

Abstract
Previously we have shown that transient attention—the more automatic, stimulus-driven component of spatial attention—enhances spatial resolution. Specifically, transient attention improves texture segmentation at the periphery, where spatial resolution is too low, but impairs performance at central locations, where spatial resolution is already too high for the task. In the present study we investigated whether sustained attention—the more controlled component of spatial attention—can also affect texture segmentation, and if so whether its effect will be similar to that of transient attention. To that end we combined central, symbolic cues with texture displays in which the target appears at several eccentricities. We found that sustained attention can also affect texture segmentation, but unlike transient attention, sustained attention improved performance at all eccentricities. Comparing the effect of pre-cues and post-cues indicated that the benefit brought about by sustained attention is significantly greater than the effect of location uncertainty reduction. These findings indicate that sustained attention is a more flexible mechanism that can optimize performance at all eccentricities in a task where performance is constrained by spatial resolution.
Keywords: Covert attention, Sustained attention, Texture segmentation, Spatial vision
1. Introduction
Visual attention allows us to select part of the otherwise overwhelming amount of information in our visual field and process it in a privileged way. One way in which this attentional selection occurs is based on the spatial location of visual information. Typically, we foveate the location in space to which we wish to attend, but it is well known that we can direct our attention to a region in our visual field without moving our eyes towards this location (e.g., Eriksen & Hoffman, 1972; Posner, 1980). The selection of information based on its spatial location in the absence of eye movements is referred to as spatial covert attention (Posner, 1980). There are two components of spatial attention, ‘sustained’ and ‘transient’. ‘Sustained attention’ is a conceptually-driven component that requires conscious effort and is activated in about 300 ms. ‘Transient attention’ is a stimulus-driven component, and it is activated in an automatic manner in about 100 ms (e.g., Cheal & Lyon, 1991; Jonides, 1981; Müller & Rabbitt, 1989; Nakayama & Mackeben, 1989; Posner, 1980; Remington, Johnston, & Yantis, 1992). Transient attention is considered to operate at an earlier stage of visual cortical processing than the sustained component (e.g., Nakayama & Mackeben, 1989). Indeed, neurophysiological studies suggest that whereas sustained attention is cortical in nature, transient attention is mediated by both cortical and subcortical networks (Corbetta & Shulman, 2002; Kastner & Ungerleider, 2000). Previously, we have documented the effects of transient attention on a texture segmentation task constrained by spatial resolution (e.g., Carrasco, Loula, & Ho, 2006; Yeshurun & Carrasco, 1998; Yeshurun & Carrasco, 2000). In this study, we investigate the effects of sustained attention on such a texture segmentation task.
A common method to directly manipulate the allocation of spatial attention employs attentional cues, which precede target presentation and induce observers to direct their attention to the target location. Different types of attentional cues are used to manipulate transient and sustained attention: The former is manipulated by a peripheral cue presented adjacent to the target location, and the latter is manipulated by a central cue presented in the center of the visual field. Peripheral cues induce a more rapid shift of attention than central cues, and usually produce larger attentional effects at short intervals. The effects of peripheral cues peak at ∼100 ms but then decay rapidly whereas the effects of central cues peak at ∼300 ms and then remain for a longer duration (e.g., Carrasco, Ling, & Read, 2004; Cheal & Lyon, 1991; Jonides, 1981; Ling & Carrasco, 2006; Müller & Findlay, 1988; Müller & Rabbitt, 1989; Nakayama & Mackeben, 1989). In addition, whereas the shifts of attention by central cues appear to be under conscious control, the effects of peripheral cues are involuntary (but see Ristic, Friesen, & Kingstone, 2002; Tipples 2002, who have shown that there may be a reflexive component for faces and arrows presented at central locations). For instance, peripheral cues cannot be voluntarily interrupted or ignored, even if observers are instructed to do so (Jonides, 1981), and their effect is little influenced by cue validity (i.e., the probability with which the cue predicts the target location). Moreover, shifts of attention induced by peripheral cues have been shown to occur even if the cue is uninformative (e.g., Jonides, 1981; Müller & Rabbitt, 1989 Pestilli & Carrasco, 2005; Pestilli, Viera, & Carrasco, 2007), or if the cue actually impairs performance due to its low validity (e.g., Giordano, McElree, & Carrasco, 2004). In contrast, effects of central cues depend on cue validity and might not occur when the central cues are uninformative (e.g., Giordano et al., 2004; Jonides, 1981; Kinchla, 1969; Kinchla, 1980; Sperling & Melchner, 1978; but see Gibson & Bryant, 2005).
Previously, we have proposed that transient attention can enhance spatial resolution, thus allowing us to resolve finer details at the attended location (‘resolution hypothesis'). Several studies have provided evidence supporting the hypothesis that transient attention can enhance spatial resolution. For example, directing transient attention to the target location improves performance in both acuity and hyper-acuity tasks even when a supra-threshold target is presented without distracters (Carrasco, Williams, & Yeshurun, 2002; Golla, Ignashchenkova, Haarmeier, & Their, 2004; Yeshurun & Carrasco, 1999). Similarly, the decrement in visual search performance that occurs as the target is presented at farther peripheral locations is significantly reduced when transient attention is directed to the target location. This finding suggests that attention can reduce resolution differences between the fovea and the periphery (Carrasco & Yeshurun, 1998), much like the effect obtained when the elements of a search display are cortically magnified (Carrasco & Frieder, 1997). Additionally, the hypothesis that transient attention enhances resolution is consistent with neurophysiological studies demonstrating that a neuron's response to its preferred stimulus is greatly reduced when the preferred stimulus is not attended, and an attended, non-preferred stimulus is also presented within the neuron's receptive field. These findings suggest that attention contracts the cell's receptive field around the attended stimulus (e.g., Desimone & Duncan, 1995; Luck, Chelazzi, Hillyard, & Desimone, 1997; Moran & Desimone, 1985; Reynolds & Chelazzi, 2004; Reynolds & Desimone, 1999; Womelsdorf, Anton-Erxleben, Pieper, & Treue, 2006).
In another study we further tested the ‘resolution hypothesis’ by exploring the effects of transient attention on performance in a basic texture segmentation task (Yeshurun & Carrasco, 1998). We found that transient attention can affect texture segmentation, and that the pattern of this attentional effect depends on the spatial resolution at the attended location. Specifically, we found that transient attention improves performance at locations in which performance is limited by a spatial resolution that is too low (i.e., at peripheral locations), but impairs performance at locations in which performance is limited by a spatial resolution that is too high (i.e., at central locations). The current study was designed to explore whether sustained attention can also affect texture segmentation, and whether the pattern of such an effect would be similar to that of transient attention.
The texture segmentation task we employed in our transient attention study required the detection of a target patch composed of oblique line elements embedded in a larger background of orthogonally oriented line elements (Yeshurun & Carrasco, 1998; Fig. 1). Performance in this task peaks when the target appears at mid-peripheral locations, and drops as the target appears at more central or farther peripheral locations (e.g., Gurnsey, Pearson, & Day, 1996; Joffe & Scialfa, 1995; Kehrer, 1989; Kehrer, 1997; Meinecke & Kehrer, 1994; Morikawa 2000; Potechin & Gurnsey, 2003). It has been hypothesized that performance is suboptimal when there is a mismatch between the specific scale of the texture and the size of spatial linear filters. These linear filters are thought to underlie the visual processing of textures (e.g., Bergen & Landy, 1991; Caelli, 1985; Landy & Bergen, 1991; Sutter, Beck, & Graham, 1989; See Landy & Graham, 2004 for a review), and a large body of evidence indicates that they are tuned to a specific band of spatial frequency and orientation and that their average size increases as a function of eccentricity (e.g., Rovamo & Virsu, 1979; for a review, De Valois & De Valois, 1988; Graham, 1989; Wilson, Levi, Maffei, Rovamo, & De Valois 1990). According to this view, the optimal average filter size for the segmentation of the given texture is available at mid-peripheral locations, where performance peaks. At the fovea, where resolution is too high for the scale of the texture, performance drops because the filters are too small. At the far periphery, where resolution is too low, performance drops because the filters are too large (Gurnsey et al., 1996).
Fig. 1.

A schematic example of the texture displays used in Experiments 1 and 2. The texture line elements are oriented ±45° from vertical and the texture display extends along the horizontal meridian. A 3 × 3 target patch is visible in the display on the right.
In light of the resolution hypothesis, Yeshurun and Carrasco (1998) hypothesized that if attention indeed enhances spatial resolution, attending to the target location should enhance performance at the periphery, but should impair performance at the fovea. To test this prediction we combined peripheral cues with this texture segmentation task. On the cued trials, a small peripheral cue indicated the target location prior to its appearance. Such a peripheral cue is considered to capture attention in a stimulus-driven, “automatic” manner (e.g., Jonides, 1981; Müller & Rabbitt, 1989; Pestilli & Carrasco, 2005; Posner, 1980; Yantis, 1996). On the neutral trials, a pair of lines, appearing above and below the entire display, indicated that the target was equally likely to appear at any location. As predicted, accuracy was higher for the cued than the neutral trials at the more peripheral locations, but was lower at central locations resulting in a central attentional impairment. This pattern of results, and especially the central attentional impairment, is predicted only by the resolution hypothesis, and therefore it lends strong support to the hypothesis that transient attention enhances the spatial resolution at the attended location (Yeshurun & Carrasco, 1998). Any other model of attention, like those based on uncertainty or noise reduction, would predict a benefit throughout all eccentricities. These findings have been replicated by several studies demonstrating that these attentional effects on texture segmentation, considered to be pre-attentive (e.g., Chubb & Talevich, 2002; Julesz, 1981), are robust and can be generalized to textures of a very different nature. For instance, similar results have been obtained in a study in which the texture was presented along the vertical rather than horizontal meridian (Talgar & Carrasco, 2002); when the peripheral cue was a frame around the central element in the texture target (Yeshurun & Carrasco, 2008); when observers had to indicate the orientation of the target-texture after selectively adapting to different spatial frequencies (Carrasco et al., 2006); and when the first- and second-order content of the textures was manipulated (Yeshurun & Carrasco, 2000).
The studies described thus far have documented the effect of transient attention on this texture segmentation task. In this study we explored the nature of the more controlled attentional component—sustained attention. Specifically, we were interested in the following questions: First, we asked whether attentional effects on texture segmentation will be found when attention is manipulated via a central cue ensuring the activation of sustained attention. Given that sustained attention seems to operate at a later stage of visual processing than transient attention (e.g., Corbetta & Shulman, 2002; Kastner & Ungerleider, 2000; Liu, Stevens, & Carrasco, 2007; Nakayama & Mackeben, 1989), and that texture segmentation appears to take place at a relatively early stage of visual processing (e.g., Lamme, 1995; Lamme, Van Dijk, & Spekreijse, 1993; Purpura, Victor, & Katz, 1994), sustained attention may not be able to affect performance in the basic texture segmentation task employed here. Second, if effects of sustained attention on texture segmentation were found, would these effects be similar to the effects of transient attention on texture segmentation? That is, would both an attentional benefit and an attentional impairment emerge? Can the sustained attentional component, like the transient one, affect spatial resolution at the attended location?
