

Abstract
GAIP (G Alpha Interacting Protein) is a member of the recently described RGS (Regulators of G-protein Signaling) family that was isolated by interaction cloning with the heterotrimeric G-protein Gαi3 and was recently shown to be a GTPase-activating protein (GAP). In AtT-20 cells stably expressing GAIP, we found that GAIP is membrane-anchored and faces the cytoplasm, because it was not released by sodium carbonate treatment but was digested by proteinase K. When Cos cells were transiently transfected with GAIP and metabolically labeled with [35S]methionine, two pools of GAIP—a soluble and a membrane-anchored pool—were found. Since the N terminus of GAIP contains a cysteine string motif and cysteine string proteins are heavily palmitoylated, we investigated the possibility that membrane-anchored GAIP might be palmitoylated. We found that after labeling with [3H]palmitic acid, the membrane-anchored pool but not the soluble pool was palmitoylated. In the yeast two-hybrid system, GAIP was found to interact specifically with members of the Gαi subfamily, Gαi1, Gαi2, Gαi3, Gαz, and Gαo, but not with members of other Gα subfamilies, Gαs, Gαq, and Gα12/13. The C terminus of Gαi3 is important for binding because a 10-aa C-terminal truncation and a point mutant of Gαi3 showed significantly diminished interaction. GAIP interacted preferentially with the activated (GTP) form of Gαi3, which is in keeping with its GAP activity. We conclude that GAIP is a membrane-anchored GAP with a cysteine string motif. This motif, present in cysteine string proteins found on synaptic vesicles, pancreatic zymogen granules, and chromaffin granules, suggests GAIP’s possible involvement in membrane trafficking.
Keywords: G protein, regulator of G-protein signaling, GTPase-activating protein, cysteine string
Using the yeast two-hybrid system, we recently identified GAIP, a human protein that specifically interacts with the heterotrimeric G protein Gαi3 (1). GAIP is a member of the newly described RGS family (for Regulators of G-protein Signaling) (1–5) whose ≈15 members share an ≈125-aa homologous core domain and are thought to regulate G-protein signaling. This core domain, now referred to as the RGS domain, is the site of interaction with the Gα subunit (1). Mutants of two RGS family members, EGL-10 in Caenorhabditis elegans and Sst2 in Saccharomyces cerevisiae, show a delay in egg-laying behavior (3) and desensitization to pheromone (6), respectively. Another family member, RGS4, was shown to inhibit mitogen-activated protein (MAP) kinase activity stimulated through G-protein-coupled receptors (2).
The recent demonstration that GAIP, RGS4, and other RGS proteins function as GTPase-activating proteins (GAPs) for Gαi subunits in vitro (7–9) indicates that these molecules negatively regulate heterotrimeric G proteins by stimulating their intrinsically low GTPase activity, returning them to the inactive GDP-bound state. A number of GAPs have been isolated for the small GTP-binding proteins. The distribution and interaction of rasGAP with ras is particularly well documented (10, 11). To date no information is available on the distribution and the nature of the interaction with G proteins for any member of the RGS family. In this paper we investigated the distribution of GAIP and the nature of the GAIP/Gαi3 interaction in cells stably or transiently expressing GAIP in vivo. We provide evidence that GAIP interacts preferentially with the GTP-bound form of members of the Gαi subfamily. We also obtained the surprising finding that, in contrast to most other GAPs, GAIP is membrane-anchored.
MATERIALS AND METHODS
Cloning Procedures.
cDNAs encoding the full-length human GAIP and the fragment GAIP23–217 were inserted into the EcoRI (5′) and XhoI (3′) sites of mammalian expression vector pCDNA3 (Invitrogen). An HA epitope (derived from hemagglutinin protein of human influenza virus) was fused to the N terminus of GAIP by ligating autohybridized oligonucleotides coding for the epitope (YPYDVPDYA) into the BamHI (5′) and EcoRI (3′) sites of the above vector (pCDNA3 HA-GAIP1-217 and pCDNA3 HA-GAIP23–217). An HA epitope was placed at the alternative splice site of the long form of the rat Gαs as previously described (12). The plasmids used for coupled in vitro transcription–translation (Promega TnT kit) and for production of glutathione S-transferase (GST)-GAIP fusion protein were as described (1). Full-length Gα subunit cDNA sequences were constructed in the pGBT9 bait vector (Clontech) after PCR on their respective vectors as templates. The rat Gαi3(Q204L) and Gαi3(G203A) mutants were obtained from A. Spiegel (National Institutes of Health); rat Gαi2, mouse Gαq, and mouse Gα13, from P. Insel (University of California, San Diego); rat Gαi1 and the rat Gαs(Q227L) mutant, from T. Kozasa (University of Texas, Southwestern Medical Center); rat Gαo from E. Neer (Brigham and Women’s Hospital); rat Gαz, from E. Ross (University of Texas, Southwestern Medical Center); S. cerevisiae Gpa1 from J. Noel (Salk Institute); and rat Gαs, from H. Bourne (University of California, San Francisco). The C-terminal 10-aa truncation mutant, Gαi3(Δ345–354) of Gαi3, the Gαq-Gαi3 C-terminal chimera, Gαq/i3(345–354)—a 10-aa swap between Gαq and Gαi3 C termini, and the Gαi3(G352N) point mutant constructs were made by PCR with modifying primers (sequences available upon request). All constructs were verified by automated sequencing through the cloning sites and through the mutated regions using 5′ and 3′ pGBT9 sequencing primers.
A rat GAIP ortholog (rGAIP) isolated from a rat pituitary library (in pACT2 prey vector) by a yeast two-hybrid screen with rat Gαi3 as bait was used in some of the yeast two-hybrid assays described below. No noticeable differences in interaction levels were detected between the rat and human proteins (data not shown).
Cell Culture and Transfections.
