

Abstract
Neuroglobin (Ngb) is a recently discovered tissue globin with a high affinity for oxygen that is widely and specifically expressed in neurons of vertebrate central and peripheral nervous systems. Our laboratory and others have showed Ngb over-expression can protect neurons against hypoxic/ischemic insults, but the underlying mechanisms remain poorly understood. In this study, we examined the effects of Ngb over-expression on mitochondrial function, oxidative stress and neurotoxicity in primary cortical neurons following hypoxia/reoxygenation (H/R). Ngb over-expressing transgenic neurons (Ngb-Tg) were significantly protected against H/R-induced cell death. Rates of decline in ATP levels, MTT reduction and mitochondrial membrane potential were significantly ameliorated in Ngb-Tg neurons. Furthermore, Ngb overexpression reduced superoxide anion generation after H/R, whereas glutathione levels were significantly improved compared to WT controls. Taken together, these data suggest that Ngb is neuroprotective against hypoxia, in part by improving mitochondria function and decreasing oxidative stress.
Keywords: Neuroglobin, Oxidative Stress, Hypoxia, Mitochondria, Neuroprotection
Introduction
Neuroglobin (Ngb) is a recently discovered tissue globin with a high affinity for oxygen that is widely expressed in vertebral central and peripheral nerve systems as well as retina and endocrine tissues (Awenius et al. 2001; Burmester and Hankeln 2004; Burmester et al. 2000; Garry and Mammen 2003; Zhang et al. 2001). As a newly discovered member of globin family, Ngb has been considered as the brain or nerve equivalent of tissue haemoglobin (Mammen et al. 2002). Previous studies from the Greenberg laboratory and our laboratory demonstrated that over-expression of Ngb is neuroprotective against hypoxic/ischemic brain injuries (Khan et al. 2007; Sun et al. 2001; Wang et al. 2008).
However, how Ngb protects neurons from hypoxia/ischemia still remains unclear (Greenberg et al. 2008). In general, tissue globins mediate multiple cellular and molecular responses to hypoxic/ischemic insults. For example, myoglobin in cardiomyocytes and oxidative skeletal myofibers helps facilitate oxygen transport, maintain nitric oxide homoeostasis, and scavenge reactive oxygen species (Garry et al. 2003; Marcinek et al. 2003; Flögel U et al. 2004). It is possible that Ngb has similar actions in brain. Importantly, Ngb expression is confined to metabolically most active, oxygen-consuming cell types (Burmester and Hankeln 2004). At the subcellular level, Ngb is associated with the presence of mitochondria and thus linked to the oxidative metabolism (Burmester et al. 2007). By immunohistochemistry, the neuroglobin immunoreactivity is restricted to the cytoplasma of neurons (Geuens et al. 2003); double immunostaining showed a largely overlapping, but slightly shifted labeling pattern in the inner segments of retina (Bentmann et al. 2005). A most recent study showed that Ngb over-expression eliminated hypoxia-induced mitochondrial aggregation and neuron death (Khan et al. 2008). Those reports suggest subcellular colocalization and functional association of Ngb to mitochondria. Furthermore, it has been well demonstrated that mitochondria comprises a central locus for energetic perturbations and oxidative stress in hypoxia/ischemia (Nicholls and Budd 2000; Sims and Anderson 2002). Therefore, in this study we used primary cortical neurons isolated from our newly created Ngb overexpressing transgenic mice, to test the hypothesis that Ngb promotes neuron survival in part by improving mitochondrial function and reducing oxidative stress after hypoxic/reoxygenation insults.
Materials and Methods
All animal experiments were performed following protocols approved by the Massachusetts General Hospital Institutional Animal Care and Use Committee in compliance with the NIH Guide for the Care and Use of Laboratory Animals.
Western blot
At 9 days of culture, cortical neurons were collected for protein extraction. Levels of Ngb protein expression were examined by western blotting following a previously described protocol (Wang et al. 2008). Optical density of Ngb protein bands was quantified with NIH Image software.
Primary cultures of Ngb-Tg and matching wild-type mouse cortical neurons
Primary mouse cortical neurons were isolated from 15 days embryonic cortex obtained from the pregnant Ngb-Tg (heterozygous) female mouse (mated with male wild type C57 BL/6 mouse). Neurons from each embryo cortex were isolated and seeded into multiple well plates separately with equal cell amounts. Non-cortical embryonic tissues were used for DNA extraction and PCR genotyping, in terms of identifying Ngb-Tg neurons and wild-type littermate neurons. Neurons from one brain cortex (totally about 4−5× 106 cells) were seeded onto 20 wells in 24 well plate or 4 wells in 6 well plates, and maintained in Neurobasal medium (NBM) with 3% B27 and 0.3 mM glutamine (Invitrogene). Medium was half changed in each 4 days. At 9−10 days after culture and before hypoxia, the media was changed with pre-warmed NBM supplemented with 0.1% B27 (AO free) for 10−12 hrs, and the cortical neurons were maintained in the media for the duration of all experiments. The purity of neurons was over 90% after 8 days of initial seeding examined by NeuN and GFAP immunostaining.