On the one hand, sustained attention may always increase spatial resolution regardless of the consequences of such enhancement (i.e., performance increment at peripheral locations and decrement at central locations), as does transient attention. On the other hand, it is possible that sustained attention, a more flexible mechanism, could optimize performance across all eccentricities. The latter outcome is expected if sustained attention affects texture segmentation by adopting the best resolution at a given eccentricity, or by improving the signal to noise ratio at all eccentricities. Finally, if texture segmentation is impenetrable to sustained attention, performance in those tasks will not vary as a function of cueing conditions.
To investigate these issues, we manipulated either sustained attention (Experiment 1) via central cues, or transient attention (Experiment 2) via peripheral cues maintaining task and stimuli identical. In these two experiments the texture segmentation task was the same as the one employed with transient attention in previous studies (Talgar & Carrasco, 2002; Yeshurun & Carrasco, 1998). In Experiments 3 and 4 the texture was modified from a homogeneous background to a heterogeneous background to ensure that performance is limited only by spatial factors. Finally, in Experiment 5 we evaluated the contribution of location uncertainty at the decisional level to the effect of sustained attention.
2. Experiment 1
The aims of this experiment were twofold: To test (1) whether sustained attention can also affect observers' performance in texture segmentation; (2) whether the attentional effect differs as a function of target eccentricity. We employed texture displays similar to the ones used with transient attention (Yeshurun & Carrasco, 1998; Fig. 1) and a two interval forced-choice task (2IFC; Fig. 2). A trial consisted of two temporal intervals, each containing a texture display preceded by a cue. On the cued trials, a central, symbolic cue (a digit and a line) provided the exact location of the target with 100% validity. On the neutral trials, the cue did not provide information regarding the target location. The target appeared equally often in each temporal interval and at one of 9 possible locations. Observers were instructed to report whether the target was present in the first or the second interval.
Fig. 2.

Schema of the frame sequence in a typical trial of Experiments 1 (sustained attention) and 2 (transient attention) in a 2IFC task. In this example, the target is present in the second interval. The cue location (sustained: central; transient: peripheral) and the timing (sustained cue duration: 200 ms, ISI: 600 ms; transient cue duration: 47 ms, ISI: 47 ms) differ as they were set to maximize the effects of sustained and transient attention, respectively.
Precueing effects have been ascribed to changes in decisional, rather than sensory, factors. According to this view, precueing effects would arise from the fact that observers are encouraged to adopt a more liberal criterion or to assign more weight to visual information extracted from the cued location (Kinchla, 1980; Kinchla, Chen, & Evert, 1995; Palmer, 1994; Shaw, 1984). However, this can only be the case when the adopted precueing paradigm implies that one of the responses is more probable and thus conveys information about which response is more likely to be correct on a given trial. In the present study, however, in each trial the precues always appeared both in the target interval and in the non-target interval. They did therefore not convey any information about which response would be correct on a given trial. Moreover, if observers were to assign more weight to the information extracted from the cued location, they would do so for the cued locations of both intervals. As discussed in previous papers (Carrasco et al., 2006; Talgar & Carrasco, 2002; Yeshurun & Carrasco, 1998; Yeshurun & Carrasco, 2000), given that the precue appeared in both temporal intervals, it did not associate higher probability with one of the responses (target in first or second interval), and observers could not rely on its presence to reach a decision about target presence.
2.1. Methods
2.1.1. Observers
Sixteen observers from NYU participated in Experiment 1 (14 from the Introductory Psychology Subject Pool). All observers had normal or corrected-to-normal vision.
2.1.2. Apparatus
Stimuli were presented on a 21” monitor attached to a PowerMac G4 computer. Data collection and stimulus presentation were controlled using PsyScope for OS9 (Cohen, MacWhinney, Flatt, & Provost, 1993). The monitor resolution was set to 1280 × 1024 pixels at a frame rate of 85 Hz. A chinrest was used to stabilize the head and to maintain a viewing distance of 57 cm. Observers' eye position was recorded using an iSCAN infrared camera pointed at one of the eyes.1 The experiment was conducted in a dark and quiet room.
2.1.3. Stimuli
The textures were composed of black line elements (each subtending 0.1 × 0.7°, oriented at ±45° from vertical) presented on a white background. The luminance of the line elements was about 0.01 cd/m2 and that of the background was about 92 cd/m2. The luminance of the texture display was about 79 cd/m2. Each texture display consisted of 287 line elements arranged on a virtual grid of 7 rows × 41 columns, subtending 5 × 28°. The line elements were jittered by 0.3°. In each trial, one of the two texture displays contained a target patch of 2 × 2°, consisting of 3 × 3 line elements, whose orientation was orthogonal to the lines of the background texture. The target patch appeared in either of the two temporal intervals with 0.5 probability, and was presented at one of 9 possible locations within the texture-background along the horizontal meridian, corresponding to 5 eccentricities (one at the center and 4 to each side of fixation: 0°, 1.5°, 5°, 7.5°, and 11.5°). The mask-texture was composed of 287 black ‘X’ elements (7 rows × 41 columns).
On the cued trials the cue provided the exact location of the target. It was composed of a gray digit (“0” to “4”), presented at the center of the screen (within a region of 0.5 × 0.8°), indicating the eccentricity at which the target may appear, and a small horizontal line (0.1 × 0.5 ), which appeared to the right or left side of the digit (unless it was “0”, denoting the central location), indicating the hemifield in which the target may appear. The cue was 100% valid— it provided the exact location of the target, for the interval in which it was present. In the interval without a target the cue was equally likely to indicate any other location. Thus, in the two intervals of each trial the sustained cues indicated two different locations.
On the neutral trials, the cue was composed of two long, gray horizontal lines (0.2 × 28 ) appearing 0.5° above and below the entire display. This cue indicated that the target, if present in that interval, was equally likely to appear at any of the 9 possible locations. Both cues provided information regarding display onset. Trials were either neutral or cued. Thus, even if observers were to adopt a different decisional criterion for the different cueing conditions, there was no reason for observers to assign different weights or adopt different criteria across the two intervals of a given trial. The described characteristics of the precueing paradigm adopted here guarantee that the precueing effects can be ascribed to sensory rather than decisional factors.
2.1.4. Procedure
Each trial of the 2IFC task consisted of two temporal intervals, each containing a texture display preceded by a cue. Each temporal interval began with a central fixation dot (Fig. 2). 500-ms after the onset of the fixation dot the cue was presented for 200-ms and was followed by an ISI of 600-ms. Half the trials were cued trials and half were neutral trials. After the ISI, the texture was displayed for a median of 30-ms (range 20–50 ms). The duration of the texture display was adjusted for each observer during a set of practice trials to achieve an overall performance level of about 75% correct. A 200-ms mask was presented immediately following the texture display. The observers were instructed to keep their eyes fixated at the center, and were asked to indicate the interval (first or second) whose texture display contained a target, by pressing the ‘1’ or ‘2’ key of the keyboard. Given that the time between the onset of the cue and the offset of the texture display was longer than the time estimated to complete a saccade (Mayfrank, Kimmig, & Fischer, 1987), eye position was monitored to ensure that observers did not break fixation.
In the cued trials, observers had to associate the symbolic cues to a given location on the screen. To do so, at the beginning of each block of trials, a display was shown in which each digit was placed at the corresponding location it would cue. Moreover, observers learned this association during the practice (120 trials) preceding the actual data collection (160 trials per observer). To obtain the same amount of measurements per eccentricity we collected twice as many observations from the central location.
2.2. Results and discussion
To evaluate the effects of cueing condition (neutral vs. cued) as a function of target eccentricity in this texture segmentation task, we conducted a two-way repeated-measures ANOVA on the accuracy data.2 As can be seen in Fig. 3, there was a significant main effect of target eccentricity: accuracy was highest at mid periphery and dropped at more central or farther peripheral locations [F(4, 60) = 9.85, p < 0.001, g=hp 2 = 0.396]. This pattern of results as a function of eccentricity is consistent with previous studies documenting the central performance drop (e.g., Gurnsey et al., 1996; Joffe & Scialfa, 1995; Kehrer, 1989; Morikawa 2000; Talgar & Carrasco, 2002; Yeshurun & Carrasco, 1998). Most important for the purpose of this study, however, is the finding that whereas there was a significant main effect of cueing—accuracy was significantly higher in the cued than neutral condition [F(1, 15) = 13.47, p < 0.005, g=hp 2 = 0.473] —there was no significant interaction of cueing and eccentricity (F < 1). Analysis of reaction time (RT) confirmed that there were no speed-accuracy tradeoffs. The main effect of eccentricity was significant [F(4,60) = 3.219, p < 0.05, ηp 2 = 0.177]: Reaction times were shorter at mid-peripheral locations, compared with more central and more peripheral locations. This is consistent with previous studies reporting the central performance drop (e.g., Gurnsey et al., 1996). The main effect of cueing [F < 1], and its interaction with cue type [F <1] were not significant.
Fig. 3.
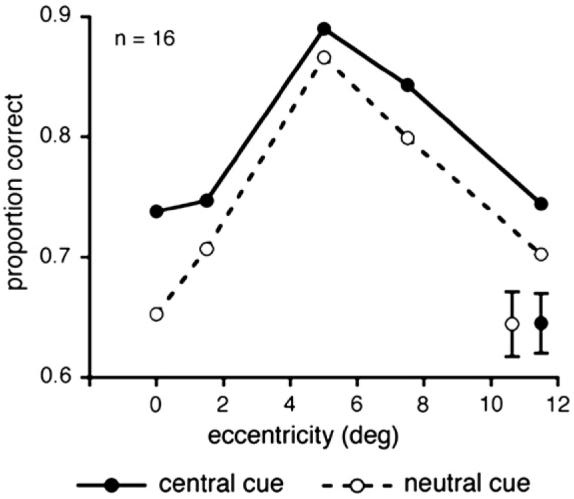
Average proportion of correct responses as a function of target eccentricity and cue type (central cue vs. neutral cue) in Experiment 1. In the neutral condition (open circles, dashed line), performance peaks at mid-periphery (5° of eccentricity) and drops towards more central and more peripheral locations, replicating the central performance drop. Precueing the target location with a central cue (filled circles, continuous line) improved performance across all eccentricities. Error bars are indicated in the lower right of the graph and represent ± 1s SEM per cueing condition, averaged across eccentricities.
This pattern of results indicates that sustained attention, like transient attention, can affect performance in texture segmentation task. However, as opposed to transient attention, precueing the target location with a central cue aided performance across all eccentricities—there was no central attentional impairment. The finding that transient attention impairs texture segmentation at central locations was found in several studies employing different tasks and textures (Carrasco et al. 2006; Talgar & Carrasco, 2002; Yeshurun & Carrasco, 1998, 2000, 2008). Nevertheless, we conducted Experiment 2 to ensure that a central attentional impairment is indeed found with transient attention under the exact experimental conditions used in Experiment 1.