Cos-7 monkey kidney cells were grown in Dulbecco’s modified Eagle’s medium (DMEM-high glucose) supplemented with 10% (vol/vol) fetal calf serum, penicillin G, and streptomycin sulfate. Murine AtT-20/D-16v pituitary cells, obtained from Richard Mains (Johns Hopkins University), were grown in DMEM-high glucose with 10% horse serum. For transient transfections of Cos-7 cells with full-length GAIP, cells grown in 75-cm2 flasks were transfected with 5 μg of the pCDNA3 HA-GAIP1–217 or pCDNA3 HA-Gαs plasmids by using the DEAE-dextran method (13). For stable transfections 50 μg of pCDNA3 HA-GAIP23–217 plasmid was transfected into AtT-20 cells by using Lipofectin (GIBCO/BRL). After 3 days of nonselective growth in complete medium, followed by 14 days of growth in the same medium with geneticin (G418, GIBCO/BRL) at 500 μg/ml, clones were selected by serial dilution of surviving foci and maintained in geneticin at 100 μg/ml. Expression of GAIP23–217 was verified by immunoblotting, and eight clones showing different levels of expression were obtained. A more detailed analysis of the cell lines stably expressing GAIP23–217 will be described elsewhere. Two of these clones (clones 1 and 14) were used in this study.
Antibodies.
mAb 12CA5 against the HA epitope was purchased from Boehringer Mannheim. Polyclonal rabbit antibodies against calnexin (14) and Cab45 (15) were obtained from J. Bergeron (McGill University) and H. Lodish (Whitehead Institute), respectively. Affinity-purified anti-β-COP IgG, characterized earlier (16), was raised against the EAGE peptide of Duden et al. (17).
Preparation and Analysis of Membrane Fractions.
Confluent cultures of AtT-20 clones 1 and 14 stably expressing HA-GAIP23–217 were harvested by scraping into ice-cold phosphate-buffered saline (PBS) containing protease inhibitors (1 mM phenylmethanesulfonyl fluoride, 2 mg/ml aprotinin, 0.5 mg/ml leupeptin, and 5 mM EDTA). Cells were passed 10 times through a 30-gauge needle, and the lysate was centrifuged (600 × g for 3 min at 4°C) in a microcentrifuge to pellet unbroken cells and nuclei. The supernatant was centrifuged at 100,000 × g for 1 hr at 4°C in a Beckman TLA45 rotor. The pellet (crude membrane fraction) was resuspended in ice-cold PBS containing protease inhibitors and divided into aliquots (≈75 μg of protein). Some aliquots were treated with 0.1 M Na2CO3 (pH 11.3) for 30 min at 4°C and centrifuged again at 100,000 × g for 1 hr (18). Others were treated with 50 μg/ml proteinase K (GIBCO/BRL) for 25 min at 4°C. Proteins from all fractions—i.e., supernatants and untreated and treated pellets—were separated by SDS/PAGE (19) and transferred to poly(vinylidene difluoride) (PVDF) membranes (Millipore), using a semidry blotter (Millipore). Immunoblotting was performed for epitope-tagged GAIP using mAb 12CA5 (2-hr incubation, dilution 1:80) and for Cab45, calnexin, and β-COP using polyclonal rabbit antibodies (1-hr incubation, dilutions 1:3000, 1:10,000, and 1:3000, respectively). Secondary antibodies consisted of horseradish peroxidase (HRP)-conjugated goat anti-mouse or anti-rabbit IgG (Bio-Rad) (1-hr incubation, dilution 1:3000). The ECL (enhanced chemiluminescence) kit from Amersham was used for detection of immunoreactivity. The bands corresponding to GAIP were quantified by laser scanning densitometry (Ultra Scan XL, LKB).
Metabolic Labeling and Immunoprecipitation.
For labeling with [3H]palmitic acid, Cos-7 cells in 75-cm2 flasks were transfected 48 hr earlier and incubated in DMEM-high glucose for 2 hr, after which the medium was changed to 5 ml of DMEM-high glucose containing 1% dimethyl sulfoxide and 2.5 mCi of [9,10-3H]palmitic acid (specific activity 60 Ci/mmol; American Radiolabeled Chemicals; 1 Ci = 37 GBq), and incubation was resumed for 1 hr. For labeling with [35S]methionine, cells were incubated 1 hr in DMEM-high glucose (methionine-free) containing 200 μCi of Met-35S-Label per ml (1175 Ci/mmol; American Radiolabeled Chemicals). Cells were washed three times in PBS and harvested by scraping in homogenization buffer [5 mM Hepes, pH 7.4/1 mM EDTA/100 mg/ml soybean trypsin inhibitor/0.5 mg/ml leupeptin/2 mg/ml aprotinin/0.7 mg/ml pepstatin/10 milliunits/ml α2-macroglobulin (Boehringer Mannheim)]. The cell suspension was centrifuged (5 min at 180 × g) at 4°C and the cell pellets were homogenized by passing them through a 25-gauge needle 25 times. Crude membrane and soluble fractions were prepared as described for the AtT-20 cells except the crude membrane fraction was resuspended in homogenization buffer. Immunoprecipitation was performed on either 1 mg of protein from the [3H]palmitic acid-labeled cells or 100 μg of protein from the [35S]methionine-labeled cells in 1 ml of solubilization buffer (50 mM Tris·HCl, pH 7.5/150 mM NaCl/1% Triton X-100/0.2% SDS/1 mM EDTA). One microgram of the 12CA5 mAb or 10 μg of nonimmune rabbit IgG was added, and the samples were incubated overnight at 4°C with gentle mixing. The immunoprecipitates were recovered by incubation for 2 hr with protein A-Sepharose CL-4B (Pharmacia LKB) and washed twice in solubilization buffer and once in the same buffer without detergents. After centrifugation (8000 × g for 10 min) the immunoprecipitate was solubilized in Laemmli gel loading buffer (19), separated by SDS/PAGE on 8–16% gradient Tris–glycine gels (Novex), and prepared for autoradiography using EN3HANCE (DuPont/New England Nuclear) and Biomax-MR film (Kodak). The labeled products were quantified by densitometry using scan analysis (Biosoft, Cambridge, U.K.) software.
