

Abstract
CLAVATA pathway is one of best-characterized signaling pathway involves in the regulation of meristem development in Arabidopsis. Increasing evidence indicated that this pathway also exist in the monocots as well as in the dicots. We have recently identified FON4 in rice as an ortholog of CLV3 in Arabidopsis. FON4 is putative ligand of FON1, which play a role in restricting the meristem size in rice. FON4 and CLV3 are the members of CLE gene family, which encode small functional secreted peptide with a conserved 14-amino acid motif (CLE motif) near or at the C termini.
Key Words: FON4, CLAVATA, CLE, meristem, floral organ number
The meristems in angiosperms have the extraordinary capacity to produce new lateral organs, however, the stem cell, a small group undifferentiated cells in the central area of meristem have the ability to grow and divide to replace cells consumed in organ initiations. The balance of meristem maintenances and differentiations in plants is precisely regulated by some cell-to-cell communications.1
A putative peptide ligand, CLVATA3 (CLV3), of Arabidopsis thaliana interacts with a disulphide-linked CLV1/CLV2 receptor complex to restrict the stem cell population in a appropriate size in shoot apical meristem (SAM).2 Mutations in any of the three CLV locus result in over-proliferation of meristem.3–5 By contrast, a homeodomain transcription factor gene, WUSCHEL(WUS), is a positive regulator in maintaining the meristem, and the wus mutant has the opposite phenotype of clv.6 And the WUS expression was downregulated by the CLV3,7 and the CLV3 was upregulated by the WUS.8 This feedback loop plays a central role in insuring the balance of SAM maintenances and differentiations.
Some evidence indicated that a CLAVATA-like pathway for regulating SAM size is functionally conserved in monocots as well as in dicots. Two maize genes, fea2 and td1, encoded the putative orthologs of CLV2 and CLV1, respectively, and play the roles to restrict the meristem size during maize development.9,10 Further evidence comes from the identification and characterization of FON1 in rice, which is the putative ortholog of CLV1. Mutation in FON1 caused enlargement floral meristem and increase floral organ numbers.11 But the ligand participating in this pathway remains unknown in monocots.
We have recently reported the characterization of the rice mutant floral organ number 4 (fon4). The fon4 mutants produced abnormal enlargement of the SAM, inflorescence meristem and floral meristem, and resulted in thick culms and increase of both primary rachis branches and floral organs.12 The defect of floral development of fon4 is very similar to that of fon1 suggest that FON1 and FON4 may participate in same pathway.11 Using map-based cloning approach, we have identified the FON4 gene. The FON4 encoded a putative secreted peptide as an ortholog of CLV3. In Arabidopsis, CLV3 is predicted to act as a ligand for the CLV1 receptor kinase.2 Therefore, we speculated that FON4 may play as a ligand for FON1 in the regulation of meristem development in rice. Although FON1 is expressed in all meristems responsible for development of the aerial part of rice, the vegetative and inflorescence meristem almost normal in fon1 mutant,11 suggested that FON1 is probably not the only receptor of FON4. We treated fon1 mutant with 50µm a synthetic 14-amino acid peptide, FON4p, corresponding to the predicted CLE (CLV3/ESR-related) motif of FON4. After 15 days treatment, significant inhibition of apical growth was observed in the fon1 mutant (Fig. 1), this implies that other receptor(s) of FON4, but not FON1, also exist(s) in rice regulating SAM.
Figure 1.
