

Abstract
A theoretical analysis is given for the rate of change of domain sizes in lipid monolayers at the air–water interface. The calculation is applicable to liquid domains formed from binary mixtures of lipids that form two coexisting liquid phases. Under conditions where the two lipid molecules have approximately equal areas, the equilibration rate does not involve macroscopic hydrodynamic flow in the subphase but rather depends on the diffusion coefficient of the lipid molecules. The calculation shows that the equilibration rate in binary mixtures of cholesterol and phosphatidylcholine is remarkably slow, the radius of a typical 20-μm diameter domain changing by as little as a part in a million per second. Under these circumstances, equilibration times of the order of days or weeks are expected. Even with such long times, the final state reached by the monolayer will in general be a state of metastable equilibrium, rather than true equilibrium.
Keywords: molecular diffusion, phase equilibria, fluorescence, line tension, cholesterol
Monolayers of lipids as well as other amphiphiles at the air–water interface often display coexisting thermodynamic phases. These phases are generally observed with optical microscopic techniques (1–3). The domains of one phase, which are surrounded by a second phase, typically exhibit a wide variety of shapes and sizes that are strongly affected by a competition between long-range dipole–dipole forces and line tension forces. The theoretical description of the various shapes of lipid domains is in satisfactory agreement with experimental observations for those monolayers where the molecular dipoles are on average perpendicular to the monolayer surface. Leading references to recent work are given in ref. 4.
An outstanding problem has concerned the equilibrium sizes of lipid domains in certain lipid mixtures. In some of these mixtures, described later, a wide variety of domain sizes are seen in a single monolayer (1–3). A recent thermodynamic treatment of the equilibrium sizes of lipid domains shows that this broad distribution of sizes is not compatible with thermodynamic equilibrium (4). It has also been shown that domains may assume a variety of metastable equilibrium sizes, but we suspect that such metastabilities cannot account for the permanence of broad distributions of sizes sometimes seen. The purpose of the present paper is to show that in certain lipid mixtures, a theoretical rate of size equilibration is exceedingly small, thus accounting, at least qualitatively, for the experimental results.
Background
Certain binary mixtures of lipids, specifically mixtures of phosphatidylcholines and cholesterol, exhibit coexisting liquid phases under defined conditions of temperature, pressure, and composition. Such mixtures are ideal for testing theoretical models of domain sizes and shapes, because the molecular dipoles are, on average, perpendicular to the plane of the air–water interface, thus greatly simplifying the calculation of dipole–dipole energies. These mixtures are also particularly useful since they exhibit critical mixing/demixing as a function of monolayer pressure. One can experimentally modulate both line tension and dipole density simply by changing monolayer pressure (2, 4).
An example of circular lipid domains in a mixture of 30 mol % cholesterol and 68 mol % dipalmitoyl phosphatidylcholine is given in Fig. 1. The monolayer contains 2 mol % fluorescent lipid probe. [Experimental details are the same as those given by Rice and McConnell (5).] The “black” (weakly fluorescent) domains are rich in cholesterol, and the “white” (strongly fluorescent) domains are rich in the phosphatidylcholine. It will be seen that there is a wide variety of domain sizes, with diameters ranging all the way from the resolution of the fluorescence microscope (≈1 μm) to 100 μm. This diversity of domain sizes may be contrasted with the near uniformity of domain sizes that has sometimes been observed in monolayers of other compositions, especially monolayers with a single component. The domain pattern seen in Fig. 1 shows no major changes over periods of many minutes, and perhaps many hours.
Figure 1.
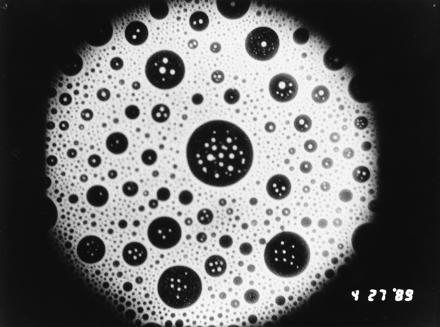
Epifluoresence microscope photo of a lipid monolayer at the air–water interface composed of 68 mol % dipalmitoyl phosphatidylcholine, 30 mol % cholesterol, and 2 mol % fluorescent lipid probe. The experimental conditions are the same as those given earlier (5). The dark liquid domains are rich in cholesterol, and the white liquid domains are rich in the phosphatidylcholine. The large domain in the center of the photo has a diameter of ≈100 μm (P. Rice and H.M.M., unpublished work).