Two-Hybrid Assays.
Filter and liquid β-galactosidase (β-gal) assays were as described (1) except that yeast strain SFY526 was used on selective medium (Clontech). Transformations of yeast were performed according to Schiestl and Gietz (20).
In Vitro Interactions.
Purified GST-GAIP was prepared, immobilized on glutathione-agarose beads (Pharmacia LKB), and incubated with 35S-labeled in vitro translated Gαi3 as described (1), except that 5 μM guanosine 5′-[γ-thio]triphosphate (GTP[γS]) or AlF4− was added to some aliquots of the in vitro translated Gαi3 and to the washes before incubating the sample with GST-GAIP. The labeled products were identified and quantified by autoradiography using a PhosphorImager (Molecular Dynamics) and imagequant software.
blast Searches.
Online blast searches were performed through the National Center for Biotechnology Information at the National Institutes of Health, Bethesda, MD (21).
RESULTS
GAIP Interacts with the Gαi Subfamily.
We have shown previously that GAIP interacts specifically with Gαi3 but not with Gαq. Using the yeast two-hybrid system (22), we investigated the specificity of GAIP for all Gα subfamilies. As shown in Table 1, Gαi1, Gαi2, Gαi3, Gαo, and Gαz, all considered Gαi subfamily members (24), interacted with GAIP in the two-hybrid assay although not to the same degree (Gαi3 > Gαi1, Gαo ≫ Gαz, Gαi2). Other G-protein subfamilies such as Gαs, an activated Gαs(Q227L) mutant (25), Gαq, and Gα13 showed no interaction. These results demonstrate the specificity of GAIP for the Gαi subfamily.
Table 1.
GAIP interacts specifically with Gαi subfamily members
| Bait | β-gal
|
|
|---|---|---|
| Filter | Liquid, % | |
| Gαi1 | +++ | 55 |
| Gαi2 | + | 6.3 |
| Gαi3 | +++ | 100 |
| Gα0 | +++ | 52 |
| Gαz | + | 5.0 |
| Gαs | — | <1 |
| Gαs(Q224L) | — | <1 |
| Gαq | — | <1 |
| Gα13 | — | <1 |
The β-gal filter assay was performed on (Leu−, Trp−) plates, and intensity of color was scored after 8 hr. —, no color; +, weak color; +++, strong color. For the β-gal liquid assay (Leu−, Trp−), the Gαi3/GAIP interaction [12.5 Miller units (23)] was taken as 100%. Yeast cotransformed with void bait and prey vectors were taken as background. Baits were constructed in pGBT9, and GAIP prey vector was pACT2-rGAIP. For each experiment three colonies were picked. Values represent the mean of three independent experiments.
The C Terminus of Gαi3 Is Important for Interaction with GAIP.
The fact that GAIP interacts specifically with the Gαi subfamily led us to investigate the importance of the C terminus of Gαi3 for interaction with GAIP in more detail, because the two signatures for the Gαi subfamily—i.e., pertussis toxin sensitivity (26) and the glycine residue in the third position from the C terminus (27, 28)—are located there, and the last 10 amino acids of Gα subunits have been implicated in specific receptor interaction (29). Using the two-hybrid system, we tested a C-terminal deletion mutant of Gαi3, truncated by 10 amino acids [Gαi3(Δ345–354)]. As shown in Table 2, deletion of 10 amino acids at the C terminus of Gαi3 is sufficient to significantly reduce its interaction with GAIP. Previous studies have shown that Gαq does not interact with GAIP (1), and Gαq has an asparagine instead of a glycine three residues from the C terminus. We also investigated the importance of the glycine residue (G352) in the third position from the C terminus of Gαi3 by replacement of G352 with the corresponding asparagine residue (N357) in Gαq. We observed a small but significant drop in the degree of interaction between Gαi3 and GAIP (G352N) (Table 2). We also constructed a Gαq/Gαi3 chimera [Gαq/i3(345–354)], in which we replaced the 10 C-terminal amino acids of Gαq with those of Gαi3. As shown in Table 2, the Gαq/i3(345–354) chimera did not interact with GAIP. The yeast homolog Gpa1, which can be considered a Gαi family member based on the fact that it contains the signature glycine residue (30), showed the strongest interaction with GAIP in the β-gal liquid assay (Table 2). The above results suggest the importance of the C terminus of the Gαi subfamily in their interaction with GAIP. However, based on the lack of interaction with the Gαq/i3 chimera, we conclude that the extreme C terminus of Gαi3 is not the only site of binding to GAIP.
Table 2.
GAIP interacts with the C terminus of Gαi3
| Bait | β-gal
|
|
|---|---|---|
| Filter | Liquid, % | |
| Gαi3 | +++ | 100 |
| Gαq | — | <1 |
| Gαi3(Δ345–354) | + | 14 |
| Gαi3(G352N) | +++ | 80 |
| Gαq/i3(345–354) | — | <1 |
| Gpa1 | +++ | 160 |
Footnotes are as for Table 1. Values represent the mean of five independent experiments.
GAIP Interacts with the GTP-Bound Form of Gαi3.