Hypoxia/reoxygenation
Hypoxia was induced with a modular chamber (Billups-Rothenber) perfused with 90% N2/5% CO2/5% H2 for 25 min at 37°C as previously described (Lee and Lo 2003). The chamber was then sealed and placed at 37 °C for indicated time periods of hypoxia. At the end of the hypoxia exposure, cells were removed from the chamber and maintained in a regular incubator for reoxygenation. Control cultures were incubated under normoxic conditions for the corresponding duration.
Neurotoxicity
Two assays, lactate dehydrogenase (LDH) release and live cell counts (by calcein AM staining) were used to measure neurotoxicity. LDH release is an indicator of plasma membrane damage and commonly used for the determination of neurotoxicity as we previously described (Wang et al. 2002). Primary cortical neurons were cultured onto 24-well plates for 9−10 days. LDH release was measured after hypoxia for 10 hrs followed by reoxygenation for 2 hrs, 6 hrs and 14 hrs. Live cell counting was performed after 10 hrs hypoxia followed by 14 hrs reoxygenation. Staining with calcein AM was based upon intracellular esterase activity which can be used to determine survival cells of cultured cortical neurons. Cells were washed with pre-warmed neurobasal medium, and incubated with 5 μM calcein AM (Molecular Probes) at room temperature for 10 min, then washed twice with pre-warmed neurobasal medium. Under fluorescence microscopy, cells with bright yellow cytoplasm were identified as living cells. Three fields of fluorescent images per culture well were taken randomly under 200X magnification. Live cell counts and quantifications were assessed thereafter. Results were expressed as percentage of normal controls.
ATP levels
Intracellular ATP levels in cultured cortical neurons were measured with an ATP Assay Kit (Calbiochem) on a microplate luminometer (LB 96V, Berthold) according to the manufacturer's instructions. Live cell counting with calcein AM staining was performed from sister culture wells and used to normalize measurements to avoid reflection of different survival rates. The same normalization by live cell counting was also applied to the following measurements of MTT reduction, mitochondrial membrane potential, glutathione detection, and superoxide anion generation.
MTT reduction
(3-(4, 5-dimethylthiazolyl-2)-2, 5-diphenyltetrazolium bromide) (MTT) reduction by viable mitochondria was used as an indicator of neuronal viability and mitochondria function, as previously described (Wang et al. 2002). MTT reduction was measured after hypoxia for 10 hrs followed by reoxygenation for 2 hrs and 6 hrs. Percentages of live cell counts were used for assay normalization.
Mitochondrial membrane potential
Mitochondrial membrane potential (ΔΨm) is a commonly used surrogate biomarker for mitochondrial function. In this study, a fluorescent lipophilic cationic dye, JC-1 (5,5’,6,6’-tetrachloro-1,1’,3,3’-tetraethylbenzimidazol carbocyanine iodide) mitochondrial membrane potential detection kit (Cell Technology) was used to measure ΔΨm according to the manufacturer's instructions with minor modification. Briefly, primary cortical neurons were cultured onto 96-well plates with black-wall flat bottom for 9−10 days. After 10 hrs hypoxia followed by reoxygenation or 2 hrs, the cultures were incubated with 5 μM JC-1 for 15 min at 37°C; then washed twice with phenol free neurobasal medium. Fluorescent signals of ΔΨm with excitation 545 nm and emission 620 nm were read on a microplate fluorescence reader (FL600, Bio-Tek). The phenol free neurobasal medium in non-cell wells served as internal and blank controls for both hypoxia treated and non-treated normal control plates. Results were expressed as reduction percentage of normal controls. Percentages of live cell counts were used for assay normalization.
Superoxide anion generation
Generation of intracellular superoxide anion in cultured cortical neurons was determined according to fluorescence of ethidium as a result of oxidation of hydroethidine (Oxi-HE). Oxi-HE formation is a biomarker for ROS production. After hypoxia for 10 hrs, 10 μM hydroethidine (Molecular Probes) in dimethyl sulphoxide (DMSO) was added into culture wells. 2 hrs later, media were aspirated and replaced with PBS containing 1% Triton-X100. Fluorescence with excitation 480 nm and emission 590 nm was read on a microplate fluorescence reader (FL600, Bio-Tek). Percentages of live cell counts were used for assay normalization.
Glutathione detection
Intracellular levels of glutathione (GSH) in cultured cortical neurons were measured after 10 hrs hypoxia followed by reoxygenation for 2 hrs with ApoGSH™ Glutathione Detection Kit (Biovision) according to the manufacturer's instructions. Fluorescent signals of GSH with excitation 380 nm and emission 460 nm were read on a microplate fluorescence reader (FL600, Bio-Tek). Percentages of live cell counts were used for assay normalization.
Statistical analysis
ANOVA followed by Tukey-Kramer posthoc tests were used to analyze data for Ngb protein expression, LDH release, MTT reduction, ATP levels, ΔΨm, oxi-HE formations, and GSH levels. Differences at P<0.05 were considered statistically significant.
