

Abstract
Unconscious motivation in humans is often inferred but rarely demonstrated empirically. We imaged motivational processes, implemented in a paradigm that varied the amount and reportability of monetary rewards for which subjects exerted physical effort. We show that, even when subjects cannot report how much money is at stake, they nevertheless deploy more force for higher amounts. Such a motivational effect is underpinned by engagement of a specific basal forebrain region. Our findings thus reveal this region as a key node in brain circuitry that enables expected rewards to energize behavior, without the need for the subjects’ awareness.
Humans tend to adapt the degree of effort they expend according to the magnitude of reward they expect. Such a process has been proposed as an operant concept of motivation (1-3). Motivational processes may be obvious, as when a prospector spends days in extreme conditions seeking gold. The popular view is that motivation can also be unconscious, such that a person may be unable to report the goals or rewards that drive a particular behavior. However, empirical evidence on this issue is lacking, and the potential brain mechanisms involved in converting expected rewards into behavioral activation are poorly understood.
We developed an experimental paradigm to visualize unconscious motivational processes, using functional magnetic resonance imaging. A classical approach to trigger unconscious processing is subliminal stimulation, which can be implemented by means of masking procedures. The terminology we use in this report is based on a recent taxonomy (4), in which a process is considered subliminal if it is attended but not reportable. Successful brain imaging studies of subliminal processes have focused so far on processing words (5, 6) as well as emotional stimuli (7, 8). In our study, the object of masking was an incentive stimulus for a future action, represented by the amount of reward at stake. The question we asked is whether, and how, the human brain energizes behavior in proportion to subliminal incentives.
We developed an incentive force task, using money as a reward: a manipulation that is consistently shown to activate reward circuits in the human brain (9-11). The exact level of motivation was manipulated by randomly assigning the amount at stake as one pound or one penny. Pictures of the corresponding coins were displayed on a computer screen at the beginning of each trial, between two screenshots of “mask” images (Fig. 1). The reportabiity of the monetary stakes depended on their display duration, which could be 17,50, or 100ms. The perception of the first two durations was determined as subliminal in a preliminary behavioral test, where subjects reported not seeing anything other than the mask. The third duration was consistently associated with conscious perception of the stimuli and their associated amount.
Fig. 1.

The incentive force task. Successive screens displayed in one trial are shown from left to right, with durations in ms. Coin images, either one pound (£1) or one penny (1p), indicate the monetary value attributed to the top of the thermometer image. The fluid level in the thermometer represents the online force exerted on the hand grip. The last screen indicates cumulative total of the money won so far.
To characterize the effects of the monetary stakes, we recorded not only brain activity but also skin conductance and hand-grip force. Skin conductance response (SCR) is linked to autonomic sympathetic arousal (12) and is thereafter interpreted as reflecting an affective evaluation of the monetary stake. Hand-grip force is understood to be a measure of behavioral activation. Online visual feedback of the force exerted was displayed as a fluid level moving up and down within a thermometer depicted on the screen (Fig. 1). Subjects were instructed that the higher the fluid level rose, the more of the monetary stake they would get to keep. At the end of the trial, subjects were given visual feedback of the amount of money that they had accumulated. Thus, this cumulative total was increased after every trial, though negligibly so when one penny was at stake.
For the analysis of brain activity, we first examined the main contrast between monetary stakes, in the conscious condition, at the time of stimulus onset (Fig. 2, middle column). After correction for multiple comparisons over the whole brain (family-wise error, P < 0.05), the only significant activation was located bilaterally in the basal forebrain, bordering several structures encompassing the ventral striatum, ventral pallidum (VP), extended amygdala, and basal nucleus of Meynert. These structures have been conceptualized as forming output channels for the limbic system, which is devoted to emotional and motivational functions (13). According to fiber tracing studies, reward-related information may access these structures either by a subcortical route via the hippocampus and/or amygdala or by a cortical route via the orbitofrontal and/or anterior cingulate areas (14-17).
Fig. 2.