3. Experiment 2
Experiment 2 was designed to test directly whether the central attentional impairment, found with transient attention in previous studies (Carrasco et al. 2006; Talgar & Carrasco, 2002; Yeshurun & Carrasco, 1998, 2000, 2008), would also be found under the exact experimental conditions used in Experiment 1. This is particularly important because in Experiment 1 there were fewer possible target eccentricities than in the previous studies, which allowed observers to easily associate each cue digit to a specific target location. Thus, the goal of this experiment was to ensure that the differential attentional effect on texture segmentation found for transient and sustained attention was not simply due to this difference. To that end, this experiment was identical to Experiment 1 apart for the employment of a peripheral cue, rather than a central cue, to selectively manipulate transient attention, and the shortening of the time between cue onset and target onset to 94-ms to optimize the effect of transient attention.
3.1. Methods
3.1.1. Observers
Eighteen observers from the University of Haifa participated in this experiment. All observers had normal or corrected-to-normal vision.
3.1.2. Apparatus, stimuli and procedure
The apparatus, stimuli and procedure were the same as those used in Experiment 1 except for the following: Instead of the digit, the attentional cue was composed of a 0.3 × 0.6° green horizontal bar appearing 0.3° above the target location on the target interval, and above another, randomly chosen location in the non-target interval. In addition, the duration of cue presentation was shortened to 47-ms, and the ISI was shortened to 47-ms (Fig. 2).
3.2. Results and discussion
As can be seen in Fig. 4, the results of this experiment replicated the central attentional impairment previously found for transient attention: A significant cueing X eccentricity interaction emerged: accuracy was higher in the cued than neutral trials when the target appeared at the far periphery, but was lower when the target appeared at central locations [F(4, 68) = 2.85, p < 0.05, ηp2 = 0.14]. These results differ from those in Experiment 1 in which attention improved performance at all eccentricities. We compared the neutral functions of Experiments 1 and 2 to ensure that the performance across eccentricities follows the same pattern, and found no significant difference [p > 0.1].
Fig. 4.
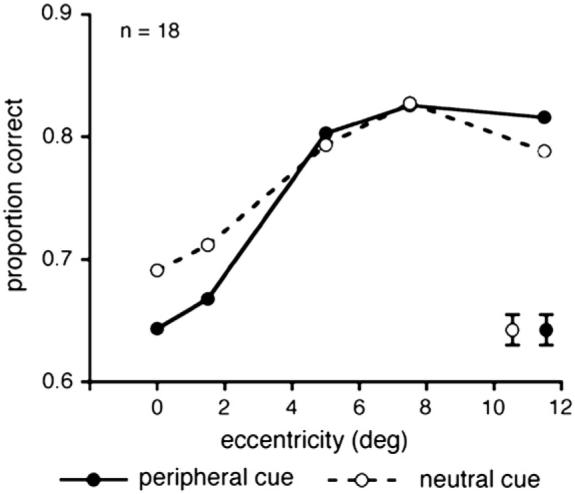
Average proportion of correct responses as a function of target eccentricity and cue type (peripheral cue vs. neutral cue) in Experiment 2. Precueing the target location with a peripheral cue (filled circles, continuous line) improved performance in the periphery, but impaired performance at more central locations. Error bars are indicated in the lower right of the graph and represent ± 1 SEM per cueing condition, averaged across eccentricities.
The finding of Experiment 1 that sustained attention improves texture segmentation at all eccentricities is not an artifact of the specific experimental conditions used in that experiment, because keeping these identical stimuli and task conditions with a manipulation of transient attention resulted in a central attentional impairment. In Experiment 3 we test whether the benefit of sustained attention across target eccentricity can be observed with a different texture and task.
4. Experiment 3
The central performance drop is commonly attributed to a mismatch between the size of spatial filters at the fovea and the scale of the texture. Specifically, it has been suggested that the size of these filters at the fovea may be too small for the scale of the texture, resulting in a central performance drop. At more peripheral regions, filters' average size increases gradually, and is presumably optimal around the peak of performance. At farther eccentricities, the filters are too big and consequently performance at the far periphery also drops (e.g., Gurnsey et al., 1996; Yeshurun & Carrasco, 1998). Many studies lend further support to the spatial explanation of the central performance drop. For instance, the finding that when this texture is presented along the vertical meridian performance peaks at farther eccentricities in the lower than in the upper visual vertical meridian, agrees with the view that resolution is higher in the lower region than the upper region of the vertical meridian (Talgar & Carrasco, 2002). Moreover, adapting to high-spatial frequencies, but not to low-spatial frequencies, prior to performing this texture segmentation task, eliminates the central performance drop (Carrasco et al., 2006).
An alternative interpretation ascribes the central performance drop to temporal factors. According to this view, because processing speed increases with distance from the fovea (Breitmeyer, 1984; Carrasco, McElree, Denisova, & Giordano, 2003), and often in those studies the texture displays have been followed by a mask (e.g., Gurnsey et al. 1996; Joffe & Scialfa 1995; Kehrer 1989; Kehrer 1997; Morikawa 2000; Talgar & Carrasco, 2002; Yeshurun & Carrasco, 1998), the drop in performance at central locations could be explained by differences in the speed of visual processing as a function of eccentricity. Specifically, at the fovea, the post mask might have interrupted the slower processing prior to completion, resulting in central performance drop, whereas at mid-peripheral locations the faster processing could be effectively completed prior to the presentation of the mask (Kehrer, 1989; Morikawa, 2000).
To guarantee that the only operant interpretation of the central performance drop is constrained by spatial factors, we did not employ a backward mask in this experiment. Instead, to prevent ceiling performance, we introduced variability in the orientation of the textures' line elements and increased the possible number of target locations. The resulting texture stimuli were very similar to the ones used by Potechin and Gurnsey (2003). Thus, whereas the average orientation of line elements in the texture display was ±45° from vertical, the actual orientation of each line element was chosen at random from a uniform distribution of orientations (see Fig. 5 for an example of the texture display). The range of sampled orientations around the mean orientation is referred to as “orientation bandwidth” (Potechin & Gurnsey, 2003). In general, as the orientation bandwidth increases the target patch becomes harder to detect. This and variants of this method of limiting performance have been used in several studies of texture perception (Kingdom, Keeble, & Moulden, 1995; Motoyoshi & Nishida, 2001; Potechin & Gurnsey, 2003; Wolfson & Landy, 1998). In this experiment, we manipulated sustained attention by precueing target location with a central cue, as in Experiment 1, in a Yes-No detection task.
Fig. 5.

Sample of the texture display used in Experiment 3. To limit performance, variability is introduced in the line elements' orientation (average orientation is ±45° from vertical). A 3 × 3 target patch is visible in the upper left quadrant of the display.
4.1. Methods
4.1.1. Observers
Eight students from NYU participated in Experiment 3. All observers had normal or corrected-to-normal vision.
4.1.2. Apparatus
The apparatus was similar to that used in Experiment 1 except for the following: The monitor resolution was set to 1600 × 1200 pixels at a frame rate of 75 Hz, and observers viewed the monitor from a distance of 100 cm. Stimuli were generated and presented using Matlab 6.0 and the Psychophysics Toolbox extensions (Brainard, 1997; Pelli, 1997).
4.1.3. Stimuli
The textures were composed of white line elements (each subtending 0.1 × 0.17°) presented on a black background. The luminance of the line elements was approximately 16 cd/m2 and the one of the background was about 0.003 cd/m2. The luminance of the texture display was about 0.45 cd/m2. The lines' average orientation was ±45° from vertical but the actual orientation of each line element was chosen at random (for each trial) from a uniform distribution of orientations. The range of sampled orientations—the orientation bandwidth—was adjusted for each observer during practice sessions to achieve an overall performance level of around 70% correct, the median was 100°, and it ranged from 55° to 110°. Each texture display consisted of 1813 line elements arranged on a virtual grid of 37 rows × 49 columns, subtending 16.9 × 22.3°. The line elements were jittered by 0.17°. In half of the trials the texture display contained a target patch comprising 3 × 3 line elements (subtending 1.37 × 1.37°), whose average orientation was orthogonal to the average orientation of the background texture (Fig. 5). The target could appear at one of 33 possible locations along the diagonal meridians, corresponding to 9 possible eccentricities, yet to allow the observers to easily associate each cue digit to a specific target location, only 17 possible locations were employed during each experimental session (4 locations on each of the 4 non-cardinal segments of the diagonal meridians—NE, NW, SE or SW—in addition to the central location). Thus, in half of the sessions the target center was placed at one of these eccentricities: 0°, 2.4°, 4.8°, 7.2°, 9.5°, and in the other half it was centered at 0°, 1.2°, 3.6°, 6°, 8.4°. We chose the diagonal meridians to prevent confounds due to asymmetries in performance along the vertical meridian in texture segregation (Graham, Rico, Offen, & Scott, 1999; Talgar & Carrasco, 2002), and along the vertical and horizontal meridian in other spatial resolution tasks (Carrasco et al., 2002). Thus, we could average performance at a given eccentricity across the four quadrants and increase statistical power.
On half of the trials, the cued trials, the cue was a central attentional cue indicating the location of the texture target. The central cue consisted of a small light gray tilted line (0.1 × 0.4°) and a digit appearing in the center of the display (within a region of 0.8 × 0.5 ). The small line indicated the quadrant (NE, NW, SE or SW) where the target, if present, would appear. The digit indicated the distance along the non-cardinal meridian in which the target would appear; the digit “0” indicated the central location, and the digits “1–4” indicated progressively more eccentric locations. Thus, for trials in which a target was present, the attentional cue indicated the exact location of the target with 100% validity. On the rest of the trials, the neutral trials, a large light gray square framing the entire texture display served as the neutral cue, indicating that the target, if present, was equally likely to appear at any of the possible locations. Both attentional and neutral cues conveyed information regarding the display onset.
4.1.4. Procedure
Each trial began with a fixation cross at the center of the screen (Fig. 6). 500-ms after the onset of the fixation cross the cue was presented for 100-ms and was followed by an ISI of 200-ms. Half of the trials were cued trials and half were neutral trials. After the ISI, the texture was displayed for 30-ms. For each cue condition, the texture display contained the target patch on 50% of the trials. The observers were instructed to keep their eyes fixated at the center (eye positions were monitored), and were asked to indicate whether the texture display contained a target. A yes-no detection task has been employed previously in studies reporting the central performance drop (Gurnsey et al., 1996). Given that in such a task only one location is pre-cued per trial, it may be less taxing for observers to allocate attention.
Fig. 6.