Results
Reduction of neurotoxicity in Ngb-Tg neurons after H/R
Primary cortical neuronal cultures were derived from Ngb-Tg and WT mouse brains. Levels of Ngb protein expression in Ngb-Tg cortical neurons were about 2.6 fold higher than WT neurons, examined by western blot (Figure 1). After hypoxia for 10 hrs, neurotoxicity in WT and Ngb-Tg neurons was compared with the LDH release assay at 2, 6, and 14 hrs after reoxygenation. Although there were no significant differences at the earlier 2 and 6 hr time points, neurotoxicity after 14 hrs reoxygenation was significantly decreased by about 25% in Ngb-Tg neurons compared to WT neurons (Figure 2A). In addition, neurotoxicity was also evaluated by live cell count assay with calcein AM staining. The percentage of live neurons was significantly increased in Ngb-Tg neurons compared to WT controls (Figure 2B), which was consistent with the LDH findings.
Figure 1.
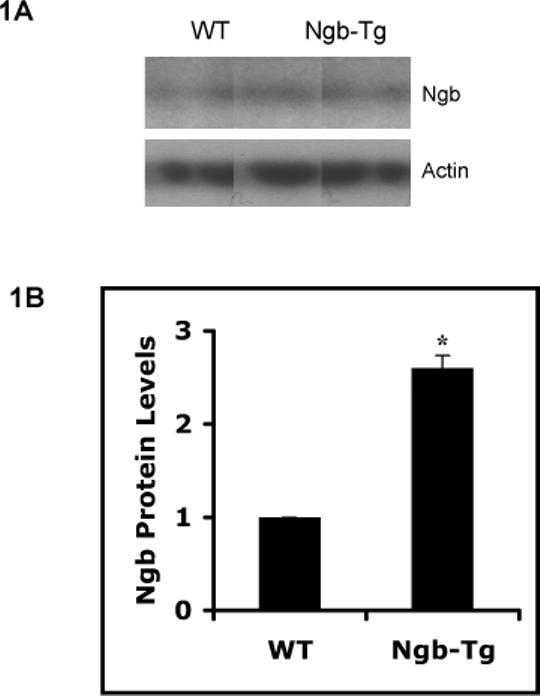
Increase of Ngb protein expression in Ngb-Tg cortical neuron cultures. A. Representative western blot showed Ngb-Tg neurons had higher Ngb protein levels compared to WT controls. Actin served as equal loading controls. B. Relative Ngb protein expression levels were quantified by optical density of Ngb protein bands. Mean±SEM, n=3 per group, *P<0.05.
Figure 2.

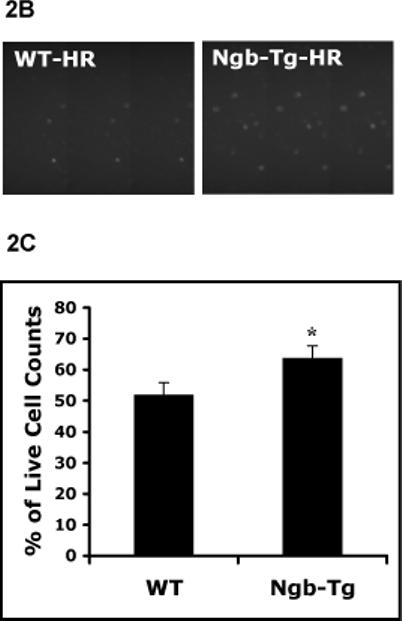
Reduction of neurotoxicity in Ngb over-expressing neurons after H/R. A. Primary cortical neuronal cultures were derived from 15 days old embryonic cortex of Ngb-Tg and WT brains. Time course of neurotoxicity after H/R was examined with LDH release assay. There was no significant difference in neurotoxicity after hypoxia 10 hrs followed by reoxygenation for 2 hrs (H10/R2) and 6 hrs (H10/R6), but Ngb-Tg neurons significantly reduced neurotoxicity after reoxygenation for 14 hrs (H10/R14), compared to WT neurons. Mean±SEM, n=5 per group, *P<0.05. B. Live cell count assay was used to measure neurotoxicity after 10 hr hypoxia followed by 14 hr reoxygenation. Up panel showed representative images of calcein AM staining identified increased live neurons in Ngb-Tg neuron cultures after H/R compared with WT controls. Lower panel showed quantification of the percentage of live cell counts was significantly increased in Ngb-Tg compared to WT neuron cultures. Mean±SEM, n=10 per group, *P<0.05.
Amelioration of mitochondrial disruption in Ngb-Tg neurons
Three biomarkers for mitochondrial function were assessed, including intracellular ATP levels, MTT reduction, and mitochondrial membrane potential (ΔΨm). In WT neurons, both ATP levels and rates of MTT reduction were markedly disrupted after 10 hrs of hypoxia followed by reoxygenation. In contrast, Ngb-Tg neurons demonstrated significantly preserved mitochondrial function. ATP levels were improved during the early periods (2 and 6 hrs) of reoxygenation after hypoxia (Figure 3A). Declines in MTT reduction were ameliorated at all time points of reoxygenation (2 and 6 hrs) after hypoxia (Figure 3B). Perturbations in mitochondrial function were also manifested as a loss of membrane potential. After 10 hrs hypoxia followed by 2 hr reoxygenation, ΔΨ values were reduced by approximately 40−50%. Over-expression of Ngb significantly ameliorated the loss of ΔΨ compared to WT controls (Figure 3C).