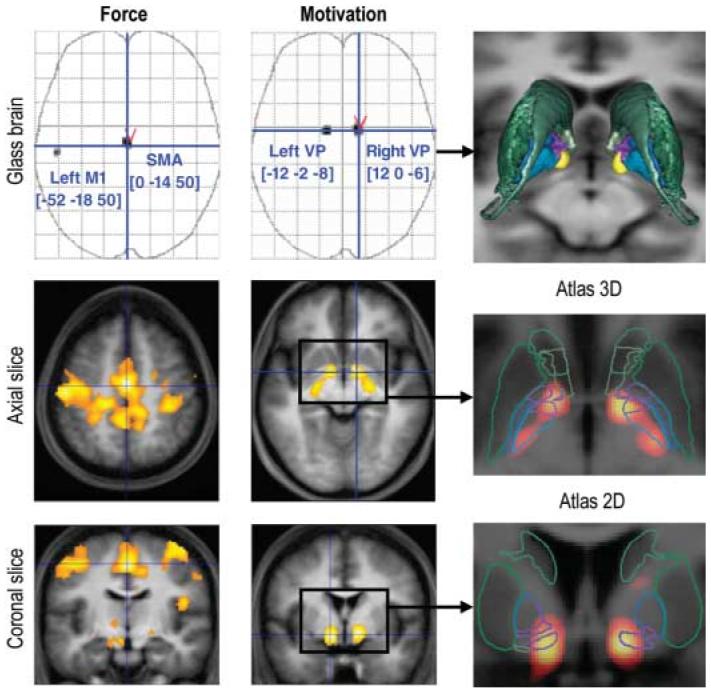
SPMs of brain activity. Voxels displayed in gray on glass brains showed a significant effect at P < 0.05 after correction for multiple comparisons over the entire brain. The [x, y, z] coordinates of the different maxima refer to the Montreal Neurological Institute (MNI) space. Axial and coronal slices were taken at global maxima of interest indicated by red symbols on the glass brains. SPMs are shown at a lower threshold (P < 0.001, uncorrected) and were superimposed on the average structural scan to localize significant activations. The images in the left column show regression with the amount of force produced, whatever the condition. The images in the middle column show contrast between conscious pounds and pennies trials (£1 to 1p, 100 ms). For this contrast, SPMs were coregistered with an atlas of the basal ganglia (right column). Caudate, putamen, and accumbens are shown in green; external and internal pallidum are shown in blue, with limbic sectors in violet.
To improve anatomical localization, we coregistered the statistical parametric map (SPM) with a recent histology-based atlas of the basal ganglia, which was designed to distinguish between functional territories (18, 19). Activation foci overlapped with limbic territories of both external and internal pallidal segments (Fig. 2, right column), which together form the VP. The main inputs to the VP come from the ventral striatum, where reward-related activations have been consistently found (9-11). VP activation might denote engagement of the same ventral striato-pallidal pathway, with a shift in its expression being related to the nature of the upcoming task. More specifically, ventral striatal activity has been linked to reward prediction and reward prediction error during learning (20, 21). Rather than concentrating on learning, our design focused on motivation during effort, which elicited specific processing in the VP. Our finding accords well with evidence in rodents, showing that VP neurons encode rewarding properties of environmental stimuli (22), and suggests a role for the VP in incentive motivation. Furthermore, lowering the threshold (P < 0.00 1, uncorrected) revealed that activation extended posteriorly, within nonlimbic territories ofthe pallidum, pointing out a plausible route by which the VP may influence cortical motor areas (14, 15).
To dissociate motivation per se from force production, we next examined brain activity that was linearly related to the amount of force produced, whatever the condition (Fig. 2, left column). After correction for multiple comparisons over the whole brain (family-wise error, P<0.05), significant activations were found in the supplementary motor area (SMA) and in the primary motor area (M1). Unlike the pallidum, these structures have previously been shown to activate in relation with the amount of force produced (23-25). Moreover, M1 activation was observed on the left side, which was consistent with the use of the right hand for the task, whereas pallidal activation was bilateral. Thus, in our analysis, the dissociation was clear-cut, probably reflecting the fact that monetary stakes were constant throughout the task, while grip force decreased trial after trial, probably as a result of fatigue (fig. S1). Such dissociation suggests that motivational processes mediated by the VP include modulation of SMA activity, which in turn drives muscular contractions via M1.
