categorization. Journal of Expeimental Psychology. 2007;2:69–93. doi: 10.1016/j.jecp.2007.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luck SJ, Hollingworth A, editors. Visual memory. New York: Oxford University Press; in press. [Google Scholar]
- Oakes LM, Ribar RJ. A comparison of infants’ categorization in paried and successive presentation familairization tasks. Infancy. 2005;7 doi: 10.1207/s15327078in0701_7. [DOI] [PubMed] [Google Scholar]
- Oakes LM, Ross-Sheehy S, Luck SJ. Rapid development of binding in visual short-term memory. Psychological Science. 2006;17:781–787. doi: 10.1111/j.1467-9280.2006.01782.x. [DOI] [PubMed] [Google Scholar]
- Olson IR, Page K, Moore KS, Chatterjee A, Verfaellie M. Working memory for conjunctions relies on the medial temporal lobe. Journal of Neuroscience. 2006;26:4596–4601. doi: 10.1523/JNEUROSCI.1923-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pelli DG, Palomares M, Majaj N. Crowding is unlike ordinary masking: Distinguishing feature integration from detection. Journal of Vision. 2004;4:1136–1169. doi: 10.1167/4.12.12. [DOI] [PubMed] [Google Scholar]
- Rensink RA, O’Regan J, Clark JJ. To see or not to see: The need for attention to perceive changes in scenes. Psychological Science. 1997;8:368–373. [Google Scholar]
- Robertson LC. Binding, spatial attention and perceptual awareness. Nature Reviews Neuroscience. 2003;4:93–102. doi: 10.1038/nrn1030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ross-Sheehy S, Oakes LM, Luck SJ. The development of visual short-term memory capacity in infants. Child Development. 2003;74:1807–1822. doi: 10.1046/j.1467-8624.2003.00639.x. [DOI] [PubMed] [Google Scholar]
- Shaddy DJ, Colombo J. Developmental changes in infant attention to dynamic and static stimuli. Infancy. 2004;5:355–365. [Google Scholar]
- Shafritz KM, Gore JC, Marois R. The role of the parietal cortex in visual feature binding. Proceedings of the National Academy of Science. 2002;99:10917–10922. doi: 10.1073/pnas.152694799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simons DJ, Rensink RA. Change blindness: Past, present, and future. Trends in Cognitive Sciences. 2005;9:16–20. doi: 10.1016/j.tics.2004.11.006. [DOI] [PubMed] [Google Scholar]
- Todd JJ, Marois R. Capacity limit of visual short-term memory in human posterior parietal cortex. Nature. 2004;428:751–754. doi: 10.1038/nature02466. [DOI] [PubMed] [Google Scholar]
- Vogel EK, Machizawa MG. Neural activity predicts individual differences in visual working memory capacity. Nature. 2004;428:748–751. doi: 10.1038/nature02447. [DOI] [PubMed] [Google Scholar]
- Vogel EK, Woodman GF, Luck SJ. Storage of features, conjunctions, and objects in visual working memory. Journal of Experimental Psychology. 2001;27:92–114. doi: 10.1037//0096-1523.27.1.92. [DOI] [PubMed] [Google Scholar]
- Wilcox T, Chapa C. Priming infants to attend to color and pattern information in an individuation task. Cognition. 2004;90:265–302. doi: 10.1016/s0010-0277(03)00147-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu F. The development of object individuation in infancy. In: Hayne H, Fagen JW, editors. Progress in infancy research. Vol. 3. NJ, US: Lawrence Erlbaum Associates, Publishers; 2003. pp. 159–192. [Google Scholar]
- Xu Y, Chun MM. Dissociable neural mechanisms supporting visual short-term memory for objects. Nature. 2006 doi: 10.1038/nature04262. [DOI] [PubMed] [Google Scholar]