Figure 3.

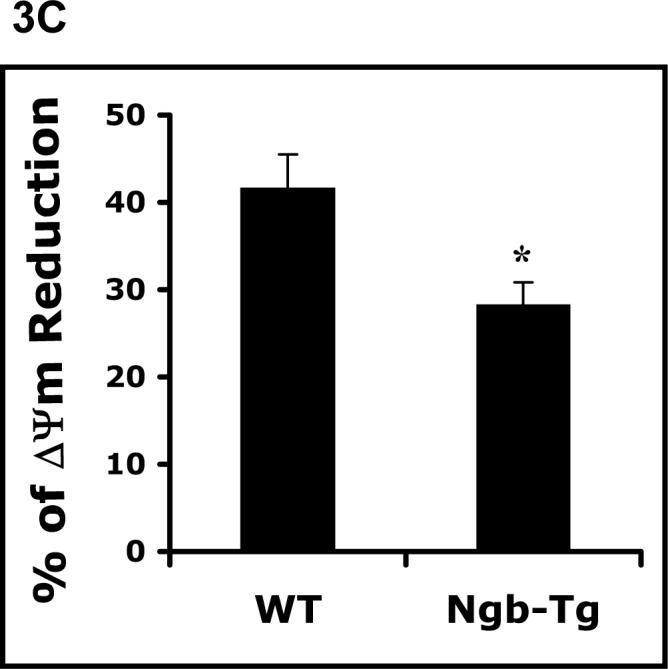
Amelioration of mitochondrial disruption in Ngb over-expressing neurons after H/R. A. Time-course of ATP levels was examined after hypoxia 10 hrs (H10), and followed by repxygenation for 2 hrs and 6 hrs. Levels of ATP in Ngb-Tg neurons were significantly higher at all three time points compared with WT neurons. B. Time- course of MTT reduction was measured after hypoxia 10 hrs followed by reoxygenation for 2 hrs (H10/R2), and 6 hrs (H10/R6). MTT reduction in Ngb-Tg neurons were significantly increased at the two time points compared with WT neurons. Mean±SEM, n=4, *p<0.05. C. ΔΨm was measured after 10 hr hypoxia followed by 2 hr reoxygenation, the ΔΨm reduction rate in Ngb-Tg neurons was significantly inhibited compared to WT controls. Mean±SEM, n=10 per group, *p<0.05.
Reduction of oxidative stress in Ngb-Tg neurons after HR
Oxi-HE formation and levels of endogenous GSH were measured as indirect markers of oxidative stress. Early oxidative stress was evident in WT neurons after 10 hrs of hypoxia and 2 hrs of re-oxygenation. Oxi-HE formation was significantly ameliorated in Ngb-Tg neurons compared to WT neurons (Figure 4A). Similarly, GSH was rapidly diminished in WT neurons, whereas in Ngb-Tg neurons, GSH level was comparatively preserved (Figure 4B).
Figure 4.
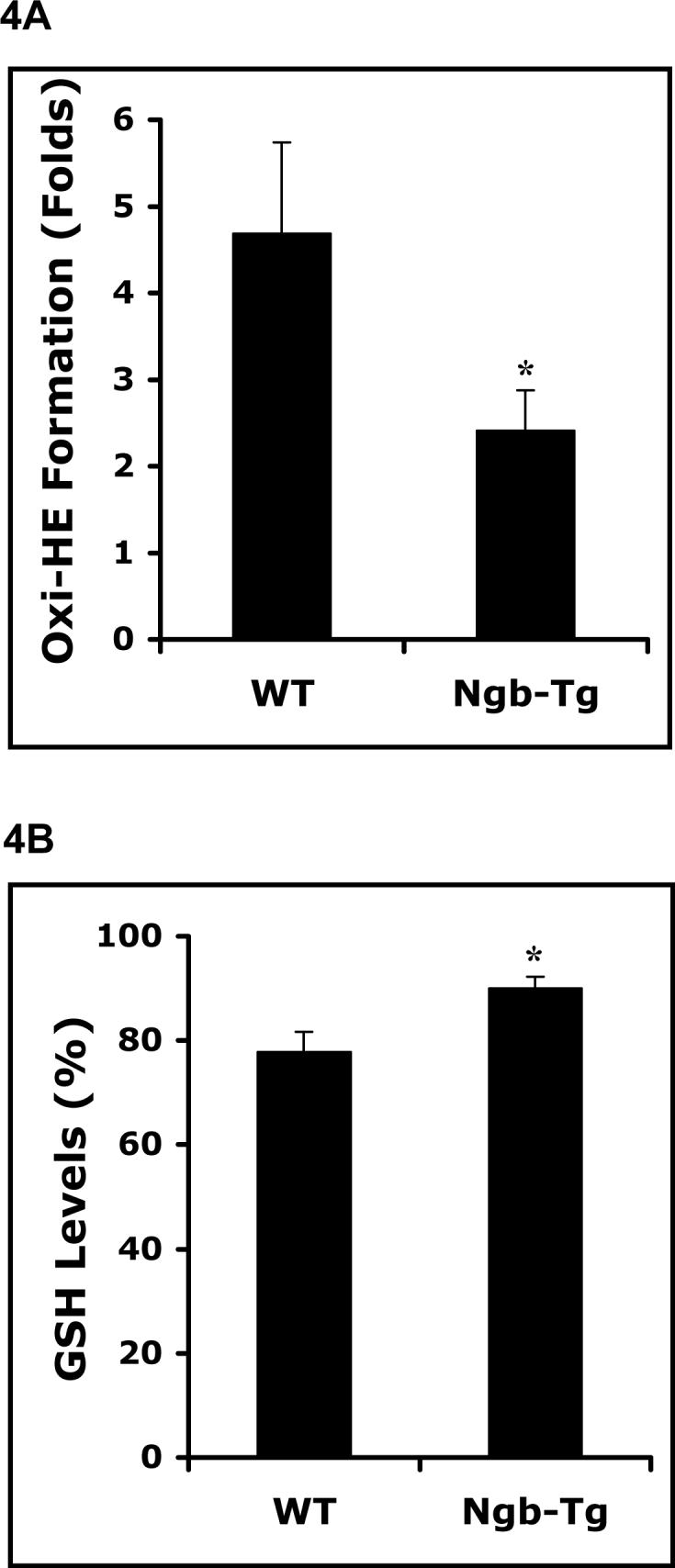
Reduction of oxidative stress in Ngb over-expressing neurons after H/R. A. Oxidized HE production was reduced in Ngb-Tg neurons after hypoxia for 10 hrs followed by 2 hr reoxygenation compared with WT neurons examined with fluorescent plate reader. B. Glutathione level of normal controls in Ngb-Tg neurons was significantly increased after hypoxia for 10 hrs and followed by 4 hrs reoxygenation compared with WT neurons. Mean±SEM, n=4 for each group. *P<0.05.
Discussion
In present study, energy failure in Ngb over-expressing neurons seemed to be prevented by an enhancement in ATP levels and maintenance of mitochondrial MTT formation. Concomitantly, there was augmented recovery of mitochondrial respiration which ultimately downregulated ROS formation. Superoxide anion formation was reduced and conversely, endogenous level of the anti-oxidant GSH was sustained. Altogether, these salutary effects may explain Ngb's ability to protect against hypoxia-induced neuron death.
However, insight of molecular mechanisms of Ngb's effects in mitochondrial function and oxidative stress after hypoxia/ischemia remains to be further investigated. There are some potential mechanisms that might be involved as discussed below. It has been suggested that Ngb may function as a NADH oxidase, which facilitate the glycolytic production of ATP under semi-anaerobic conditions (Sowa et al. 1998). This role of Ngb may help neurons