To determine whether GAIP preferentially interacts with the GTP- or GDP-bound form of Gαi3 in vivo, we assessed the interaction between GAIP and point mutants of Gαi3, using the yeast two-hybrid system. In the filter assay (Table 3), the interaction between GAIP and Gαi3(G203A), the inactivated (GDP) form of Gαi3, was significantly weaker than that between GAIP and Gαi3(Q204L), the activated (GTP) form (25, 31, 32). These results were quantitated by the β-gal liquid assay (Table 3), where the interaction between GAIP and Gαi3(G203A) was at least 20 times weaker than that between GAIP and Gαi3(Q204L). The yeast two-hybrid results were reinforced by results of an in vitro assay. GST-GAIP fusion protein bound to glutathione-agarose beads interacted with in vitro transcribed–translated Gαi3 (Fig. 1, lane 1), and this interaction was enhanced 4-fold in the presence of GTP[γS] (Fig. 1, lane 2) and 5-fold in the presence of AlF4− (Fig. 1, lane 3). GTP[γS], a nonhydrolyzable analog of GTP, maintains the Gα subunit in its activated state. AlF4− also activates GDP-bound heterotrimeric G proteins by mimicking the GDP-to-GTP transition state during GTP hydrolysis (33, 34). These results indicate that GAIP interacts preferentially, if not exclusively, with the GTP-bound state of Gαi3.
Table 3.
GAIP interacts with the GTP-bound form of Gαi3
| Bait | β-gal
|
|
|---|---|---|
| Filter | Liquid, % | |
| Gαi3 | +++ | 100 |
| Gαi3(Q204L) | +++ | 110 |
| Gαi3(G203A) | + | 5 |
Footnotes are as for Table 1. GAIP prey vector was pGADGH-GAIP1-217. Values represent the mean of two experiments.
Figure 1.
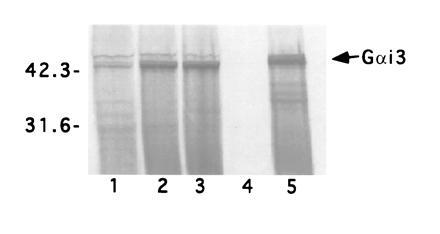
GAIP interacts with the GTP-bound form of Gαi3 in vitro. GST-GAIP fusion protein bound to glutathione-agarose beads was incubated with in vitro translated Gαi3 for 2 hr in the presence or absence of GTP[γS] and AlF4−. The bound products were separated by SDS/10% PAGE and detected by autoradiography. 35S-labeled in vitro translated Gαi3 binds to GST-GAIP beads 4–5 times more efficiently in the presence of GTP[γS] (lane 2) or AlF4− (lane 3) than in their absence. Lane 4, control beads with GST alone. Lane 5, 35S-labeled in vitro translated Gαi3 (arrow). Molecular mass markers (kDa) are indicated on the left.
GAIP Is a Membrane Protein.
Most GAPs for small GTP-binding proteins are cytosolic proteins (8), with one possible exception—i.e., rasGAP (35). To determine whether GAIP is a membrane protein or a cytosolic protein, we assessed the distribution of GAIP in crude membrane (100,000 × g pellet) and cytosolic (100,000 × g supernatant) fractions prepared from AtT-20 cells stably expressing GAIP. We found that 80–90% of the GAIP pelleted with the membrane fraction (Fig. 2). When this fraction was treated with 0.1 M Na2CO3 (pH 11.3) to strip peripheral membrane proteins (18), GAIP remained associated with the membrane fraction (Fig. 3A), indicating that it behaves as an integral membrane protein. By contrast, sodium carbonate treatment resulted in release of Cab45 (Fig. 3B), a soluble luminal protein of the Golgi (15), into the cytosolic fraction, indicating that the high pH treatment was effective. Digestion of the membrane pellet with proteinase K resulted in complete digestion of GAIP (Fig. 4A) and β-COP (Fig. 4B), a peripheral coat protein that faces the cytoplasm (17). Under the same conditions calnexin, an integral membrane protein that faces the lumen of the endoplasmic reticulum (14), was protected (Fig. 4C). Taken together, the above results show that (i) there are two pools of GAIP, a membrane and a soluble pool, and (ii) the membrane-associated pool faces the cytoplasm.
Figure 2.
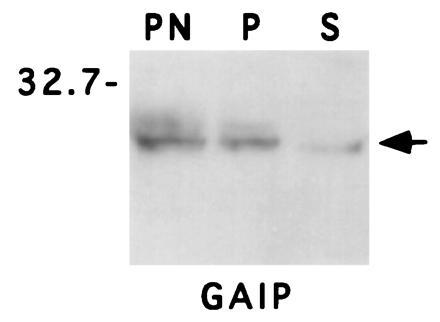
GAIP is present in both membrane and soluble fractions. A postnuclear supernatant (PN) from AtT-20 cells stably expressing GAIP (clone 14) was centrifuged at 100,000 × g to yield a crude membrane fraction (P) and a soluble fraction (S). These fractions were immunoblotted with 12CA5 mAb (anti-HA) and detected by ECL, and the amount found in the membrane and cytosolic fractions was quantified by densitometry. Most of the GAIP (80–90%) is associated with the membrane pellet, but the remainder is found in the soluble fraction.
Figure 3.
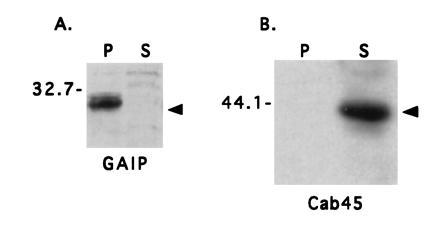
GAIP behaves as an integral membrane protein. Membrane fractions (100,000 × g pellet) from AtT-20 cells stably expressing GAIP (clone 14) were treated with Na2CO3. Proteins were separated by SDS/12% PAGE and immunoblotted. (A) GAIP remains associated with the membrane fraction (P) after Na2CO3 treatment and is not detected in the soluble fraction (S). (B) Cab45, a soluble luminal Golgi protein, is released from the membrane fraction (P) and appears in the supernatant (S) after Na2CO3 treatment.