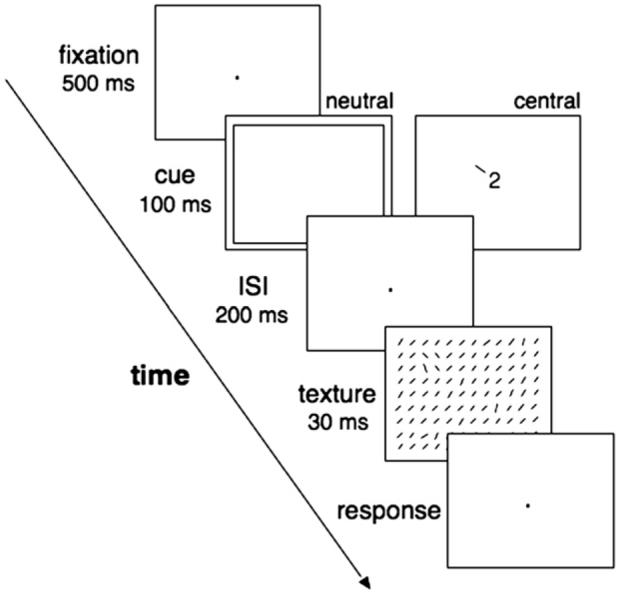
Schema of the frame sequence in a typical trial of Experiments 3, 4 and pre-cue condition of Experiment 5 (yes-no detection task). In this example, the target is present. For clarity, the stimuli are represented in black on a white background, in the actual experiment stimuli where white (the texture line elements) or light gray (the cues, the fixation cross) on a black background.
As in Experiment 1, to enable observers to associate the symbolic cues to a given location on the screen, at the beginning of each block of trials, a display was shown comprising the digits, later used as the cues, placed at their corresponding location. All observers had at least 1 h of practice (288 trials) to familiarize them with the task and to establish the associative connection between the symbolic cue and the designated location. Moreover, every experimental session (288 trials) began with 72 practice trials. Each observer participated in 4 sessions, for a total of 1152 experimental trials. Extensive practice was provided so that observers would learn the association between the central cue and the location it indicated, thus ensuring that performance was stable before data collection.
4.2. Results and discussion
We conducted a two-way (cue type X target eccentricity) repeated-measures ANOVA on the d′ values (Macmillan & Creelman, 1991). This analysis revealed a significant main effect of cue type: Performance was significantly better in the cued than the neutral trials [F(1, 7) = 47.851, p < 0.001, ηp2 = 0.872; Fig. 7]. The analysis also revealed a significant main effect of target eccentricity [F(8, 56) = 35.964, p < 0.001, ηp2 = 0.837]: As in previous studies (e.g., Gurnsey et al., 1996; Joffe & Scialfa, 1995; Kehrer, 1989; Morikawa 2000; Talgar & Carrasco, 2002; Yeshurun & Carrasco, 1998, 2008), performance peaked at the mid periphery and deteriorated at more central or farther peripheral eccentricities. The finding of a central performance drop even without the employment of a post-mask is consistent with demonstrations that a backward mask is not necessary to elicit a central performance drop (Morikawa, 2000; Potechin & Gurnsey, 2003), and it lends further support to the spatial interpretation of the central performance drop. Moreover, as in Experiment 1, there was no significant cueing X eccentricity interaction (p > 0.10). Directing sustained attention to the target location improved performance at all target eccentricities except at the farthest periphery, where performance was at chance.
Fig. 7.
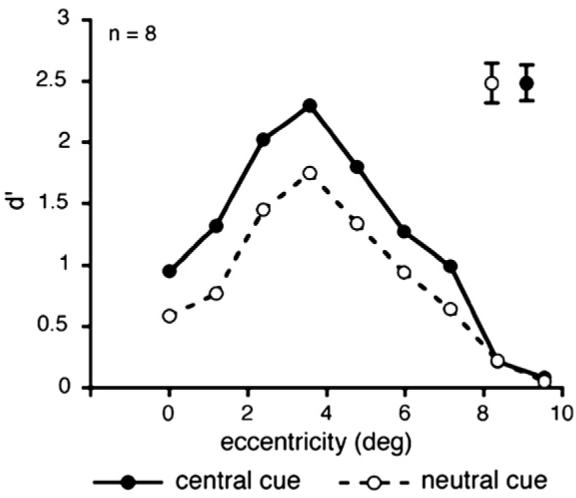
Average d′ across observers as a function of target eccentricity and cue type (central cue vs. neutral cue) in Experiment 3. Smaller texture scale (100 cm viewing distance). In the neutral condition (empty circles, dashed line), performance peaks at mid-periphery (3.6° of eccentricity) and drops towards more central and more peripheral locations, replicating the central performance drop. Precueing the target location with a central cue (filled circles, continuous line) improved performance across eccentricity. Error bars are indicated in the upper right of the graph and represent ± 1 SEM per cueing condition, averaged across eccentricities.
Analysis of RT on target present trials indicated that there were no speed–accuracy tradeoffs. There was a significant main effect of eccentricity [F(8, 56) = 3.505, p < 0.005, ηp2 = 0.334]: RTs were faster at mid-peripheral locations than at the more central or more peripheral locations; a significant main effect of cueing condition [F(1, 7) = 16.372, p = 0.005, ηp2 = 0.7]: RTs were faster in the cued than the neutral trials; as well as a significant interaction [F(8, 56) = 4.767, p < 0.001, ηp2 = 0.405]. This interaction emerged because RTs in the cued condition were faster than RTs in the neutral condition at mid-peripheral locations but not at the more central or more peripheral locations.
In sum, in this experiment we tested the generalizability of the finding that sustained attention improves performance across target eccentricity for this texture segmentation task. We introduced variability in the orientation of the textures' line elements, to avoid the employment of a post mask, and increased the number of possible target locations. Thus, the spatial uncertainty was larger than in Experiment 1. In addition, we used a yes-no rather than a 2IFC task. Regardless of these differences, we replicate the findings of Experiment 1: sustained attention improved texture segmentation across eccentricities, and there was no attentional impairment at central locations. These findings indicate that the attentional benefit that emerged in both experiments is robust and can be generalized to different textures and tasks. In Experiment 4 we explore whether the scale of the texture affects this benefit and the flexibility of sustained attention with respect to differences in task demands (in terms of optimal spatial resolution) introduced by different texture scales.
5. Experiment 4
The goal of this experiment was twofold: First, to further characterize how performance with the texture introduced by Potechin and Gurnsey (2003), which is limited by spatial constraints, varies with variation in the scale of the texture. Second, to investigate whether the effects of sustained attention on performance with this texture also vary as a function of the texture scale.
Previous studies that investigated the central performance drop with texture displays that do not include orientation variability found that performance peaks at different eccentricities when the scale of the texture is manipulated by changing the size of the textural elements, the space between them, or the viewing distance (e.g., Gurnsey et al., 1996; Joffe & Scialfa, 1995; Kehrer, 1989). Enlarging the scale of the texture shifts the peak of performance to farther eccentricities, supporting the idea that the segmentation of larger textures requires larger filters, which are more abundant at farther eccentricities. In view of that, when we manipulated transient attention we hypothesized that if the increase in texture scale extends the mismatch between the texture's scale and filters' size farther towards the periphery, and if attending to a location is similar to reducing the size of spatial filters, then directing transient attention to the target location of an enlarged texture should impair performance for a wider range of eccentricities. Indeed when we enlarged the scale of the texture, by moving the observers closer to the screen, the peak of performance shifted to farther eccentricities, and transient attention impaired performance over a larger range of central eccentricities (Yeshurun & Carrasco, 1998).
To better understand the factors that affect performance with the texture used in Experiment 3, in which variability in the orientation of the textures' elements was introduced, in this experiment we doubled the texture scale (by halving the viewing distance). Given that the central performance drop in this task has been interpreted in terms of spatial resolution, we hypothesized that the peak of performance with the enlarged texture would be displaced towards a more peripheral location. Moreover, enlarging the texture scale allows us to pose an interesting question regarding the flexibility of sustained attention. Given that at a certain eccentricity spatial resolution may be too low for the smaller texture (i.e., Experiment 3), yet too high for the enlarged texture employed in this experiment, we asked: Can sustained attention enhance performance at such eccentricity for both texture scales?
5.1. Methods
5.1.1. Observers
Four observers from NYU, who had participated in Experiment 3, also participated in Experiment 4.
5.1.2. Apparatus, stimuli and procedure
The apparatus, stimuli and procedure were identical to Experiment 3 except for the fact that the viewing distance was 50 cm, hence, doubling the degrees of visual angle subtended by the texture display.
5.2. Results and discussion
First, similar to Experiment 3, we conducted a two-way (cue type X target eccentricity) repeated-measures ANOVA on the d′ values. Like Experiments 1 and 3 we found a significant main effect of cue type [F(1, 3) = 37.442, p < 0.001, ηp2 = 0.926] and of eccentricity [F(8, 24) = 39.512, p < 0.001, ηp2 = 0.929], but here we found a significant interaction between these two factors [F(8, 24) = 6.705, p < 0.001, ηp2 = 0.691]. The significant interaction found here is due to the fact that there is a large effect of cueing at the central and mid-peripheral locations, but no difference at the three most eccentric locations (Fig. 8, note that the abscissa scale is twice that of Fig. 7), which may be due to a difficulty in precisely allocating sustained attention in the far periphery. This pattern of results is the same as the one found in Experiment 3 (see Fig. 7).
Fig. 8.
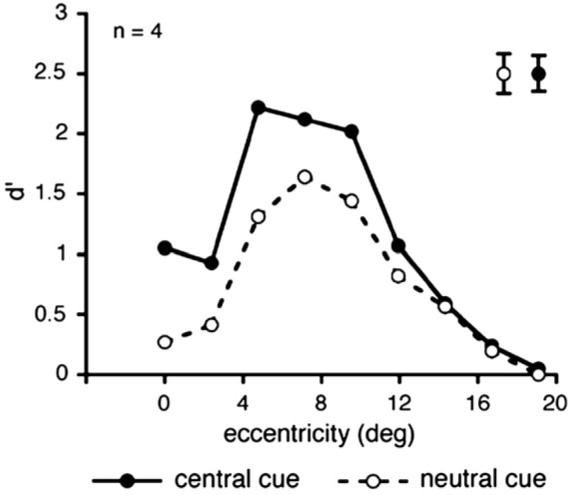
Average d′ across observers as a function of target eccentricity and cue type (central cue vs. neutral cue) in Experiment 4. Larger texture scale (50 cm viewing distance). Notice how the scale on the abscissa is doubled compared to Fig. 7. In the neutral condition (open circles, dashed line), performance peaks at 7.2° of eccentricity and drops towards more central and more peripheral locations, replicating the central performance drop. Doubling the texture scale shifted the peak of performance to more peripheral locations. Precueing the target location with a central cue (filled circles, continuous line) improved performance across eccentricity. Error bars are indicated in the upper right of the graph and represent ± 1 SEM per cueing condition, averaged across eccentricities.