sustain homeostasis, preserve mitochondrial function by preventing massive energetic imbalance. Ngb may also function as an O2 sensor, participating in a signal transduction pathway that modulates the activities of regulatory proteins in response to changes in O2 concentration. Indeed, it has been documented that large conformational changes occur in the heme pocket of hexacoordinated globins upon ligand binding, and which might very well trigger signals in hypothetical downstream regulators (Hargrove et al. 2000; Kriegl et al. 2002). Additionally, oxygen sensors associated with heme proteins may respond to hypoxia by altering the rate of formation and release of reactive oxygen species that activate transcription factors (Brunori et al. 2005; Bunn and Poyton 1996; Vanden Hoek et al. 1998). Interestingly, human ferric Ngb has been found to act as a guanine nucleotide dissociation inhibitor for the α subunit of heterotrimetric G proteins, it suggests that Ngb may function as a regulator of signal transduction in the brain and tightly correlated with its neuroprotective activity (Wakasugi et al. 2005; Watanabe and Wakasugi 2008). A most recent study showed Ngb over-expression eliminated hypoxia-induced mitochondrial aggregation and neuron death. This phenomenon was correlated to inhibition of PaK1 kinase activity and Rac1-GDP-dissociation inhibitor disassociation, inhibition of actin assembly and death-signaling module polarization (Khan et al. 2008). Furthermore, Ngb may function as a scavenger of nitric oxide (NO) or a regulator of reactive oxygen species (ROS) (Brunori et al. 2005; Garry and Mammen 2003; Herold et al. 2004). By altering these free radicals, Ngb may protect against oxidative stress in stroke. In our previous study, by using newly created Ngb overexpressing transgenic mice, we showed that Ngb is neuroprotective against focal cerebral ischemia in vivo (Wang et al. 2008), consistent with findings from the Greenberg group (Khan et al. 2006). Importantly, we also observed that reduced ROS biomarker malondialdehyde level found earlier at 8 hrs after transient focal cerebral ischemia in Ngb-Tg mice compared to WT mice, indicating Ngb might play roles for amelioration of oxidative damage in the development of ischemic brain infarction (Wang et al. 2008). A number of recent reports showed the enhanced survival of Ngb-transfected SH-SY5Y and PC12 cells after exposure to H(2)O(2), and Ngb-transfected HN33 cells treated with NO donor, all suggests that neuroglobin may protect against different types of oxidative damages (Fordel et al. 2006; Jin et al. 2008; Li et al. 2007). However, the insight of molecular interaction of Ngb in modulating mitochondrial function and oxidative stress after hypoxia/ischemia needs to be further elucidated.