In previous theoretical work (2, 4), it has been shown that in an ensemble of weakly interacting circular domains—all far removed from one another—the domains have a theoretical equilibrium radius Rq, where:
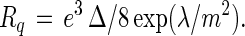 |
1 |
Here m is the difference in dipole density in the two phases, λ is the line tension associated with the interface between the two phases, and Δ is a dipole–dipole cut-off distance. This cut-off distance can be thought of as a distance of closest approach between neighboring dipoles in the monolayer, and for lipids in the liquid state it is taken to be of the order of magnitude of 10 Å. The energy of an isolated circular domain of radius R can be expressed in terms of Rq as follows:
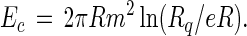 |
2 |
This energy expression is referred to as an “edge energy,” since it gives that part of the total thermodynamic energy that is nonlinear in the size of the domain. The line tension and the long-range part of the dipolar electrostatic energy are included in this edge energy. The minimization of the energy of an ensemble of noninteracting circular domains with constant total area leads to the equilibrium radius in Eq. 1.
The large breadth of the distribution of domain sizes seen in Fig. 1 is not surprising in that these domains are formed by “quenching”—that is, the pressure of a homogeneous monolayer is reduced, often through the critical pressure, until the two phases are formed. The distribution of domain sizes is determined by the kinetics of this phase separation. What is surprising is the long lifetime of this broad distribution in the cholesterol/phosphatidylcholine mixtures. The heterogeneous domain patterns persist for many minutes, probably hours, with seemingly little change. This slow rate of domain size equilibration might plausibly be due to the formation of long-lived metastable equilibrium states, as discussed previously (4). Alternatively, the persistence of these heterogeneous patterns might be due to an intrinsically slow rate of equilibration. The following order-of-magnitude calculation shows that the intrinsic rate of domain size equilibration is very small indeed.
The Kinetic Model
Fig. 2 depicts a doughnut-shaped lipid domain with a central black circular domain of radius Ri. This is surrounded by a white domain torus with inner radius Ri and outer domain radius Ro. The white torus is in turn surrounded by a black domain torus with inner radius Ro and fixed outer domain radius. We consider processes in which the size of the central domain increases or decreases and in which the area of white phase and total area of black phase, remain constant. The compositions of the black and white phases also remain constant in this process, where XA and XB are the mole fractions of A and B in the black phase domains, and YA and YB are the mole fractions of A and B in the white phase domain. To clarify the discussion, assume that XA > YA and XB < YB. Suppose further that in the process of equilibration the central black domain becomes smaller and ultimately disappears. With this picture there is a flux of A molecules out of the central black domain, through the white domain, and into the outer black domain. Likewise there is a flux of B molecules out of the outer black domain, and through the white domain, so as to replace the A molecules that leave the central black domain. For some lipid mixtures, one is justified in assuming that the molecular areas of the A and B molecules are approximately the same. In this case there is no net molecular flux of molecules in the monolayer, and thus no macroscopic hydrodynamic flow in the subphase, even though there are oppositely directed fluxes of the A and B molecules. In the following calculation, these oppositely directed molecular fluxes are related to the change in the size of the central domain.
Figure 2.
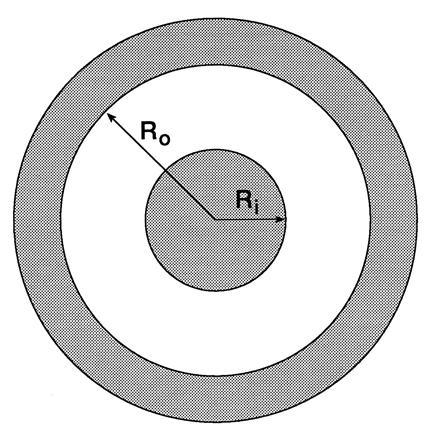
The configuration of domains used for the model calculation of the equilibration rate. The outer radius of the black torus domain is fixed, whereas the radius of the inner domain, Ri, and the inner radius of the torus, Ro, change with time. The area of the white phase is fixed, as is the total area of the black phase. The monolayer is assumed to have two chemical components, each with equal molecular areas.