As in Experiment 3, RT analysis on target present trials revealed no speed-accuracy trade-off. There was a significant main effect of eccentricity [F(8, 24) = 4.120, p < .0.005, ηp2 = 0.579]: RTs were faster at mid-peripheral locations than at the more central or more peripheral locations; a significant main effect of cueing condition [F(1, 3) = 28.772, p < 0.05, ηp2 = 0.906]: RTs were faster in the cued than the neutral trials; as well as a significant interaction between eccentricity and cueing [F(8, 24) = 4.245, p < 0.005, ηp2 = 0.586]. This interaction emerged because whereas observers were faster in the cued than neutral trials at mid-peripheral locations, this effect was not present at the most central or most peripheral locations. Thus, the general pattern of results found here with the larger texture scale is very similar to that found with the smaller scale of Experiment 3.
Second, we compared the data of Experiments 3 and 4. In accordance with previous studies that manipulated the scale of the texture (e.g., Gurnsey et al., 1996; Yeshurun & Carrasco, 1998), the peak of performance in neutral trials was shifted from 3.6° to 7.2° when the scale of the texture was doubled (Experiment 3 vs. Experiment 4, respectively). This shift lends further support to the spatial interpretation of the central performance drop with this texture. However, in contrast to our findings that the effects of transient attention on texture segmentation depend on both the target eccentricity and the texture scale (Yeshurun & Carrasco, 1998), here we found that the effects of sustained attention are independent of either factor. That is, sustained attention improved texture segmentation at all eccentricities (unless performance is at chance level) and with both texture scales. Specifically, in a given small range of eccentricities (4.8–7.2°, highlighted in blue in Fig. 9) where performance is not optimal, presumably because at that eccentricity spatial resolution is too low for the smaller texture scale yet too high for the enlarged texture, sustained attention enhanced performance for both texture scales. This finding portrays sustained attention as a more flexible mechanism that is able to aid performance regardless of the spatial resolution at a given location, and it is consistent with the hypothesis that to optimize performance sustained attention can either increase spatial resolution when it is too low for the task (small texture) or decrease spatial resolution when it is too high (enlarged texture) for the task.
Fig. 9.
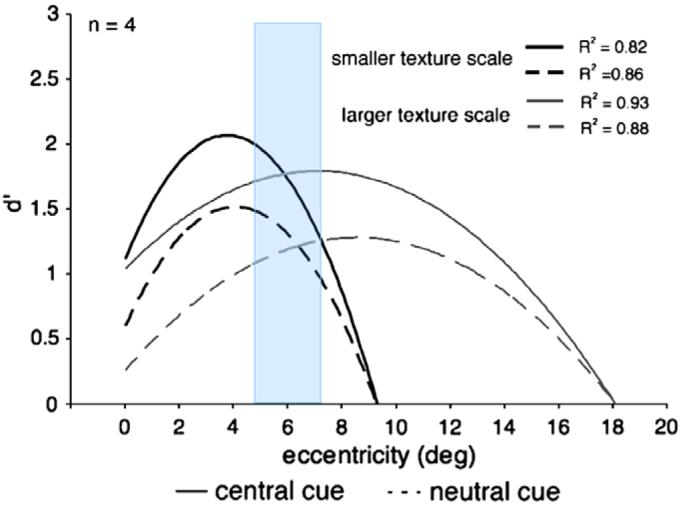
Second-order polynomials were fitted to average d′ data from 4 observers who participated both in Experiments 3 and 4. Fits for the larger texture scale are represented in gray, fits for the smaller texture in black. Fits for the neutral cue condition are represented as dashed lines, fits for the central cue condition are represented as continuous lines. Corresponding R2 values are indicated on the graph. In the neutral condition the peak of performance shifts to farther eccentricities when the scale of the texture is enlarged. In the range of eccentricities from 4.8°–7.2° (highlighted in blue), where performance is not optimal, sustained attention enhanced performance for both texture scales (see text). (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this paper.)
An alternative account of the eccentricity-independent effect of the sustained cue would attribute it to a mechanism that affects texture segmentation by improving the signal to noise ratio at all eccentricities by reducing location uncertainty. We evaluate the contribution of this mechanism in the next experiment.
6. Experiment 5
Some authors have explained precueing effects as the result of a reduction of location uncertainty. That is, given that the cue allows observers to prioritize information gathered at the relevant location, it reduces the statistical noise introduced at the irrelevant locations. According to this view, the benefit in performance found at cued locations can be ascribed to a reduction of the statistical uncertainty in the decision process rather than to changes in the quality of the representation of the texture (e.g., Eckstein, Shimozaki, & Abbey, 2002; Palmer, 1994; Palmer, Ames, & Lindsey, 1993; Shaw, 1984; Sperling & Dosher, 1986). This experiment was conducted to directly evaluate the contribution of location uncertainty reduction at the decisional level to the performance benefit with sustained attention (Experiments 1, 3 and 4). We compared the effect of the sustained pre-cues to the effect of post-cues, which indicate the target location after the offset of the texture display. Spatial post-cues, like post-masks, are considered to effectively reduce location uncertainty (e.g., Carrasco, Penpeci-Talgar, & Eckstein, 2000; Carrasco & Yeshurun, 1998; Kinchla et al. 1995; Lu & Dosher, 2004; Luck, Hillyard, Mouloua, & Hawkins, 1996; Luck et al., 1994; Smith 2000). Both pre- and post-cues reduce location uncertainty, as both allow the observer to assign lower weights to information extracted from the non-cued locations; however, only the pre-cues allow for a change in the quality of the texture representation due to the advanced allocation of attention to the location of the upcoming target. Thus, any additional benefit yielded by pre-cues compared to post-cues could be ascribed to an attentional modulation of the quality of the texture representation rather than to the mere reduction of location uncertainty at the decisional stage.
6.1. Methods
6.1.1. Observers
Six observers participated in Experiment 5; one of them had participated in Experiments 3 and 4. All observers had normal or corrected-to-normal vision.
6.1.2. Apparatus, stimuli and procedure
The apparatus, stimuli and procedure were identical to Experiment 3 except for the fact that there were 5 instead of 9 target eccentricities (range 0°–8.4° instead of 0°–9.5°), and 4 (instead of 2) experimental conditions: neutral pre-cue, central (attention) pre-cue, neutral post-cue and central post-cue. The timing of the pre-cues was identical to that of Experiments 3 and 4. In the post-cue conditions, the order of the texture display and cue was inverted, and the duration and ISI were the same as in the pre-cue conditions. Observers were informed that both the central pre-cues and the central post-cues were 100% valid, indicating where, if present, the target was going to appear or had just appeared, respectively.
6.2. Results and discussion
As in Experiments 3 and 4, we conducted a two-way (cue type X target eccentricity) repeated-measures ANOVA on the d' values. We found a significant main effect of cue type [F(3, 15) = 18.05, p < 0.0001, ηp2 = 0.61] and of eccentricity [F(4, 20) = 19.70, p < 0.0001, ηp2 = 0.9] but no significant interaction [F < 1; Fig. 10]. Performance for the central (attention) pre-cue was higher than for the neutral pre-cue [F(1, 5) = 62.93, p < 0.0005, ηp2 = 0.75] whereas there was only a marginal difference between performance for the neutral post-cue and the central post-cue [F(1, 5) = 5.83, p < 0.1, ηp2 = 0.4]. Furthermore, performance for the central pre-cue was significantly higher than for the central post-cue [F(1, 5) = 11.34, p = 0.01, ηp2 = 0.52] whereas performance for the neutral pre-cue and neutral post-cue conditions did not differ [F(1, 5) = 1].
Fig. 10.
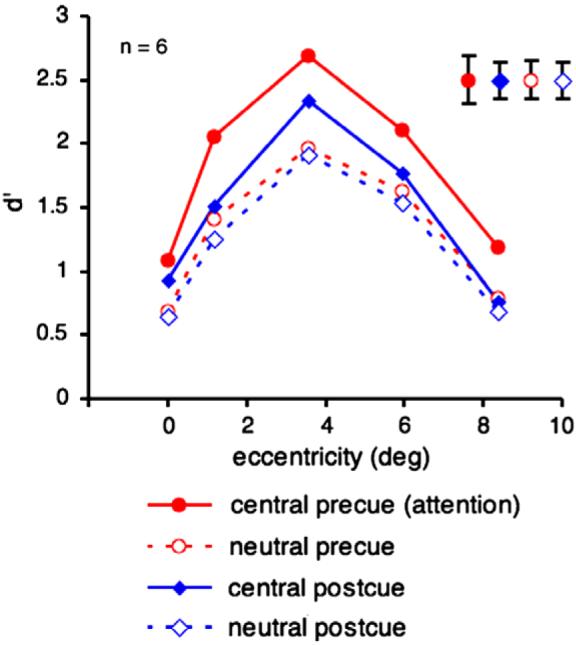
Average d' across observers as a function of target eccentricity and cue type (central pre-cue, central post-cue, neutral pre-cue, neutral post-cue) in Experiment 5. Precueing the target location with a central precue improved performance across all eccentricities in comparison to the other three conditions. Error bars are indicated in the upper right of the graph and represent ± 1 SEM per cueing condition, averaged across eccentricities.
Analysis of RT on target present trials indicated that there were no speed-accuracy tradeoffs for either the precue or the post-cue conditions. There was a significant main effect of eccentricity for both pre-cue and post-cue conditions (p < 0.005): RTs were faster at the central location and increased towards more foveal and more peripheral locations. For the pre-cue condition, RT differed more at the foveal and peripheral locations than at the central location; for the post-cue condition, RT were faster for the central-cue than the neutral-cue at all locations.
The results show that performance with the central precue, which triggers sustained attention, was significantly higher than performance with its neutral condition, whereas performance for the central post-cue was only marginally higher than its neutral condition. Moreover, the central pre-cue elicited a significantly better performance than the central post-cue. These results indicate that the benefit of the central pre-cue went well beyond the mere effect of location uncertainty at the decisional stage–it improved the quality of the texture representation.
7. General discussion
The goal of this study was to test whether sustained attention can affect performance in a basic texture segmentation task, and if so whether this effect will depend on the eccentricity of the texture-target, as does the effect of transient attention (i.e., enhancement at peripheral target locations and decrement at central target locations; Experiment 2). In four experiments (Experiments 1, 3, 4 and 5) we compared performance on trials in which central, symbolic cues indicated the location of the target prior to its presentation, allowing observers to direct sustained attention in advance to that location, with performance on trials in which a neutral cue did not convey information regarding the target location. The pattern of results was very similar in all four experiments: sustained attention, like transient attention, can affect performance in texture segmentation. Thus, even though texture segmentation is believed to take place at relatively early stages of visual processing (e.g., Bergen & Landy, 1991; Braun & Sagi, 1990; Chubb, Olzak, & Derrington, 2001; Dakin, Williams, & Hess, 1999; Julesz, 1981; Lamme, 1995; Lamme et al., 1993; Purpura et al., 1994), and although sustained attention seems to operate at a later level of processing than transient attention (e.g., Corbetta & Shulman, 2002; Kastner & Ungerleider, 2000; Liu et al., 2007; Nakayama & Mackeben, 1989), it can nevertheless affect performance in basic texture segmentation tasks as the ones employed here.