There are a few caveats in this study. First, direct and quantitative measurements of mitochondrial function and oxidative stress are difficult to achieve. Our assays mostly comprise surrogate markers of mitochondrial function and oxidative stress. For example, MTT reduction is an indirect index of mitochondria viability, but changes in non-mitochondria metabolisms may exert nonspecific effects on the measurement (Caughlan et al. 2004; Cohen et al. 1997). Similarly, we do not have direct quantitation of actual levels of free radicals and reactive species in our model system. Our assays for oxi-HE and GSH levels only serve as biomarkers of oxidative and nitrosative injury, albeit ones that are widely used. A second caveat pertains to the modes of neuronal cell death that are involved. Our results from these H/R experiments indirectly suggest an important role for oxidative and mitochondrial stress. However, cell death occurs following ischemic/hypoxic insults can proceed with multiple phenotypes and mechanisms, including a complex mix of apoptosis, programmed necrosis, or even autophagy (Bredesen et al. 2006; Dawson and Dawson 2004). How Ngb interacts with the multiplicity of these overlapping cell death pathways remains to be determined. Finally, a major caveat in our study relates to causality. Our data here implicated that Ngb might affect both mitochondrial function and free radical generation as its potential neuroprotective mechanisms. But there are multiple and probably inextricable feedback loops between preservation of mitochondrial energetics versus direct radical scavenging (Chan 2001; Perez-Pinzon et al. 2005; Saito et al. 2005; Sims and Anderson 2002). We acknowledge that it will likely be impossible to unequivocally separate mitochondrial effects versus ROS effects of Ngb.
In summary, there was an early augmented recovery of mitochondrial respiration and concomitantly diminished biomarkers of ROS formation in Ngb over-expressing neurons after H/R insult. Altogether, these salutary effects may ultimately protect against hypoxia-induced late neuron death. The cell-based findings here suggest that the beneficial effects of Ngb are mediated in part by the preservation of mitochondrial function and amelioration of oxidative stress in neurons.
Acknowledgments
This work was supported in part by a NIH grant R01-NS049476 and a Scientist Development Grant 0435087N from American Heart Association (to X.W.), and NIH grants R01-NS48422, R01-NS53560, and P01-NS55104 (to E.H.L).
Grant supports: NIH grant R01-NS049476 and a Scientist Development Grant 0435087N from American Heart Association (to X.W.), and NIH grants R01-NS48422, R01-NS53560, and P01-NS55104 (to E.H.L).
Footnotes
Publisher's Disclaimer: This PDF receipt will only be used as the basis for generating PubMed Central (PMC) documents. PMC documents will be made available for review after conversion (approx. 2−3 weeks time). Any corrections that need to be made will be done at that time. No materials will be released to PMC without the approval of an author. Only the PMC documents will appear on PubMed Central -- this PDF Receipt will not appear on PubMed Central.
References
- Awenius C, Hankeln T, Burmester T. Neuroglobins from the zebrafish Danio rerio and the pufferfish Tetraodon nigroviridis. Biochem Biophys Res Commun. 2001;287(2):418–421. doi: 10.1006/bbrc.2001.5614. [DOI] [PubMed] [Google Scholar]
- Bentmann A, Schmidt M, Reuss S, Wolfrum U, Hankeln T, Burmester T. Divergent distribution in vascular and avascular mammalian retinae links neuroglobin to cellular respiration. J Biol Chem. 2005;280(21):20660–20665. doi: 10.1074/jbc.M501338200. [DOI] [PubMed] [Google Scholar]
- Bredesen DE, Rao RV, Mehlen P. Cell death in the nervous system. Nature. 2006;443(7113):796–802. doi: 10.1038/nature05293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brunori M, Giuffre A, Nienhaus K, Nienhaus GU, Scandurra FM, Vallone B. Neuroglobin, nitric oxide, and oxygen: functional pathways and conformational changes. Proc Natl Acad Sci U S A. 2005;102(24):8483–8488. doi: 10.1073/pnas.0408766102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bunn HF, Poyton RO. Oxygen sensing and molecular adaptation to hypoxia. Physiol Rev. 1996;76(3):839–885. doi: 10.1152/physrev.1996.76.3.839. [DOI] [PubMed] [Google Scholar]