Figure 4.
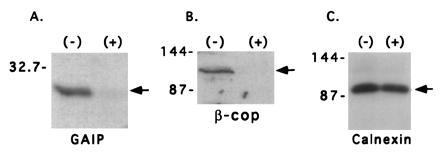
GAIP faces the cytoplasm. Membrane fractions (100,000 × g pellet) from AtT-20 cells stably expressing GAIP (clone 14) were treated with proteinase K (+) or buffer alone (−), after which the membranes were solubilized, separated by electrophoresis on SDS/12% polyacrylamide gels, and immunoblotted. Detection was by ECL. (A) GAIP is digested and not detectable after proteinase K treatment. (B) β-COP, a peripheral coat protein facing the cytoplasm, is also digested by proteinase K. (C) Calnexin, a membrane protein facing the lumen of the endoplasmic reticulum, is protected from proteinase K digestion.
GAIP Has a Cysteine String Motif and Is Palmitoylated.
GAIP lacks a signal peptide and from its hydropathy plot has no evident transmembrane domain (1). A blastp search (21) of the N-terminal 79 residues (exclusive of the RGS domain) of GAIP revealed that GAIP possesses a cysteine-rich region (8 of 11 residues are cysteines) between residues 39 and 49, analogous to those in cysteine string proteins (Fig. 5). Cysteine string proteins are a family of proteins found on synaptic vesicles in Drosophila, Torpedo, and rat brain (36–38). The fact that cysteine string proteins are heavily palmitoylated on their cysteine string motif (39, 40) suggested that GAIP might also be palmitoylated. To investigate this possibility we metabolically labeled HA-GAIP-transfected Cos cells with [3H]palmitate or [35S]methionine and immunoprecipitated HA-GAIP from the membrane and cytosolic fractions with anti-HA mAb. 35S-labeled HA-GAIP was found in both the supernatant (80%) and the membrane pellet (20%) (Fig. 6A). The increased amount of GAIP in the soluble fraction of the Cos cells compared with AtT-20 cells may be due to a greater level of overexpression with transient transfection. By contrast, 3H-labeled HA-GAIP was present exclusively in the membrane pellet (Fig. 6B). The fact that there are two pools of GAIP and that the palmitoylated form is found exclusively in the membrane pellet in transfected Cos cells suggests that GAIP is anchored to membranes by means of palmitoylation. To compare levels of [3H]palmitate incorporation, cells were also transfected with HA-tagged Gαs because this protein has only one site of palmitoylation (41). While the amount of 35S-labeled HA-Gαs was equivalent to the amount of the 35S-labeled HA-GAIP in the membrane pellet, the amount of 3H-labeled HA-GAIP was much greater than the 3H-labeled HA-Gαs (data not shown), suggesting that GAIP is more heavily palmitoylated than Gαs.
Figure 5.

GAIP has a cysteine string motif. The alignment shows the cysteine string motifs in six cysteine string proteins (Csp) from Drosophila, Torpedo, rat, and bovine origin, two human expression sequence tags (GenBank number given) that are putative cysteine string proteins, human GAIP, and two yeast proteins of unknown function. Alignment of cysteine string motifs was based on blastp and blastn searches in GenBank/SwissProt with GAIP’s cysteine-rich domain (amino acids 39–49) as query. The number of cysteines in the motif varies from 8 in GAIP and yeast YBW3 to 16 in human T60736.
Figure 6.
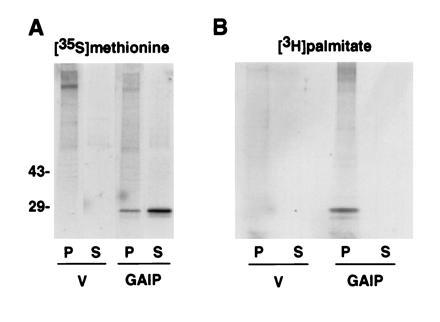
GAIP is palmitoylated in vivo. COS cells were transfected with pCDNA3 vector alone (V) or with pCDNA3 HA-GAIP1-217 vector (GAIP) and radiolabeled 48 hr later with either [35S]methionine (A) or [3H]palmitic acid (B). The cells were homogenized and membrane (P) and soluble (S) fractions were prepared as described for Fig. 2. Samples, 100 μg of the [35S]methionine-labeled proteins and 1 mg of the [3H]palmitate-labeled proteins, were immunoprecipitated with the 12CA5 mAb and analyzed by SDS/PAGE and fluorography. The film was exposed at −70°C for 7 days (A) or 30 days (B). GAIP is distributed into two pools: 20% of the [35S]methionine-labeled GAIP sediments with membrane fraction (P), and 80% is present in the cytosolic (S) fraction. Palmitoylated GAIP is detected only in the membrane fraction (P).
DISCUSSION
Our main finding is that GAIP is membrane-anchored. In AtT-20 cells stably expressing GAIP we found 80–90% of the GAIP in a crude membrane fraction (100,000 × g pellet). Resistance to stripping with Na2CO3 and susceptibility of the membrane fraction to digestion with proteinase K showed that GAIP is membrane-anchored and faces the cytoplasm. A blast search revealed that GAIP has a cysteine string motif, which is heavily palmitoylated in cysteine string proteins. We further demonstrated that GAIP can be palmitoylated in vivo and that palmitoylated GAIP is located solely in the membrane-associated pool, suggesting that membrane anchoring occurs by palmitoylation. Palmitoylation (the addition of a 16-carbon saturated fatty acid) is a reversible posttranslational modification on cysteine residues and is considered a rapid and dynamic regulatory event on many signal transduction proteins, including G-protein α subunits (reviewed in ref. 42); it enhances membrane binding and can also alter protein activity. GAIP lacks other features involved in membrane anchoring, such as a transmembrane domain, a consensus N-myristoylation site at its N terminus, a CaaX prenylation box found at the C terminus of several small GTP-binding proteins (43), or a polyleucine C terminus (44). Our finding that the membrane-associated pool of GAIP is palmitoylated suggests two possibilities whereby palmitoylation might regulate its activity, (i) by targeting a part of the cytosolic pool of GAIP to the membrane and/or (ii) by directly modifying the conformation of GAIP and hence its activity.