We next asked whether such a circuit was engaged by subliminal incentives. We averaged parameter estimates (Fig. 3A, left panels) over the pallidal voxels that showed significant activation in the previous SPM. The contrast between monetary stakes was significant for 100 and 50 ms (paired t tests, both P values < 0.001) but not for 17 ms. No significant activation was found elsewhere in an SPM estimated for this contrast at 50 ms, even with our liberal threshold (P < 0.001, uncorrected). Thus, only the VP appeared in position to modulate behavioral activation according to subliminal incentives and hence to underpin a low-level motivational process, as opposed to a conscious cost-benefit calculation. Again, such a role accords well with experiments on rodents, which show that VP manipulations influence goal-directed behavior, as seen with self-stimulation after electrode implantation in the VP (26) or impaired acquisition of conditioned-place preference after the generation of VP lesions (27).
Fig. 3.
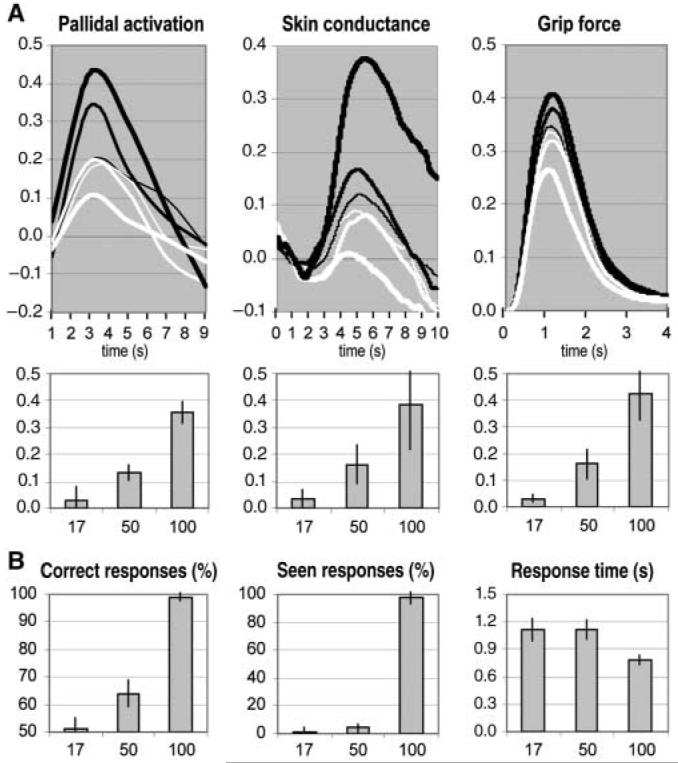
Main effects of stimulus duration. (A) Incentive force task. Time courses were averaged across trials for the different stimuli (black lines indicate £1 and white lines indicate 1p) and durations (thin, intermediate, and thick lines indicate 17, 50, and 100 ms, respectively). Time 0 corresponds to the moment of stimulus display. The histograms indicate the effect of motivation (£1 to 1p), and the error bars indicate SEM. Pallidal activation is expressed as percentage of blood oxygen level-dependent signal change. Force and skin conductance are expressed in proportion of the highest measure. (B) Perception task. Stimuli were the same as in (A). Possible responses were “seen £1,” “seen 1p,” “guess £1,” and “guess 1p.” A “correct” response means that the subject chose the stimulus that had been displayed. A “seen” response means that the subject perceived all or part of the stimulus. Error bars indicate SEM.