This finding is consistent with psychophysical studies showing that sustained attention increases other basic visual dimensions, such as contrast sensitivity (Dosher & Lu, 2000; Ling & Carrasco, 2006), as well as with neurophysiological studies demonstrating, by means of either single-cell recordings (e.g., Luck et al., 1997; Maunsell & Cook, 2003; McAdams & Reid, 2005; Motter, 1994; Reynolds & Desimone, 1999; Womelsdorf et al., 2006) or fMRI studies (e.g., Brefczynski & DeYoe, 1999; Gandhi, Heeger, & Boynton, 1999; Liu, Pestilli, & Carrasco, 2005; Martinez et al., 1999; Somers, Dale, Seiffert, & Tootell, 1999), attentional modulation of activity in early visual areas, such as V1 and V2.
However, in contrast to transient attention, the effects of sustained attention did not vary as a function of eccentricity. Directing sustained attention to the target location improved performance at all eccentricities (unless performance was at chance level). This eccentricity-independent attentional benefit was found even when: (a) the experimental conditions were identical to those used with transient attention (Experiment 1 vs. Experiment 2); (b) performance was not limited by a post mask, spatial uncertainty was doubled, and the task was a yes-no task (Experiments 3 and 5); and (c) the scale of the texture was doubled by halving the viewing distance (Experiment 4).
Experiment 5 evaluated the contribution of location uncertainty to the performance benefit found with the central, attentional cues in Experiments 1, 3 and 4. To control for location uncertainty at the decisional stage, we employed both pre-cues, identical to the ones used in Experiments 1, 3 and 4, and post-cues that appear after the presentation of the target display. Such post-cues can effectively reduce location uncertainty by indicating the exact target location, but they do not allow for any changes in the quality of the texture representation because they are presented after target offset (e.g., Carrasco & Yeshurun, 1998; Kinchla et al. 1995; Lu & Dosher, 2004; Luck et al., 1994). The fact that performance with the central, attentional pre-cue of Experiment 5 was significantly better than performance with the central post-cue indicates that the effect of sustained attention on texture segmentation goes well beyond reduction of location uncertainty at the decisional stage, and most likely reflect changes in the quality of the texture representation.
It is important to note that the effect of transient attention on texture segmentation, namely the attentional impairment of performance at central locations, could not be accounted for by uncertainty reduction because location uncertainty reduction would predict a benefit on performance throughout all eccentricities. A biologically plausible mechanism that can explain the central attentional impairment found with transient attention is an increase in spatial resolution (Carrasco et al. 2006; Talgar & Carrasco, 2002; Yeshurun & Carrasco, 1998, 2000, 2008).
In contrast, the finding that directing sustained attention to the target location via central pre-cues (Experiments 1, 3, 4 & 5) improved texture segmentation both at central and peripheral locations could be accounted for by an attentional mechanism that is capable of either enhancement or decrement of spatial resolution to optimize performance. According to this view, sustained attention optimized performance at all eccentricities via resolution enhancement at the periphery where performance is limited by a resolution that is too low, and via resolution decrement at central locations where performance is limited by a resolution that is too high. This view of sustained attention portrays a highly adaptive mechanism that can adjust its operation on a trial-by-trial basis. Indeed, the fact that it requires a longer cue-lead time than transient attention may allow it to adjust its operation.
Note, however, that the eccentricity-independent effects of sustained attention could also be attributed to an attentional mechanism that affects texture segmentation by improving the signal to noise ratio at all eccentricities through other means than resolution modification, like reduction of external noise at early levels of processing (e.g., Dosher & Lu, 2000), possibly via distracter suppression (e.g., Shiu & Pashler, 1994). The findings of this study cannot rule out such accounts, and a more decisive ruling regarding the nature of the mechanism underlying these effects of sustained attention awaits future research.
The finding that sustained attention affect texture segmentation in a different manner than transient attention is consistent with studies demonstrating differential effects for sustained and transient attention. For instance, Briand and Klein (1987; Briand, 1998) examined the effects of peripheral and central cues on performance in visual search. They found that with peripheral cues the effects of attention was larger for a conjunction search than for a feature search. However, with central cues, the magnitude of the cueing effect did not differ for feature and conjunction search. Another differential effect was demonstrated in a study in which the central cue and target appeared on opposite sides of the vertical meridian, both saccadic and manual responses were associated with greater costs than when they appeared on the same side (i.e., the meridian effect). In contrast, with peripheral cues, the vertical meridian had no effect on costs for either response system (Reuter-Lorenz & Fendrich, 1992). In addition, when the effects of sustained and transient attention were tested under low-noise vs. high-noise conditions, it was reported that sustained attention could affect performance only under high-noise conditions, but not under low-noise conditions (e.g., Dosher & Lu, 2000). Transient attention, however, could operate under both low-noise and high-noise conditions (Lu & Dosher, 1998; Lu & Dosher, 2000). A more recent study has shown that both sustained and transient attention increase contrast sensitivity, even in low-noise conditions, but whereas the former is mediated by a contrast gain mechanism, the latter seems to be mediated by both a contrast-gain and response-gain mechanisms (Ling & Carrasco, 2006). Finally, a recent study showed that an involuntary allocation of attention via peripheral non-informative cues impaired temporal order judgment, whereas a voluntary allocation of attention via central informative cues improved it (Hein, Rolke, & Ulrich, 2006).
In this study we found yet another differential effect of sustained and transient attentional mechanisms. Whereas the effects of transient attention on texture segmentation depended on the eccentricity of the texture target—improving performance at the periphery but impairing performance at central locations, the effect of sustained attention on texture segmentation was not modulated by target eccentricity—performance was improved at all eccentricities. These results suggest that whereas transient attention increases spatial resolution even when it is detrimental to performance, sustained attention can adjust its operation to optimize performance across target eccentricity. These different results might be due to the more adaptable nature of sustained attention in comparison to transient attention. A growing body of evidence suggests that sustained attention is a more flexible mechanism than transient attention. For example, whereas sustained attention allows selection of locations encoded in both egocentric coordinates (i.e., coding locations relative to the observer) and allocentric coordinates (i.e., coding locations relative to other items in the display), transient attention seems to operate exclusively in egocentric coordinates (Barrett, Bradshaw, & Rose, 2003; Barrett, Bradshaw, Rose, Everatt, & Simpson, 2001; Tipper, Jordan, & Weaver, 1999). Moreover, a study that employed the response-signal speed-accuracy trade-off (SAT) procedure, which enables conjoint measures of discriminability and temporal dynamics, has shown that with central cues, the observed benefits and costs increased with cue validity. However, with peripheral cues, the valid-cue benefits and the invalid-cue costs in both discriminability and processing speed were comparable across the range of cue validities. These results provide compelling time-course evidence that transient attention is automatic, but sustained attention can be flexibly allocated in a manner that increases the benefit of the valid cue and decreases the cost of the invalid cue (Giordano et al., 2004).
In conclusion, this study demonstrates that sustained attention, like transient attention, can affect texture segmentation. However, in contrast to transient attention, the effect of sustained attention on texture segmentation does not vary as a function of target eccentricity: directing sustained attention to the target location improved performance at both central and peripheral locations. This pattern of results indicates that the mechanism underlying sustained attention is different than the one underlying transient attention, with the former being more flexible, capable of optimizing performance across the visual field.
Acknowledgments
We thank Jennifer Corbett, Stuart Fuller, Anna Marie Giordano, Samuel Ling, Taosheng Liu, Franco Pestilli, and Dennis Santella for useful discussions and comments on previous drafts. We also thank Dennis Santella for assistance with data collection for Experiments 3 and 4. This research was supported by Grant No. 2001169 from the United States-Israel Binational Science Foundation (BSF), Jerusalem, Israel to Y.Y. and M.C., and by NIH to M.C. (R01 EY016200-01A2).
Footnotes
Given that fixation was broken in very few trials (∼3%) in all experiments, we analyzed all the data in all the experiments.
For all experiments described in this paper, when appropriate, significance levels were corrected for violations of the sphericity assumption using the Greenhouse–Geisser method; however, original degrees of freedom are reported.