Although the exact function of cysteine string proteins is still unclear, they have been detected on synaptic vesicles in brain (37) and on pancreatic zymogen granules (45) and chromaffin granules (46) and are thought to be involved in exocytosis and/or membrane fusion (47). It has been shown that cysteine string proteins undergo extensive palmitoylation—on up to 11 cysteine residues (39). The increased incorporation of [3H]palmitate into GAIP in comparison to Gαs, which undergoes palmitoylation on only one site (41), is a strong indicator that GAIP is palmitoylated at more than one site, probably on its cysteine string motif. Although the increased 3H signal could also be explained by a faster turnover of palmitoylation on GAIP, the short labeling period (1 hr) with [3H]palmitic acid renders this possibility unlikely. It is not yet known whether palmitoylated GAIP activates or inactivates the protein in its interaction with Gαi3. The activity of GAP-43, a protein that increases GTP binding of the Gαo subunit (48), is regulated by palmitoylation and gives rise to an inactive, membrane-associated pool of the protein (49). The behavior of specific point mutants of GAIP in and around the cysteine string motif is likely to provide answers to some of these questions.
Previously we have shown that GAIP specifically interacts with Gαi3. In this study we extended our survey by analyzing GAIP’s interaction with members of all four subclasses of Gα subunits (24). We found that GAIP interacts specifically with members of the Gαi subfamily in the yeast two-hybrid assay, while the other subclasses—i.e., Gαs, Gαq, and Gα12/13—do not. The fact that neither wild-type Gαs nor its activated form Gαs(Q227L) (25, 50) gave a positive result in this assay suggests that adoption of an activated conformation by any G protein is not sufficient for interaction, and other determinants must be involved.
We found that the C terminus of Gαi subunits plays an important role in the binding to GAIP. Both a C-terminal deletion mutant Gαi3(Δ345–354) and a point mutant Gαi3(G352N) show reduced—but not abolished—interaction with GAIP. The importance of the C-terminal region of the Gαi subunits is again indicated by the strong GAIP/Gpa1 interaction, because Gpa1 is most homologous to the Gαi subfamily in the C-terminal region (30). However, the C terminus of Gαi3 is not sufficient for maximal interaction with GAIP, because the Gαq/i3(345–354) chimera gave a negative result in our assay. Regions other than the C terminus must also be involved in the binding to GAIP, because Gαi1 interacts much more strongly than Gαi2, although both have identical C-terminal decapeptide sequences. The C-terminal region of Gα subunits has also been demonstrated to be important for the specificity of receptor–Gα interactions (27, 29), and, interestingly, this region is disordered in crystals of Gαt (51), suggesting structural mobility of that region. Perhaps by competing for the same domain on Gαi, GAIP might be able to interrupt contacts between Gαi subunits and serpentine receptors. Interruption of receptor–G-protein interaction has already been described for the rhodopsin–rhodopsin kinase–arrestin complex in visual light transduction and for the β2-adrenergic receptor system. In the latter systems the targets for interruption are the receptors themselves, not the Gα subunits (52).
Our results show that GAIP interacts preferentially with the activated (GTP-bound) form of Gαi3. Recent reports show that GAIP has GTPase-activating activity on Gαi1 and other members of the Gαi subfamily, indicating GAIP is a GAP (7–9). Though not obtained by a direct GTPase assay, our data from in vivo and in vitro assays are consistent with these observations. They are also consistent with the hypothesis that GAIP is a downstream target of Gαi3, much as rasGAP has been suggested to be a downstream effector of ras (53, 54).
GAIP is a composite molecule containing an RGS domain and a cysteine string motif, suggesting that it could display more than one function. Its RGS domain binds Gαi subunits and enhances their GTPase activity. The presence of a cysteine string motif, also found in cysteine string proteins localized to synaptic vesicles (37), suggests that GAIP might be involved in membrane trafficking. Although GAIP has a cysteine string motif, it cannot be considered a cysteine string protein because it lacks the dnaJ domain—a putative HSP70 interacting domain—present in all cysteine string proteins isolated so far (55). The recent findings that cysteine string proteins are expressed outside the brain (56), are found on zymogen granule membranes in the pancreas (45), and are associated with chromaffin granules (46) suggest a more general involvement of this class of proteins in exocytosis. Localization of GAIP to specific membranes will be required to provide clues as to which step(s) of the trafficking process, if any, GAIP is involved.
Acknowledgments
We thank Sheryl Denker for subcloning Gαs and the Gαs(Q227L) mutant into the pGBT9 bait vector, Thierry Fischer and Joe Noel for helpful discussions, and Michael McCaffery for assistance with the preparation of the figures. This research was supported by Grants CA58689 and DK17780 from the National Institutes of Health (to M.G.F.). E.E. is a graduate student in the Biomedical Sciences Graduate program and was supported (1995–1996) by National Institutes of Health Training Grant GM07752. L.H. is a recipient of a National Institutes of Health National Research Service Award (CA66289).
Footnotes
Abbreviations: β-gal, β-galactosidase; GAP, GTPase-activating protein; GAIP, Gα-interacting protein; GST, glutathione S-transferase; GTP[γS], guanosine 5′-[γ-thio]triphosphate; HA, hemagglutinin protein of influenza virus; RGS, regulator of G-protein signaling.