Consider an incremental change in the radius of the inner domain, dRi, where this region is initially occupied by black phase, and is subsequently occupied by white phase. The number of molecules in this incremental region is 2πcRidRi, where c is the two-dimensional lipid concentration, in molecules per square centimeter. When this region is occupied by black phase, it contains a number of A molecules equal to 2πXAcRidRi. After the change, the incremental region is occupied by 2πYAcRidRi molecules of A. Thus the outward flux of A molecules due to this incremental change is:
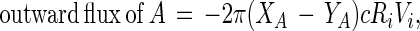 |
3 |
where Vi = dRi/dt. Under steady state conditions, this outward flux of A molecules in the white phase must be constant, independent of radius R. Thus:
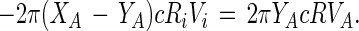 |
4 |
In the above equation Vi is defined as the rate at which the radius of the inner circle changes, whereas VA is the (average) molecular velocity of the A molecules moving outward through the white phase at the radius R. From Eq. 4 and the corresponding equation for the B molecules, one obtains simply:
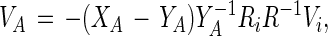 |
5 |
and
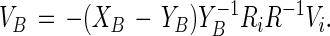 |
6 |
Note that the average molecular velocities of the A and B molecules are not equal to one another. The net molecular flux, c(YAVA + YBVB), is nonetheless equal to zero.
The molecular velocities VA and VB are related to the forces FA and FB that arise from gradients in the chemical potentials of A and B. Thus VA = μAFA and VB = μBFB, where we take the mobilities μA and μB to be equal to one another and equal to D/kT, where D is the molecular diffusion coefficient in the monolayer. The steady state viscous dissipation per unit time in the monolayer is the integral of c(YAVAFA + YBVBFB) over the white domain, which is simply:
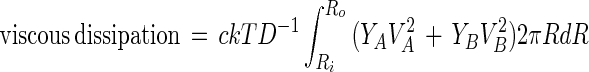 |
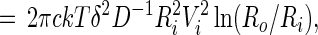 |
7 |
where:
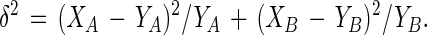 |
8 |
We equate the rate of viscous dissipation of energy to the change in the electrostatic and line tension energy E for the domain configuration in Fig. 2. This energy is:
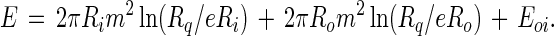 |
9 |
The first and second terms on the right hand side of Eq. 9 are the edge energies of circular domains with radii equal to Ri and Ro, and Eoi is an interaction energy between the edges. [This energy involves elliptic integrals (6).] We shall assume that Ro ≫ Ri, in which case this interaction energy can be neglected. The rate of energy loss is then:
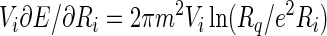 |
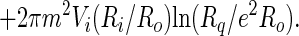 |
10 |
Again, when the inner radius is much smaller than the outer radius, the calculation can be simplified, this time by neglecting the second term on the right side of Eq. 10. When this rate of energy change in equated to the negative of the rate of viscous dissipation of energy, one obtains:
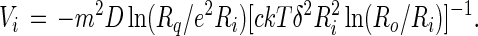 |
11 |
As noted elsewhere (4), the energy of an isolated circular domain is a maximum when R = Rq/e2, and accordingly Vi in Eq. 11 changes sign at this point. That is, the domain decreases in size when Ri < Rq/e2 and increases in size when Ri > Rq/e2.