References
- Barrett DJ, Bradshaw MF, Rose D. Endogenous shifts of covert attention operate within multiple coordinate frames: Evidence from a feature-priming task. Perception. 2003;32:41–52. doi: 10.1068/p3298. [DOI] [PubMed] [Google Scholar]
- Barrett DJ, Bradshaw MF, Rose D, Everatt J, Simpson PJ. Reflexive shifts of covert attention operate in an egocentric coordinate frame. Perception. 2001;30:1083–1091. doi: 10.1068/p3165. [DOI] [PubMed] [Google Scholar]
- Bergen JR, Landy MS. Computational modeling of visual texture segregation. In: Landy MS, Movshon JA, editors. Computational models of visual processing. MIT Press; Cambridge, MA: 1991. pp. 253–271. [Google Scholar]
- Brainard DH. The psychophysics toolbox. Spatial Vision. 1997;10:433–436. [PubMed] [Google Scholar]
- Braun J, Sagi D. Vision outside of the focus of attention. Perception & Psychophysics. 1990;48:45–58. doi: 10.3758/bf03205010. [DOI] [PubMed] [Google Scholar]
- Brefczynski JA, DeYoe EA. A physiological correlate of the ‘spotlight’ of visual attention. Nature Neuroscience. 1999;2:370–374. doi: 10.1038/7280. [DOI] [PubMed] [Google Scholar]
- Breitmeyer BG. Visual masking: An integrative approach. Oxford University Press; New York: 1984. [Google Scholar]
- Briand KA. Feature integration and spatial attention: More evidence of a dissociation between endogenous and exogenous orienting. Journal of Experimental Psychology: Human Perception and Performance. 1998;24:1243–1256. [Google Scholar]
- Briand KA, Klein RM. Is Posner's “beam” the same as Treisman's “glue”? On the relation between visual orienting and feature integration theory. Journal of Experimental Psychology: Human Perception and Performance. 1987;13:228–241. doi: 10.1037//0096-1523.13.2.228. [DOI] [PubMed] [Google Scholar]
- Caelli T. Three characteristics of visual texture segmentation. Spatial Vision. 1985;1:19–30. doi: 10.1163/156856885x00044. [DOI] [PubMed] [Google Scholar]
- Carrasco M, Frieder KS. Cortical magnification neutralizes the eccentricity effect in visual search. Vision Research. 1997;37(1):63–82. doi: 10.1016/s0042-6989(96)00102-2. [DOI] [PubMed] [Google Scholar]
- Carrasco M, Ling S, Read S. Attention alters appearance. Nature Neuroscience. 2004;7:308–313. doi: 10.1038/nn1194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carrasco M, Loula F, Ho Y-X. How attention enhances spatial resolution: Evidence from selective adaptation to spatial frequency. Perception & Psychophysics. 2006;68:1004–1012. doi: 10.3758/bf03193361. [DOI] [PubMed] [Google Scholar]
- Carrasco M, McElree B, Denisova K, Giordano AM. Speed of visual processing increases with eccentricity. Nature Neuroscience. 2003;6:699–700. doi: 10.1038/nn1079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carrasco M, Penpeci-Talgar C, Eckstein M. Spatial attention increases contrast sensitivity across the CSF: Support for signal enhancement. Vision Research. 2000;40:1203–1215. doi: 10.1016/s0042-6989(00)00024-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carrasco M, Williams PE, Yeshurun Y. Covert attention increases spatial resolution with or without masks: Support for signal enhancement. Journal of Vision. 2002;2:467–479. doi: 10.1167/2.6.4. [DOI] [PubMed] [Google Scholar]
- Carrasco M, Yeshurun Y. The contribution of covert attention to the set-size and eccentricity effects in visual search. Journal of Experimental Psychology: Human Perception & Performance. 1998;24:673–692. doi: 10.1037//0096-1523.24.2.673. [DOI] [PubMed] [Google Scholar]
- Cheal M, Lyon D. Central and peripheral precuing of forced-choice discrimination. Quarterly Journal of Experimental Psychology: Human Experimental Psychology. 1991;43A:859–880. doi: 10.1080/14640749108400960. [DOI] [PubMed] [Google Scholar]
- Chubb C, Olzak L, Derrington A. Second-order processes in vision: Introduction. Journal of the Optical Society of America A. 2001;18:2175–2178. doi: 10.1364/josaa.18.002175. [DOI] [PubMed] [Google Scholar]
- Chubb C, Talevich J. Attentional control of texture orientation judgments. Vision Research. 2002;42:311–330. doi: 10.1016/s0042-6989(01)00282-6. [DOI] [PubMed] [Google Scholar]
- Cohen JD, MacWhinney B, Flatt M, Provost J. PsyScope: A new graphic interactive environment for designing psychology experiments. Behavioral Research Methods, Instruments, and Computers. 1993;25(2):257–271. [Google Scholar]
- Corbetta M, Shulman GL. Control of goal-directed and stimulus-driven attention in the brain. Nature Reviews Neuroscience. 2002;3:201–215. doi: 10.1038/nrn755. [DOI] [PubMed] [Google Scholar]
- Dakin SC, Williams CB, Hess RF. The interaction of first- and second-order cues to orientation. Vision Research. 1999;39(17):2867–2884. doi: 10.1016/s0042-6989(98)00307-1. [DOI] [PubMed] [Google Scholar]
- De Valois RL, De Valois KK. Spatial vision. Oxford University Press; New York: 1988. [Google Scholar]
- Desimone R, Duncan J. Neural mechanisms of selective visual attention. Annual Review of Neuroscience. 1995;18:193–222. doi: 10.1146/annurev.ne.18.030195.001205. [DOI] [PubMed] [Google Scholar]
- Dosher BA, Lu L. Mechanisms of perceptual attention in precuing of location. Vision Research. 2000;40(1012):1269–1292. doi: 10.1016/s0042-6989(00)00019-5. [DOI] [PubMed] [Google Scholar]
- Eckstein MP, Shimozaki SS, Abbey CK. The footprints of visual attention in the Posner cueing paradigm revealed by classification images. Journal of Vision. 2002;2:25–45. doi: 10.1167/2.1.3. [DOI] [PubMed] [Google Scholar]
- Eriksen CWW, Hoffman JE. Temporal and spatial characteristics of selective encoding from visual displays. Perception & Psychophysics. 1972;12:201–204. [Google Scholar]
- Gandhi SP, Heeger DJ, Boynton GM. Spatial attention affects brain activity in human primary visual cortex. Proceedings of the National Academy of Sciences. 1999;96:3314–3319. doi: 10.1073/pnas.96.6.3314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gibson BS, Bryant TA. Variation in cue duration reveals top-down modulation of involuntary orienting to uninformative symbolic cues. Perception & Psychophysics. 2005;67(5):749–758. doi: 10.3758/bf03193530. [DOI] [PubMed] [Google Scholar]
- Giordano AM, McElree B, Carrasco M. Vision Sciences Society. Sarasota FL: 2004. On the automaticity and flexibility of covert attention; p. F59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Golla H, Ignashchenkova A, Haarmeier T, Their P. Improvement of visual acuity by spatial cueing: A comparative study in human and non-human primates. Vision Research. 2004;44(13):1589–1600. doi: 10.1016/j.visres.2004.01.009. [DOI] [PubMed] [Google Scholar]
- Graham N. Visual pattern analyzers. Oxford University Press; New York: 1989. [Google Scholar]
- Graham N, Rico M, Offen S, Scott W. Texture segregation shows only a very small lower-hemifield advantage. Vision Research. 1999;39:1171–1177. doi: 10.1016/s0042-6989(98)00170-9. [DOI] [PubMed] [Google Scholar]
- Gurnsey R, Pearson P, Day D. Texture segmentation along the horizontal meridian: nonmonotonic changes in performance with eccentricity. Journal of Experimental Psychology: Human Perception and Performance. 1996;22:738–757. doi: 10.1037//0096-1523.22.3.738. [DOI] [PubMed] [Google Scholar]
- Hein E, Rolke B, Ulrich R. Visual attention and temporal discrimination: Differential effects of automatic and voluntary cueing. Visual Cognition. 2006;13(1):20–50. [Google Scholar]
- Joffe KM, Scialfa CT. Texture segmentation as a function of eccentricity, spatial frequency and target size. Spatial Vision. 1995;9:325–342. doi: 10.1163/156856895x00034. [DOI] [PubMed] [Google Scholar]
- Jonides J. Voluntary vs. automatic control over the mind's eye's movement. In: Long JB, Baddeley AD, editors. Attention and performance IX. Lawrence Erlbaum Associates; Hillsdale, NJ: 1981. pp. 187–204. [Google Scholar]
- Julesz B. Textons, the elements of texture perception, and their interactions. Nature. 1981;290:91–97. doi: 10.1038/290091a0. [DOI] [PubMed] [Google Scholar]
- Kastner S, Ungerleider LG. Mechanisms of visual attention in the human cortex. Annual Review of Neuroscience. 2000;23:315–341. doi: 10.1146/annurev.neuro.23.1.315. [DOI] [PubMed] [Google Scholar]
- Kehrer L. Central performance drop on perceptual segregation tasks. Spatial vision. 1989;4:45–62. doi: 10.1163/156856889x00040. [DOI] [PubMed] [Google Scholar]
- Kehrer L. The central performance drop in texture segmentation: A simulation based on a spatial filter model. Biological Cybernetics. 1997;77:297–305. [Google Scholar]
- Kinchla RA. Department of Psychology. McMaster Univ.; Hamilton, Ontario: 1969. An attention operating characteristic in vision. (Technical Report, No. 29). [Google Scholar]
- Kinchla RA. Attention and performance IIX. Lawrence Erlbaum Associates; Hillsdale, NJ: 1980. The measurement of attention; pp. 213–238. [Google Scholar]
- Kinchla RA, Chen Z, Evert DL. Pre-cue effects in visual search: Data or resource limited? Perception & Psychophysics. 1995;57(4):441–450. doi: 10.3758/bf03213070. [DOI] [PubMed] [Google Scholar]
- Kingdom FAA, Keeble DRT, Moulden B. Sensitivity to orientation modulation in micropattern-based textures. Vision Research. 1995;35:79–91. doi: 10.1016/0042-6989(94)e0079-z. [DOI] [PubMed] [Google Scholar]
- Lamme VA. The neurophysiology of figure-ground segregation in primary visual cortex. Journal of Neuroscience. 1995;15:1605–1615. doi: 10.1523/JNEUROSCI.15-02-01605.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lamme VAF, Van Dijk BW, Spekreijse H. Organization of texture segregation processing in primate visual cortex. Visual Neuroscience. 1993;10:781–790. doi: 10.1017/s0952523800006039. [DOI] [PubMed] [Google Scholar]
- Landy MS, Bergen JR. Texture segregation and orientation gradient. Vision Research. 1991;31:679–691. doi: 10.1016/0042-6989(91)90009-t. [DOI] [PubMed] [Google Scholar]
- Landy MS, Graham N. Visual Perception of Texture. In: Chalupa LM, Werner JS, editors. The visual neuroscience. MIT Press; Cambridge, MA: 2004. pp. 1106–1118. [Google Scholar]
- Ling S, Carrasco M. Sustained and transient covert attention enhance the signal via different contrast response functions. Vision Research. 2006;46:1210–1220. doi: 10.1016/j.visres.2005.05.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu T, Pestilli F, Carrasco M. Transient attention enhances perceptual performance and fMRI response in human visual cortex. Neuron. 2005;45:469–477. doi: 10.1016/j.neuron.2004.12.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu T, Stevens ST, Carrasco M. Comparing the time course and efficacy of spatial and feature-based attention. Vision Research. 2007;47(1):108–113. doi: 10.1016/j.visres.2006.09.017. [DOI] [PubMed] [Google Scholar]
- Lu Z-L, Dosher BA. External noise distinguishes attention mechanisms. Vision Research. 1998;38(9):1183–1198. doi: 10.1016/s0042-6989(97)00273-3. [DOI] [PubMed] [Google Scholar]