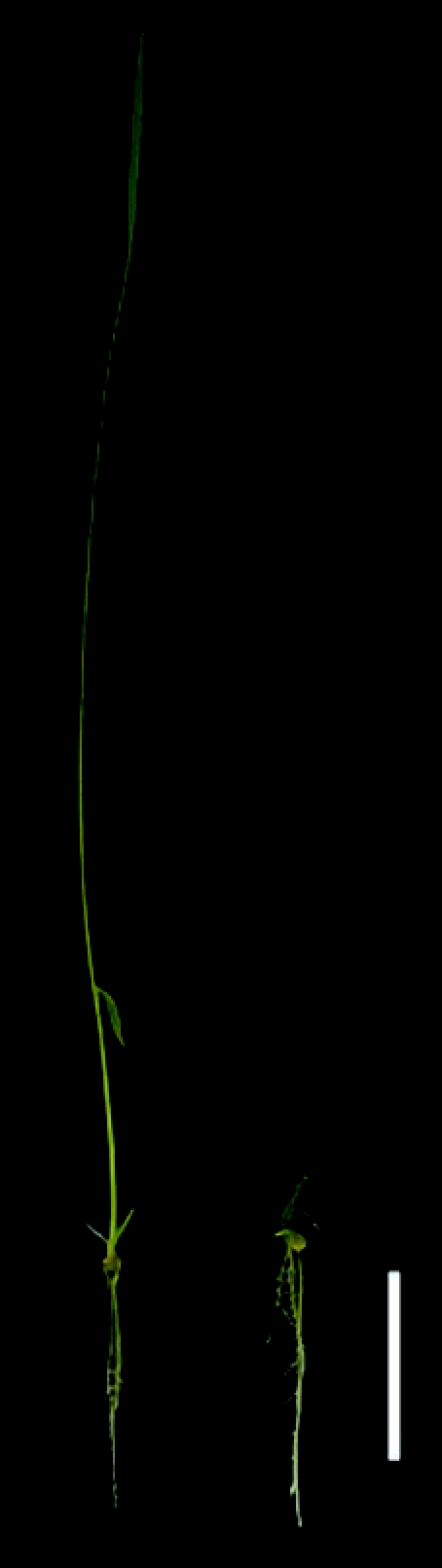
Treatment of the fon1 mutant with 50 µm FON4p (right), left is the the fon1 mutant treated with no FON4p as a control. Bar represents 2 cm.
In Arabidopsis, a process known as ligand sequestration limits the range of CLV3 diffusion, and ensures the WUS expressed in organizing center.1 Overexpression of CLV3 or treatment of synthetic 14-amino acid peptide, CLV3p, corresponding to the CLE motif of CLV3 in vitro can consume the SAM and reduce the WUS expression.7,13 This indicates that this ligand sequestration process is important for maintenance of meristems. FON4 is expressed at the apex of meristems, whereas the FON1 is expressed throughout the whole meristem,11 so we speculate that the similar ligand sequestration process of FON4 and FON1 occurs in rice. In agreement with this, treatment with FON4p also resulted in the consumption of rice SAM. Furthermore, we speculated that the rice SAM also has an organizing center as well as in Arabidopsis, where the FON4 can not reach.
In vitro application CLV3p causes the consumption of the root meristem and decrease the SAM size in Arabidopsis.13,14 Similarly, treatment of CLV3p also caused consumption of root meristem and SAM in rice as well as in Arabidopsis. However, the treatment of FON4p only caused the consumption of SAM, and no obvious defects were observed in root meristem. This suggests that a CLV-like pathway is conserved in regulating rice root meristem, and different from CLV3, FON4 does not participate in limiting rice root meristem.
Acknowledgements
We thank for Dr. Yasuo Nagato for providing fon1 mutant. This work was supported by the Funds from the National Key Basic Research Developments Program of the Ministry of Science and Technology, People's Republic of China (2006CB101700 and 2005CB120802), the National ‘863” High-Tech Project (2005AA2710330), the Program for New Century Excellent Talents in University (NCET-04-0403), the Shuguang Scholarship (04SG15), and the Shanghai Institutes of Biological Sciences (Reproductive Development Project).