References
- 1.De Vries L, Mousli M, Wurmser A, Farquhar M G. Proc Natl Acad Sci USA. 1995;92:11916–11920. doi: 10.1073/pnas.92.25.11916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Druey K M, Blumer K J, Kang V H, Kehrl J H. Nature (London) 1996;379:742–746. doi: 10.1038/379742a0. [DOI] [PubMed] [Google Scholar]
- 3.Koelle M R, Horvitz H R. Cell. 1996;84:115–125. doi: 10.1016/s0092-8674(00)80998-8. [DOI] [PubMed] [Google Scholar]
- 4.Roush W. Science. 1996;271:1056–1058. doi: 10.1126/science.271.5252.1056. [DOI] [PubMed] [Google Scholar]
- 5.Siderovski D P, Hessel A, Chung S, Mak T W, Tyers M. Curr Biol. 1996;6:211–212. doi: 10.1016/s0960-9822(02)00454-2. [DOI] [PubMed] [Google Scholar]
- 6.Dohlman H G, Apaniesk D, Chen Y, Song J, Nusskern D. Mol Cell Biol. 1995;15:3635–3643. doi: 10.1128/mcb.15.7.3635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Berman D M, Wilkie T M, Gilman A G. Cell. 1996;86:445–452. doi: 10.1016/s0092-8674(00)80117-8. [DOI] [PubMed] [Google Scholar]
- 8.Watson N, Linder M E, Druey K M, Kehrl J H, Blumer K J. Nature (London) 1996;383:172–175. doi: 10.1038/383172a0. [DOI] [PubMed] [Google Scholar]
- 9.Hunt T W, Fields T A, Casey P J, Peralta E G. Nature (London) 1996;383:175–177. doi: 10.1038/383175a0. [DOI] [PubMed] [Google Scholar]
- 10.Boguski M S, McCormick F. Nature (London) 1993;366:643–653. doi: 10.1038/366643a0. [DOI] [PubMed] [Google Scholar]
- 11.Wang Y, Boguski M, Riggs M, Rodgers L, Wigler M. Cell Regul. 1991;2:453–465. doi: 10.1091/mbc.2.6.453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Landis C A, Masters S B, Spada A, Pace A M, Bourne H R, Vallar L. Nature (London) 1989;340:692–696. doi: 10.1038/340692a0. [DOI] [PubMed] [Google Scholar]
- 13.Ausubel F M. In: Current Protocols in Molecular Biology. Ausubel F M, Brent R, Kingston R E, Moore D M, Seidman J G, Smith J A, Struhl K, editors. Vol. 1. New York: Wiley; 1990. pp. 9.2.1–9.2.6. [Google Scholar]
- 14.Wada I, Rindress D, Cameron P H, Ou W J, Doherty J J, Louvard D, Bell A W, Dignard D, Thomas D Y, Bergeron J J M. J Biol Chem. 1991;266:19599–19610. [PubMed] [Google Scholar]
- 15.Sherer P E, Lederkremer G Z, Williams S, Fogliano M, Baldini G, Lodish H F. J Cell Biol. 1996;133:257–268. doi: 10.1083/jcb.133.2.257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hendricks L C, McCaffery M, Palade G, Farquhar M G. Mol Biol Cell. 1993;4:413–424. doi: 10.1091/mbc.4.4.413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Duden R, Griffiths G, Frank R, Argos P, Kreis T E. Cell. 1991;64:649–655. doi: 10.1016/0092-8674(91)90248-w. [DOI] [PubMed] [Google Scholar]
- 18.Fujiki Y, Hubbard A L, Fowler S, Lazarow P B. J Cell Biol. 1982;93:97–102. doi: 10.1083/jcb.93.1.97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Laemmli U K. Nature (London) 1970;227:680–685. doi: 10.1038/227680a0. [DOI] [PubMed] [Google Scholar]
- 20.Schiestl R H, Gietz R D. Curr Genet. 1989;16:339–346. doi: 10.1007/BF00340712. [DOI] [PubMed] [Google Scholar]
- 21.Altschul S F, Gish W, Miller W, Myers W W, Lipman D J. J Mol Biol. 1990;215:403–410. doi: 10.1016/S0022-2836(05)80360-2. [DOI] [PubMed] [Google Scholar]
- 22.Chien C T, Bartel P L, Sternglanz R, Fields S. Proc Natl Acad Sci USA. 1991;88:9578–9582. doi: 10.1073/pnas.88.21.9578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Miller J H. Experiments in Molecular Genetics. Plainview, NY: Cold Spring Harbor Lab. Press; 1972. pp. 352–355. [Google Scholar]
- 24.Simon M I, Strathmann M P, Gautam N. Science. 1991;252:802–808. doi: 10.1126/science.1902986. [DOI] [PubMed] [Google Scholar]
- 25.Graziano M P, Gilman A G. J Biol Chem. 1989;264:15475–15482. [PubMed] [Google Scholar]
- 26.West R E J, Moss J, Vaughan M, Liu T, Liu T Y. J Biol Chem. 1985;260:14428–14430. [PubMed] [Google Scholar]
- 27.Conklin B R, Farfel Z, Lustig K D, Julius D, Bourne H. Nature (London) 1993;363:274–276. doi: 10.1038/363274a0. [DOI] [PubMed] [Google Scholar]
- 28.Dratz E A, Furstenau J E, Lambert C G, Thireault D L, Rarick H, Schepers T, Pakhlevaniants S, Hamm H E. Nature (London) 1993;363:276–281. doi: 10.1038/363276a0. [DOI] [PubMed] [Google Scholar]