- Lu Z-L, Dosher BA. Spatial attention: Different mechanisms for central and peripheral temporal precues? Journal of Experimental Psychology: Human Perception and Performance. 2000;26:1534–1548. doi: 10.1037//0096-1523.26.5.1534. [DOI] [PubMed] [Google Scholar]
- Lu Z-L, Dosher BA. Spatial attention excludes external noise without changing the spatial frequency tuning of the perceptual template. Journal of Vision. 2004;4(10):955–966. doi: 10.1167/4.10.10. 10. http://journalofvision.org/4/10/10/, doi:10.1167/4.10.10. [DOI] [PubMed] [Google Scholar]
- Luck SJ, Chelazzi L, Hillyard SA, Desimone R. Neural mechanisms of spatial selective attention in areas V1, V2, and V4 of Macaque visual cortex. Journal of Neurophysiology. 1997;77:24–42. doi: 10.1152/jn.1997.77.1.24. [DOI] [PubMed] [Google Scholar]
- Luck SJ, Hillyard SA, Mouloua M, Hawkins HL. Mechanisms of visual-spatial attention: Resource allocation or uncertainty reduction? Journal of Experimental Psychology: Human Perception and Performance. 1996;22:725–737. doi: 10.1037//0096-1523.22.3.725. [DOI] [PubMed] [Google Scholar]
- Luck SJ, Hillyard SA, Mouloua M, Woldorff MG, Clark VP, Hawkins HL. Effects of spatial cuing on luminance detectability: Psychophysical and electrophysiological evidence for early selection. Journal of Experimental Psychology: Human Perception and Performance. 1994;20:887–904. doi: 10.1037//0096-1523.20.4.887. [DOI] [PubMed] [Google Scholar]
- Macmillan NA, Creelman CD. Detection theory: A user's guide. Cambridge University Press; Cambridge, England: 1991. [Google Scholar]
- Martinez A, Anllo-Vento L, Sereno MI, Frank LR, Buxton RB, Dubowitz DJ, Wong EC, Hinrichs H, Heinze HJ, Hillyard SA. Involvement of striate and extrastriate visual cortical areas in spatial attention. Nature Neuroscience. 1999;2(4):364–369. doi: 10.1038/7274. [DOI] [PubMed] [Google Scholar]
- Maunsell JHR, Cook EP. The role of attention in visual processing. In: Parker A, Derrington A, Blakemore C, editors. The physiology of cognitive processes. Oxford University Press; Oxford, UK: 2003. pp. 157–172. [Google Scholar]
- Mayfrank L, Kimmig H, Fischer B. In: Eye movements: From physiology to cognition. O'Regan JK, Levy-Schoen A, editors. North-Holland; New York: 1987. pp. 37–45. [Google Scholar]
- McAdams CJ, Reid RC. Attention modulates the responses of simple cells in monkey primary visual cortex. Journal of Neuroscience. 2005;25(47):11023–11033. doi: 10.1523/JNEUROSCI.2904-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meinecke C, Kehrer L. Peripheral and foveal segmentation of angle textures. Perception & Psychophysics. 1994;56:326–334. doi: 10.3758/bf03209766. [DOI] [PubMed] [Google Scholar]
- Moran J, Desimone R. Selective attention gates visual processing in the extrastriate cortex. Science. 1985;229:782–784. doi: 10.1126/science.4023713. [DOI] [PubMed] [Google Scholar]
- Morikawa K. Central performance drop in texture segmentation: the role of spatial and temporal factors. Vision Research. 2000;40:3517–3526. doi: 10.1016/s0042-6989(00)00170-x. [DOI] [PubMed] [Google Scholar]
- Motoyoshi I, Nishida S. Temporal resolution of orientation-based texture segregation. Vision Research. 2001;41:2089–2105. doi: 10.1016/s0042-6989(01)00096-7. [DOI] [PubMed] [Google Scholar]
- Motter BC. Neural correlates of attentive selection for color or luminance in extrastriate area V4. The Journal of Neuroscience. 1994;14:2178–2189. doi: 10.1523/JNEUROSCI.14-04-02178.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Müller HJ, Findlay JM. The effect of visual attention on peripheral discrimination thresholds in single and multiple element displays. Acta Psychologica. 1988;69:129–155. doi: 10.1016/0001-6918(88)90003-0. [DOI] [PubMed] [Google Scholar]
- Müller HJ, Rabbitt PM. Reflexive and voluntary orienting of visual attention: Time course of activation and resistance to interruption. Journal of Experimental Psychology: Human Perception and Performance. 1989;15:315–330. doi: 10.1037//0096-1523.15.2.315. [DOI] [PubMed] [Google Scholar]
- Nakayama K, Mackeben M. Sustained and transient components of focal visual attention. Vision Research. 1989;29:1631–1647. doi: 10.1016/0042-6989(89)90144-2. [DOI] [PubMed] [Google Scholar]
- Palmer J. Set-size effects in visual search: The effect of attention is independent of the stimulus for simple tasks. Vision Research. 1994;34(13):1703–1721. doi: 10.1016/0042-6989(94)90128-7. [DOI] [PubMed] [Google Scholar]
- Palmer J, Ames CT, Lindsey DT. Measuring the effect of attention on simple visual search. Journal of Experimental Psychology: Human Perception & Performance. 1993;19:108–130. doi: 10.1037//0096-1523.19.1.108. [DOI] [PubMed] [Google Scholar]
- Pelli DG. The VideoToolbox software for visual psychophysics: Transforming numbers into movies. Spatial vision. 1997;10:437–442. [PubMed] [Google Scholar]
- Pestilli F, Carrasco M. Attention enhances contrast sensitivity at cued and impairs it at uncued locations. Vision Research. 2005;45:1867–1875. doi: 10.1016/j.visres.2005.01.019. [DOI] [PubMed] [Google Scholar]
- Pestilli F, Viera G, Carrasco M. How do attention and adaptation affect contrast sensitivity? Journal of Vision. 2007;7(7):1–12. doi: 10.1167/7.7.9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Posner MI. Orienting of attention. Quarterly Journal of Experimental Psychology. 1980;32:3–25. doi: 10.1080/00335558008248231. [DOI] [PubMed] [Google Scholar]
- Potechin C, Gurnsey R. Backward masking is not required to elicit the central performance drop. Spatial vision. 2003;16:393–406. doi: 10.1163/156856803322552720. [DOI] [PubMed] [Google Scholar]
- Purpura KP, Victor JD, Katz E. Striate cortex extracts higher-order spatial correlations from visual textures. Proceedings of the National Academy of Sciences, USA. 1994;91:8482–8486. doi: 10.1073/pnas.91.18.8482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Remington R, Johnston JC, Yantis S. Attentional capture by abrupt onsets. Perception & Psychophysics. 1992;51:279–290. doi: 10.3758/bf03212254. [DOI] [PubMed] [Google Scholar]
- Reuter-Lorenz PA, Fendrich R. Oculomotor readiness and covert orienting: Differences between central and peripheral precues. Perception & Psychophysics. 1992;52:336–344. doi: 10.3758/bf03209150. [DOI] [PubMed] [Google Scholar]
- Reynolds JH, Chelazzi L. Attentional modulation of visual processing. Annual Review of Neuroscience. 2004;27:611–647. doi: 10.1146/annurev.neuro.26.041002.131039. [DOI] [PubMed] [Google Scholar]
- Reynolds JH, Desimone R. The role of neural mechanisms of attention in solving the binding problem. Neuron. 1999;24:19–29. doi: 10.1016/s0896-6273(00)80819-3. [DOI] [PubMed] [Google Scholar]
- Ristic J, Friesen CK, Kingstone A. Are eyes special? It depends on how you look at it. Psychonomic Bulletin & Review. 2002;9:507–513. doi: 10.3758/bf03196306. [DOI] [PubMed] [Google Scholar]
- Rovamo J, Virsu V. An estimation and application of the human cortical magnification factor. Experimental Brain Research. 1979;37:495–510. doi: 10.1007/BF00236819. [DOI] [PubMed] [Google Scholar]
- Shaw ML. Division of attention among spatial locations: A fundamental difference between detection of letters and detection of luminance. In: Bouma H, Bouwhuis DG, editors. Attention and performance X. Lawrence Erlbaum Associates; Hillsdale, NJ: 1984. pp. 109–120. [Google Scholar]
- Shiu L-P, Pashler H. Neglible effect of spatial precuing on identification of single digits. Journal of Experimental Psychology: Human Perception and Performance. 1994;20:1037–1054. [Google Scholar]
- Smith PL. Attention and luminance detection: Effects of cues, masks, and pedestals. Journal of Experimental Psychology: Human Perception and Performance. 2000;26:1401–1420. doi: 10.1037//0096-1523.26.4.1401. [DOI] [PubMed] [Google Scholar]
- Somers DC, Dale AM, Seiffert AE, Tootell RBH. Functional MRI reveals spatially specific attentional modulation in human primary visual cortex. Proceedings of the National Academy of Sciences. 1999;96:1663–1668. doi: 10.1073/pnas.96.4.1663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sperling G, Dosher BA. Strategy and optimization in human information processing. In: Boff K, Kaufman L, Thomas J, editors. Handbook of perception and performance. Vol. 1. Wiley; New York: 1986. pp. 21–265. [Google Scholar]
- Sperling G, Melchner MJ. The attention operating characteristic: Examples from visual search. Science. 1978;202:315–318. doi: 10.1126/science.694536. [DOI] [PubMed] [Google Scholar]
- Sutter A, Beck J, Graham N. Contrast and spatial variables in texture segregation: Testing a simple spatial-frequency channels model. Perception & Psychophysics. 1989;46:312–332. doi: 10.3758/bf03204985. [DOI] [PubMed] [Google Scholar]
- Talgar CP, Carrasco M. Vertical meridian asymmetry in spatial resolution: Visual and attentional factors. Psychonomic Bulletin & Review. 2002;9:714–722. doi: 10.3758/bf03196326. [DOI] [PubMed] [Google Scholar]
- Tipper SP, Jordan H, Weaver B. Scene-based and object-centered inhibition of return: Evidence for dual orienting mechanisms. Perception & Psychophysics. 1999;61:50–60. doi: 10.3758/bf03211948. [DOI] [PubMed] [Google Scholar]
- Tipples J. Eye gaze is not unique: Automatic orienting in response to noninformative arrows. Psychonomic Bulletin & Review. 2002;9:314–318. doi: 10.3758/bf03196287. [DOI] [PubMed] [Google Scholar]
- Wilson HR, Levi D, Maffei L, Rovamo J, De Valois R. The perception of form: retina to striate cortex. In: Spillman L, Werner JS, editors. Visual perception: The neurophysiological foundations. Academic Press; New York: 1990. pp. 231–272. [Google Scholar]
- Wolfson SS, Landy MS. Examining edge- and region-based texture analysis mechanisms. Vision Research. 1998;38:439–446. doi: 10.1016/s0042-6989(97)00153-3. [DOI] [PubMed] [Google Scholar]
- Womelsdorf T, Anton-Erxleben K, Pieper F, Treue S. Dynamic shifts of visual receptive fields in cortical area MT by spatial attention. Nature Neuroscience. 2006;9:1156–1160. doi: 10.1038/nn1748. [DOI] [PubMed] [Google Scholar]
- Yantis S. Attentional capture in vision. In: Kramer A, Coles M, Logan G, editors. Converging operations in the study of visual selective attention. American Psychological Association; Washington DC: 1996. pp. 45–76. [Google Scholar]
- Yeshurun Y, Carrasco M. Attention improves or impairs visual perception by enhancing spatial resolution. Nature. 1998;396:72–75. doi: 10.1038/23936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yeshurun Y, Carrasco M. Spatial attention improves performance in spatial resolution tasks. Vision Research. 1999;39:293–306. doi: 10.1016/s0042-6989(98)00114-x. [DOI] [PubMed] [Google Scholar]
- Yeshurun Y, Carrasco M. The locus of attentional effects in texture segmentation. Nature Neuroscience. 2000;3:622–627. doi: 10.1038/75804. [DOI] [PubMed] [Google Scholar]
- Yeshurun Y, Carrasco M. The effects of transient attention on spatial resolution and the size of the attentional cue. Perception and Psychophysics. 2008;70(1):104–113. doi: 10.3758/pp.70.1.104. [DOI] [PubMed] [Google Scholar]