We next sought to link our imaging results to simultaneously measured autonomic and behavioral responses. The dynamics of responses recorded from skin conductance electrodes indicated that they were triggered at the time of stimulus display, with a typical SCR profile starting at 2 s post-stimulus and peaking around 5 s (12). Comparison between monetary stakes showed significant effects at 100 and 50 ms (paired ttests, both P values < 0.05) but not at 17 ms (Fig. 3A, middle panels). Thus, like fear-relevant stimuli (28), subliminal incentives could be evaluated affectively, with subjects being more responsive to images of pounds than to those of pennies. Autonomic responding was not a mere side effect of force production, because it evolved with a different temporal profile throughout the task. Indeed, grip force decreased for consciously perceived pennies, while skin conductance increased for consciously perceived pounds (fig. S1). Regarding grip force, we found similar dynamics, whatever the condition: subjects giving a short squeeze, with peak latency at around 1 s, and relaxing before the next trial (Fig. 3A, right panels). Hence, similar results were found when considering either the height of the peak or the area under the curve. Comparing between monetary stakes, significant effects were found at 100 ms, 50 ms, and even at 17 ms (paired ttests, all P values < 0.01). Thus, the brain could energize behavior in proportion to the reward at stake, even when subjects could not see it.
Finally, we controlled for subjective perception with a forced choice task (Fig. 3B). While still in the scanner, subjects were shown the same masked stimuli and had to report whether they saw a coin, and which coin they thought it was, either from seeing it or from guessing. Based on the percentage of correct responses, the analysis could then be restricted to all situations where subjects guess at chance level about stimulus identity (fig. S2). Even in these situations, pallidal activation and hand-grip force were significantly higher for pounds as compared to pennies (paired t tests, both P values <0.01). As with the preliminary test, subjects reported seeing almost no stimuli at 17 and 50 ms and almost all stimuli at 100 ms. Compared to the 100-ms condition, subjects also had similarly long response times at 17 and 50 ms, indicating that they were experiencing the same degree of uncertainty about stimulus identity. Thus, subjective perception changed as a function of category, from subliminal to conscious, between 50 and 100 ms. In contrast, objective markers of motivation (pallidal activation, SCR, and hand-grip force) gradually increased with stimulus duration.
These results indicate that motivational processes involved in boosting behavior are qualitatively similar, despite whether subjects are conscious or not of the reward at stake. Consistently, the same basal forebrain region underpinned subliminal and conscious motivation. Such subcortical localization might relate to the simple and repetitive nature of the task, rendering strategic control unnecessary. However, differential sympathetic arousal denoted by SCRs argues against an interpretation in terms of mere stimulus response habit formation, which is known to involve the basal ganglia (29). More generally, our paradigm offers a potential tool to discriminate between motor and affective components of motivation for financial reward in humans, analogous to the dissociation between wanting and liking food reward described in rodents (30). Such a tool may be particularly useful in exploring negative symptoms, like those manifested in depression or schizophrenia, involving acute dysfunction within the motivational process.
Supplementary Material
Supporting Online Material www.sciencemag.org/cgi/content/full/1140459/DC1
Acknowledgments
We thank E. Featherstone for skilful assistance in recording hand-grip data; J. Yelnik and E. Bardinet for having deformed their atlas on SPMs; and G. Lafargue, S. Kloppel, and C. Sergent for helpful discussions. This work was funded by the Wellcome Trust research program grants to C.F. and R.D. M.P. received a grant from the Fyssen Foundation.