DISCUSSION
The rate of equilibration of a circular lipid domain according to Eq. 11 can be conveniently expressed as a relative rate:
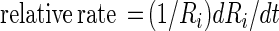 |
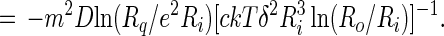 |
12 |
In centimeter-gram-second electrostatic units (cgs-esu), orders of magnitude of the various terms in Eq. 12 are:
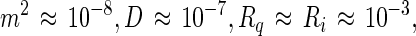 |
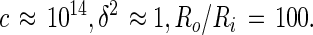 |
It will be seen that the relative rate of change of the inner radius is very small, of the order of one part in a million per second. It would thus take of the order of a week for there to be a substantial change in the size of a lipid domain under these conditions. Of course the Ri−3 dependence of the relative rate in Eq. 12 insures that very small domains, with radii of the order of 0.1 μm, will disappear rapidly, assuming the equilibrium radius is of the order of 10 μm. The rate of change of the size of the (central) domain depends on its proximity to the outer torus, which acts as source or sink. However the dependence on this proximity is only logarithmic. In spite of the approximations involved, this result provides an explanation for the apparent slow rates of equilibration of the sizes of lipid domains with two chemical components.
An important assumption made in the calculation is that the densities of the black and white phases are substantially the same. This is roughly true for the saturated phosphatidylcholine/cholesterol mixture illustrated in Fig. 1 and is true to a good approximation for mixtures of unsaturated phosphatidylcholines and dihydrocholesterol (J. Hagen & H.M.M., unpublished data). Under this condition, there is little macroscopic hydrodynamic flow in the aqueous subphase of the monolayer. In monolayer systems where the coexisting phases have significantly different densities, such as liquid and gas, or liquid and solid there can be long-range hydrostatic forces in the monolayer, as well as macroscopic hydrodynamic drag in the subphase. The net result in such systems may be a larger equilibration rate. [These systems do sometimes show domains with remarkably uniform sizes (7).] The calculated equilibration rates in the present work are to be contrasted with the rapid equilibration rates observed previously in these same monolayers when acted on by an externally applied electric field (8). The energy associated with the applied field greatly exceeds the “self-energy” due to long range dipole–dipole forces and line tension considered here.
In conclusion it should be noted that in previous work, it was pointed out that weakly interacting circular domains will in general approach a metastable equilibrium state in which all the circular domains have equal radii (2, 4). Of course this does not happen in the model used in the present paper, where the only choice allowed for the final metastable equilibrium state is either a circular black domain at the center or a circular white domain at the center. Note particularly that even a black domain at the center having an “equilibrium” radius will grow in size in the present model calculation until all, or almost all, the black phase is at the center. The lowest energy state of the system with the assumed symmetry depends on the nature of the boundary at the fixed outer radius of the outer domain, but this is of no interest for the present purposes.
Acknowledgments
This work was supported by National Science Foundation Grant MCB9316256.
References
- 1.Möhwald H. Annu Rev Phys Chem. 1990;41:441–476. doi: 10.1146/annurev.pc.41.100190.002301. [DOI] [PubMed] [Google Scholar]
- 2.McConnell H M. Annu Rev Phys Chem. 1991;42:171–195. [Google Scholar]
- 3.Andelman D, Brochard F, Knobler C M, Rondelez F. In: Micelles, Membranes, Microemulsions and Monolayers. Gelbard W M, Ben-Shaul A, Roux D A, editors. New York: Springer; 1994. pp. 559–602. [Google Scholar]
- 4.McConnell H M. Langmuir. 1996;12:4897–4904. [Google Scholar]
- 5.Rice P A, McConnell H M. Proc Natl Acad Sci USA. 1989;86:6445–6448. doi: 10.1073/pnas.86.17.6445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.McConnell H M, Moy V T. J Phys Chem. 1988;92:4520–4525. [Google Scholar]
- 7.Lösche M, Möhwald H. Eur Biophys J. 1984;11:35–42. doi: 10.1007/BF00253856. [DOI] [PubMed] [Google Scholar]
- 8.Lee K Y C, McConnell H M. Biophys J. 1995;68:1740–1751. doi: 10.1016/S0006-3495(95)80351-6. [DOI] [PMC free article] [PubMed] [Google Scholar]