Footnotes
Previously published online as a Plant Signaling & Behavior E-publication: http://www.landesbioscience.com/journals/psb/abstract.php?id=3641
References
- 1.Clark SE. Meristems: Start your signaling. Curr Opin Plant Biol. 2001;4:28–32. doi: 10.1016/s1369-5266(00)00131-x. [DOI] [PubMed] [Google Scholar]
- 2.Fletcher JC, Brand U, Running MP, Simon R, Meyerowitz EM. Signaling of cell fate decisions by CLAVATA3 in Arabidopsis shoot meristems. Science. 1999;283:1911–1914. doi: 10.1126/science.283.5409.1911. [DOI] [PubMed] [Google Scholar]
- 3.Clark SE, Running MP, Meyerowitz EM. CLAVATA1, a regulator of meristem and flower development in Arabidopsis. Development. 1993;119:397–418. doi: 10.1242/dev.119.2.397. [DOI] [PubMed] [Google Scholar]
- 4.Clark SE, Running MP, Meyerowitz EM. CLAVATA3 is a specific regulator of shoot and floral meristem development affecting the same processes as CLAVATA1. Development. 1995;121:2057–2067. [Google Scholar]
- 5.Kayes JM, Clark SE. CLAVATA2: A regulator of meristem and organ development in Arabidopsis. Development. 1998;125:3843–3851. doi: 10.1242/dev.125.19.3843. [DOI] [PubMed] [Google Scholar]
- 6.Mayer KF, Schoof H, Haecker A, Lenhard M, Jurgens G, Laux T. Role of WUSCHEL in regulating stem cell fate in the Arabidopsis shoot meristem. Cell. 1998;95:805–815. doi: 10.1016/s0092-8674(00)81703-1. [DOI] [PubMed] [Google Scholar]
- 7.Brand U, Fletcher JC, Hobe M, Meyerowitz EM, Simon R. Dependence of stem cell fate in Arabidopsis on a feedback loop regulated by CLV3 activity. Science. 2000;289:617–619. doi: 10.1126/science.289.5479.617. [DOI] [PubMed] [Google Scholar]
- 8.Schoof H, Lenhard M, Haecker A, Mayer KF, Jürgens G, Laux T. The stem cell population of Arabidopsis shoot meristems is maintained by a regulatory loop between the CLAVATA and WUSCHEL genes. Cell. 2000;100:635–644. doi: 10.1016/s0092-8674(00)80700-x. [DOI] [PubMed] [Google Scholar]
- 9.Taguchi-Shiobara F, Yuan Z, Hake S, Jackson D. The fasciated ear2 gene encodes a leucine-rich repeat receptor-like protein that regulates shoot meristem proliferation in maize. Genes Dev. 2001;15:2755–2766. doi: 10.1101/gad.208501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Bommert P, Lunde C, Nardmann J, Vollbrecht E, Running M, Jackson D, Hake S, Werr S. Thick tassel dwarf1 encodes a putative maize ortholog of the Arabidopsis CLAVATA1 leucine-rich repeat receptorlike kinase. Development. 2005;132:1235–1245. doi: 10.1242/dev.01671. [DOI] [PubMed] [Google Scholar]
- 11.Suzaki T, Sato M, Ashikari M, Miyoshi M, Nagato Y, Hirano HY. The gene FLORAL ORGAN NUMBER1 regulates floral meristem size in rice and encodes a leucine-rich repeat receptor kinase orthologous to Arabidopsis CLAVATA1. Development. 2004;131:5649–5657. doi: 10.1242/dev.01441. [DOI] [PubMed] [Google Scholar]
- 12.Chu HW, Qian Q, Liang WQ, Yin CS, Tan HX, Yao X, Yuan Z, Yang J, Huang H, Luo D, Ma H, Zhang DB. The FLORAL ORGAN NUMBER4 gene encoding a putative ortholog of Arabidopsis CLAVATA3 regulates apical meristem size in rice. Plant Physiol. 2006;142:1039–1052. doi: 10.1104/pp.106.086736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Fiers M, Golemiec E, van der Schors R, van der Geest L, Li KW, Stiekema WJ, Liu CM. The CLV3/ESR motif of CLV3 is functionally independent from the nonconserved flanking sequences. Plant Physiol. 2006;141:1284–1292. doi: 10.1104/pp.106.080671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Fiers M, Golemiec E, Xu J, van der Geest L, Heidstra R, Stiekema W, Liu CM. The 14-amino acid CLV3, CLE19, and CLE40 peptides trigger consumption of the root meristem in Arabidopsis through a CLAVATA2-dependent pathway. Plant Cell. 2005;17:2542–2553. doi: 10.1105/tpc.105.034009. [DOI] [PMC free article] [PubMed] [Google Scholar]