- Burmester T, Gerlach F, Hankeln T. Regulation and role of neuroglobin and cytoglobin under hypoxia. Adv Exp Med Biol. 2007;618:169–180. doi: 10.1007/978-0-387-75434-5_13. [DOI] [PubMed] [Google Scholar]
- Burmester T, Hankeln T. Neuroglobin: a respiratory protein of the nervous system. News Physiol Sci. 2004;19:110–113. doi: 10.1152/nips.01513.2003. [DOI] [PubMed] [Google Scholar]
- Burmester T, Weich B, Reinhardt S, Hankeln T. A vertebrate globin expressed in the brain. Nature. 2000;407(6803):520–523. doi: 10.1038/35035093. [DOI] [PubMed] [Google Scholar]
- Caughlan A, Newhouse K, Namgung U, Xia Z. Chlorpyrifos induces apoptosis in rat cortical neurons that is regulated by a balance between p38 and ERK/JNK MAP kinases. Toxicol Sci. 2004;78(1):125–134. doi: 10.1093/toxsci/kfh038. [DOI] [PubMed] [Google Scholar]
- Chan PH. Reactive oxygen radicals in signaling and damage in the ischemic brain. J Cereb Blood Flow Metab. 2001;21(1):2–14. doi: 10.1097/00004647-200101000-00002. [DOI] [PubMed] [Google Scholar]
- Cohen G, Farooqui R, Kesler N. Parkinson disease: a new link between monoamine oxidase and mitochondrial electron flow. Proc Natl Acad Sci U S A. 1997;94(10):4890–4894. doi: 10.1073/pnas.94.10.4890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dawson VL, Dawson TM. Deadly conversations: nuclear-mitochondrial cross-talk. J Bioenerg Biomembr. 2004;36(4):287–294. doi: 10.1023/B:JOBB.0000041755.22613.8d. [DOI] [PubMed] [Google Scholar]
- Fordel E, Thijs L, Martinet W, Lenjou M, Laufs T, Van Bockstaele D, Moens L, Dewilde S. Neuroglobin and cytoglobin overexpression protects human SH-SY5Y neuroblastoma cells against oxidative stress-induced cell death. Neurosci Lett. 2006;410(2):146–151. doi: 10.1016/j.neulet.2006.09.027. [DOI] [PubMed] [Google Scholar]
- Garry DJ, Kanatous SB, Mammen PP. Emerging roles for myoglobin in the heart. Trends Cardiovasc Med. 2003;13(3):111–116. doi: 10.1016/s1050-1738(02)00256-6. [DOI] [PubMed] [Google Scholar]
- Garry DJ, Mammen PP. Neuroprotection and the role of neuroglobin. Lancet. 2003;362(9381):342–343. doi: 10.1016/S0140-6736(03)14055-X. [DOI] [PubMed] [Google Scholar]
- Geuens E, Brouns I, Flamez D, Dewilde S, Timmermans JP, Moens L. A globin in the nucleus! J Biol Chem. 2003;278(33):30417–30420. doi: 10.1074/jbc.C300203200. [DOI] [PubMed] [Google Scholar]
- Greenberg DA, Jin K, Khan AA. Neuroglobin: an endogenous neuroprotectant. Curr Opin Pharmacol. 2008;8(1):20–24. doi: 10.1016/j.coph.2007.09.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hargrove MS, Brucker EA, Stec B, Sarath G, Arredondo-Peter R, Klucas RV, Olson JS, Phillips GN., Jr Crystal structure of a nonsymbiotic plant hemoglobin. Structure Fold Des. 2000;8(9):1005–1014. doi: 10.1016/s0969-2126(00)00194-5. [DOI] [PubMed] [Google Scholar]
- Herold S, Fago A, Weber RE, Dewilde S, Moens L. Reactivity studies of the Fe(III) and Fe(II)NO forms of human neuroglobin reveal a potential role against oxidative stress. J Biol Chem. 2004;279(22):22841–22847. doi: 10.1074/jbc.M313732200. [DOI] [PubMed] [Google Scholar]
- Jin K, Mao XO, Xie L, Khan AA, Greenberg DA. Neuroglobin protects against nitric oxide toxicity. Neurosci Lett. 2008;430(2):135–137. doi: 10.1016/j.neulet.2007.10.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khan AA, Ou Mao X, Banwait S, Dermardirossian CM, Bokoch GM, Jin K, Greenberg DA. Regulation of hypoxic neuronal death signaling by neuroglobin. Faseb J. Jan 15, 2008. p. 18198211. 2008. [Epub ahead of print] PMID. [DOI] [PMC free article] [PubMed]
- Khan AA, Sun Y, Jin K, Mao XO, Chen S, Ellerby LM, Greenberg DA. A neuroglobin-overexpressing transgenic mouse. Gene. 2007;398(1−2):172–176. doi: 10.1016/j.gene.2007.02.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khan AA, Wang Y, Sun Y, Mao XO, Xie L, Miles E, Graboski J, Chen S, Ellerby LM, Jin K, Greenberg DA. Neuroglobin-overexpressing transgenic mice are resistant to cerebral and myocardial ischemia. Proc Natl Acad Sci U S A. 2006;103(47):17944–17948. doi: 10.1073/pnas.0607497103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kriegl JM, Bhattacharyya AJ, Nienhaus K, Deng P, Minkow O, Nienhaus GU. Ligand binding and protein dynamics in neuroglobin. Proc Natl Acad Sci U S A. 2002;99(12):7992–7997. doi: 10.1073/pnas.082244399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee SR, Lo EH. Interactions between p38 mitogen-activated protein kinase and caspase-3 in cerebral endothelial cell death after hypoxia-reoxygenation. Stroke. 2003;34(11):2704–2709. doi: 10.1161/01.STR.0000096540.40826.BA. [DOI] [PubMed] [Google Scholar]