References and Notes
- 1.Robbins TW, Everitt BJ. Curr. Opin. Neurobiol. 1996;6:228. doi: 10.1016/s0959-4388(96)80077-8. [DOI] [PubMed] [Google Scholar]
- 2.Berridge KC. Physiol. Behav. 2004;81:179. doi: 10.1016/j.physbeh.2004.02.004. [DOI] [PubMed] [Google Scholar]
- 3.Schultz W. Annu. Rev. Psychol. 2006;57:87. doi: 10.1146/annurev.psych.56.091103.070229. [DOI] [PubMed] [Google Scholar]
- 4.Dehaene S, et al. Trends Cognit. Sci. 2006;10:204. doi: 10.1016/j.tics.2006.03.007. [DOI] [PubMed] [Google Scholar]
- 5.Dehaene S, et al. Nature. 1998;395:597. doi: 10.1038/26967. [DOI] [PubMed] [Google Scholar]
- 6.Gaillard R, et al. Neuron. 2006;50:191. doi: 10.1016/j.neuron.2006.03.031. [DOI] [PubMed] [Google Scholar]
- 7.Morris JS, Ohman A, Dolan RJ. Nature. 1998;393:467. doi: 10.1038/30976. [DOI] [PubMed] [Google Scholar]
- 8.Etkin A, et al. Neuron. 2004;44:1043. doi: 10.1016/j.neuron.2004.12.006. [DOI] [PubMed] [Google Scholar]
- 9.O’Doherty J, et al. Nat. Neurosci. 2001;4:95. doi: 10.1038/82959. [DOI] [PubMed] [Google Scholar]
- 10.Elliott R, et al. Neurosci. 2003;23:303. [Google Scholar]
- 11.Knutson B, et al. Neurosci. 2005;25:4806. doi: 10.1523/JNEUROSCI.0642-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Critchley HD. Neuroscientist. 2002;8:132. doi: 10.1177/107385840200800209. [DOI] [PubMed] [Google Scholar]
- 13.Heimer L, Van Hoesen GW. Neurosci. Biobehav. Rev. 2006;30:126. doi: 10.1016/j.neubiorev.2005.06.006. [DOI] [PubMed] [Google Scholar]
- 14.Alexander GE, DeLong MR, Strick PL. Annu. Rev. Neurosci. 1986;9:357. doi: 10.1146/annurev.ne.09.030186.002041. [DOI] [PubMed] [Google Scholar]
- 15.Haber SN. Chem. Neuroanat. 2003;26:317. doi: 10.1016/j.jchemneu.2003.10.003. [DOI] [PubMed] [Google Scholar]
- 16.Lehericy S, et al. Ann. Neurol. 2004;55:522. doi: 10.1002/ana.20030. [DOI] [PubMed] [Google Scholar]
- 17.Friedman DP, Aggleton JP, Saunders RC. Comp. Neurol. 2002;450:345. doi: 10.1002/cne.10336. [DOI] [PubMed] [Google Scholar]
- 18.Yelnik J, et al. Neuroimage. 2007;34:618. doi: 10.1016/j.neuroimage.2006.09.026. [DOI] [PubMed] [Google Scholar]
- 19.Lehericy S, et al. Cereb. Cortex. 2006;16:149. doi: 10.1093/cercor/bhi089. [DOI] [PubMed] [Google Scholar]
- 20.O’Doherty J, et al. Science. 2004;304:452. doi: 10.1126/science.1094285. [DOI] [PubMed] [Google Scholar]
- 21.Pessiglione M, et al. Nature. 2006;442:1042. doi: 10.1038/nature05051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Tindell AJ, Berridge KC, Aldridge JW. Neurosci. 2004;24:1058. doi: 10.1523/JNEUROSCI.1437-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Dettmers C, et al. Neurophysiol. 1995;74:802. doi: 10.1152/jn.1995.74.2.802. [DOI] [PubMed] [Google Scholar]
- 24.Dai TH, et al. Exp. Brain Res. 2001;140:290. doi: 10.1007/s002210100815. [DOI] [PubMed] [Google Scholar]
- 25.Vaillancourt DE, et al. Neuroimage. 2004;23:175. doi: 10.1016/j.neuroimage.2004.04.040. [DOI] [PubMed] [Google Scholar]
- 26.Panagis G, et al. Behav. Brain Res. 1995;68:165. doi: 10.1016/0166-4328(94)00169-g. [DOI] [PubMed] [Google Scholar]
- 27.McAlonan GM, Robbins TW, Everitt BJ. Neuroscience. 1993;52:605. doi: 10.1016/0306-4522(93)90410-h. [DOI] [PubMed] [Google Scholar]
- 28.Ohman A, Mineka S. Psychol. Rev. 2001;108:483. doi: 10.1037/0033-295x.108.3.483. [DOI] [PubMed] [Google Scholar]
- 29.Graybiel AM. Curr. Opin. Neurobiol. 2005;15:638. doi: 10.1016/j.conb.2005.10.006. [DOI] [PubMed] [Google Scholar]
- 30.Berridge KC. Neurosci. Biobehav. Rev. 1996;20:1. doi: 10.1016/0149-7634(95)00033-b. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supporting Online Material www.sciencemag.org/cgi/content/full/1140459/DC1