- 29.Hamm H E, Gilchrist A. Curr Opin Cell Biol. 1996;8:189–196. doi: 10.1016/s0955-0674(96)80065-2. [DOI] [PubMed] [Google Scholar]
- 30.Dietzel C, Kurjan J. Cell. 1987;50:1001–1010. doi: 10.1016/0092-8674(87)90166-8. [DOI] [PubMed] [Google Scholar]
- 31.Hermouet S, de Mazancourt P, Spiegel A M. Cell Signalling. 1993;5:215–225. doi: 10.1016/0898-6568(93)90072-t. [DOI] [PubMed] [Google Scholar]
- 32.Hermouet S, Merendino J J, Gutkind J S, Spiegel A M. Proc Natl Acad Sci USA. 1991;88:10455–10459. doi: 10.1073/pnas.88.23.10455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Coleman D E, Berghuis A M, Lee E, Linder M E, Gilman A G, Sprang S R. Science. 1994;265:1405–1412. doi: 10.1126/science.8073283. [DOI] [PubMed] [Google Scholar]
- 34.Sondek J, Lambright D J, Noel J P, Hamm H E, Sigler P B. Nature (London) 1994;372:276–279. doi: 10.1038/372276a0. [DOI] [PubMed] [Google Scholar]
- 35.Clark J D, Lin L-L, Kriz R W, Ramesha C S, Sultzman L A, Lin A Y, Milona N, Knopf J L. Cell. 1991;65:1043–1051. doi: 10.1016/0092-8674(91)90556-e. [DOI] [PubMed] [Google Scholar]
- 36.Gundersen C B, Umbach J A. Neuron. 1992;9:527–537. doi: 10.1016/0896-6273(92)90190-o. [DOI] [PubMed] [Google Scholar]
- 37.Mastrogiacomo A, Parsons S M, Zampighi G A, Jenden D J, Umbach J A, Gundersen C B. Science. 1994;263:981–982. doi: 10.1126/science.7906056. [DOI] [PubMed] [Google Scholar]
- 38.Zinsmaier K E, Hofbauer A, Heimbeck G, Pflugfelder G O, Buchner S, Buchner E. J Neurogenet. 1990;7:15–29. doi: 10.3109/01677069009084150. [DOI] [PubMed] [Google Scholar]
- 39.Gundersen C B, Mastrogiacomo A, Faull K, Umbach J U. J Biol Chem. 1994;269:19197–19199. [PubMed] [Google Scholar]
- 40.Mastrogiacomo A, Gundersen C B. Mol Brain Res. 1995;28:12–18. doi: 10.1016/0169-328x(94)00172-b. [DOI] [PubMed] [Google Scholar]
- 41.Degtyarev M Y, Spiegel A M, Jones T L Z. Biochemistry. 1993;32:8057–8061. doi: 10.1021/bi00083a001. [DOI] [PubMed] [Google Scholar]
- 42.Milligan G, Parenti M, Magee A I. Trends Biochem Sci. 1995;20:181–186. doi: 10.1016/s0968-0004(00)89004-0. [DOI] [PubMed] [Google Scholar]
- 43.Magee T. Trends Cell Biol. 1992;2:318–323. doi: 10.1016/0962-8924(92)90172-j. [DOI] [PubMed] [Google Scholar]
- 44.Whitley P, Grahn E, Kutay U, Rapoport T A, von Heijne G. J Biol Chem. 1996;271:7583–7586. doi: 10.1074/jbc.271.13.7583. [DOI] [PubMed] [Google Scholar]
- 45.Braun J E, Scheller R H. Neuropharmacology. 1995;34:1361–1369. doi: 10.1016/0028-3908(95)00114-l. [DOI] [PubMed] [Google Scholar]
- 46.Chamberlain L H, Henry J, Burgoyne R D. J Biol Chem. 1996;271:19514–19517. doi: 10.1074/jbc.271.32.19514. [DOI] [PubMed] [Google Scholar]
- 47.Gundersen C B, Mastrogiacomo A, Umbach J A. J Theor Biol. 1995;172:269–277. doi: 10.1006/jtbi.1995.0023. [DOI] [PubMed] [Google Scholar]
- 48.Strittmatter S M, Valenzuela D, Kennedy T E, Neer E J, Fishman M C. Nature (London) 1990;344:836–841. doi: 10.1038/344836a0. [DOI] [PubMed] [Google Scholar]
- 49.Sudo Y, Valenzuela D, Beck-Sickinger A G, Fishman M C, Strittmatter S M. EMBO J. 1992;11:2095–2102. doi: 10.1002/j.1460-2075.1992.tb05268.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Masters S B, Miller R T, Chi M H, Chang F H, Beiderman B, Lopez N G, Bourne H R. J Biol Chem. 1989;264:15467–15474. [PubMed] [Google Scholar]
- 51.Noel J P, Hamm H E, Sigler P B. Nature (London) 1993;366:654–663. doi: 10.1038/366654a0. [DOI] [PubMed] [Google Scholar]
- 52.Lefkowitz R J, Inglese J, Koch W J, Pitcher J, Attramadal H, Caron M G. Cold Spring Harbor Symp Quant Biol. 1992;62:127–133. doi: 10.1101/sqb.1992.057.01.016. [DOI] [PubMed] [Google Scholar]
- 53.Martin G A, Yatani A, Clark R, Conroy L, Polakis P, Brown A M, McCormick F. Science. 1992;255:192–194. doi: 10.1126/science.1553544. [DOI] [PubMed] [Google Scholar]
- 54.Duchesne M, Schweighoffer F, Parker F, Clerc F, Thang M N, Tocque B. Science. 1993;258:525–528. doi: 10.1126/science.7678707. [DOI] [PubMed] [Google Scholar]
- 55.Caplan A J, Cyr D M, Douglas M G. Mol Biol Cell. 1993;4:555–563. doi: 10.1091/mbc.4.6.555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Chamberlain L H, Burgoyne R D. J Biol Chem. 1996;271:7320–7323. doi: 10.1074/jbc.271.13.7320. [DOI] [PubMed] [Google Scholar]