- Li RC, Morris MW, Lee SK, Pouranfar F, Wang Y, Gozal D. Neuroglobin protects PC12 cells against oxidative stress. Brain Res. 2008;1190:159–166. doi: 10.1016/j.brainres.2007.11.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mammen PP, Shelton JM, Goetsch SC, Williams SC, Richardson JA, Garry MG, Garry DJ. Neuroglobin, a novel member of the globin family, is expressed in focal regions of the brain. J Histochem Cytochem. 2002;50(12):1591–1598. doi: 10.1177/002215540205001203. [DOI] [PubMed] [Google Scholar]
- Marcinek DJ, Ciesielski WA, Conley KE, Schenkman KA. Oxygen regulation and limitation to cellular respiration in mouse skeletal muscle in vivo. Am J Physiol Heart Circ Physiol. 2003;285(5):H1900–908. doi: 10.1152/ajpheart.00192.2003. [DOI] [PubMed] [Google Scholar]
- Nicholls DG, Budd SL. Mitochondria and neuronal survival. Physiol Rev. 2000;80(1):315–360. doi: 10.1152/physrev.2000.80.1.315. [DOI] [PubMed] [Google Scholar]
- Perez-Pinzon MA, Dave KR, Raval AP. Role of reactive oxygen species and protein kinase C in ischemic tolerance in the brain. Antioxid Redox Signal. 2005;7(9−10):1150–1157. doi: 10.1089/ars.2005.7.1150. [DOI] [PubMed] [Google Scholar]
- Saito A, Maier CM, Narasimhan P, Nishi T, Song YS, Yu F, Liu J, Lee YS, Nito C, Kamada H, Dodd RL, Hsieh LB, Hassid B, Kim EE, Gonzalez M, Chan PH. Oxidative stress and neuronal death/survival signaling in cerebral ischemia. Mol Neurobiol. 2005;31(1−3):105–116. doi: 10.1385/MN:31:1-3:105. [DOI] [PubMed] [Google Scholar]
- Sims NR, Anderson MF. Mitochondrial contributions to tissue damage in stroke. Neurochem Int. 2002;40(6):511–526. doi: 10.1016/s0197-0186(01)00122-x. [DOI] [PubMed] [Google Scholar]
- Sowa AW, Duff SMG, Guy PA, Hill RD. Altering hemoglobin levels changes energy status in maize cells under hypoxia. Proc Natl Acad Sci U S A. 1998;95(17):10317–10321. doi: 10.1073/pnas.95.17.10317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun Y, Jin K, Mao XO, Zhu Y, Greenberg DA. Neuroglobin is up-regulated by and protects neurons from hypoxic-ischemic injury. Proc Natl Acad Sci U S A. 2001;98(26):15306–15311. doi: 10.1073/pnas.251466698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vanden Hoek TL, Becker LB, Shao Z, Li C, Schumacker PT. Reactive oxygen species released from mitochondria during brief hypoxia induce preconditioning in cardiomyocytes. J Biol Chem. 1998;273(29):18092–18098. doi: 10.1074/jbc.273.29.18092. [DOI] [PubMed] [Google Scholar]
- Flögel U, Gödecke A, Klotz LO, Schrader J. Role of myoglobin in the antioxidant defense of the heart. FASEB. 2004;18:1156–1158. doi: 10.1096/fj.03-1382fje. [DOI] [PubMed] [Google Scholar]
- Wakasugi K, Kitatsuji C, Morishima I. Possible neuroprotective mechanism of human neuroglobin. Ann N Y Acad Sci. 2005;1053:220–230. doi: 10.1196/annals.1344.020. [DOI] [PubMed] [Google Scholar]
- Wang X, Liu J, Zhu H, Tejima E, Tsuji K, Murata Y, Atochin DN, Huang PL, Zhang C, Lo EH. Effects of neuroglobin over-expression on acute brain injury and long-term outcomes after focal cerebral ischemia. Stroke. Apr 10, 2008. p. 18403737. [Epub ahead of print] PMID. [DOI] [PMC free article] [PubMed]
- Wang X, Mori T, Sumii T, Lo EH. Hemoglobin-induced cytotoxicity in rat cerebral cortical neurons: caspase activation and oxidative stress. Stroke. 2002;33(7):1882–1888. doi: 10.1161/01.str.0000020121.41527.5d. [DOI] [PubMed] [Google Scholar]
- Watanabe S, Wakasugi K. Neuroprotective function of human neuroglobin is correlated with its guanine nucleotide dissociation inhibitor activity. Biochem Biophys Res Commun. 2008;369(2):695–700. doi: 10.1016/j.bbrc.2008.02.089. [DOI] [PubMed] [Google Scholar]
- Zhang CG, Li L, Deng MY, Xie F, Wang CL, Zhou WQ, Wang HY, He FC. [Coding region cDNA sequence cloning of rat neuroglobin gene, its polymorphism feature and tissue expression profile analysis]. Yi Chuan Xue Bao. 2001;28(11):997–1001. [PubMed] [Google Scholar]