

Abstract
To examine the role of microtubules in growth cone turning, we have compared the microtubule organization in growth cones advancing on uniform laminin substrates with their organization in growth cones turning at a laminin–tenascin border. The majority (82%) of growth cones on laminin had a symmetrical microtubule organization, in which the microtubules entering the growth cone splay out toward the periphery of the growth cone. Growth cones at tenascin borders had symmetrically arranged microtubules in only 34% of cases, whereas in the majority of cases the microtubules were displaced toward one-half of the growth cone, presumably stabilizing in the direction of the turn along the tenascin border. These results suggest that reorganization of microtubules could underlie growth cone turning. Further evidence for the involvement of microtubule rearrangement in growth cone turning was provided by experiments in which growth cones approached tenascin borders in the presence of nanomolar concentrations of the microtubule stabilizing compound, Taxol. Taxol altered the organization of microtubules in growth cones growing on laminin by restricting their distribution to the proximal regions of the growth cone and increasing their bundling. Taxol did not stop growth cone advance on laminin. When growing in the presence of Taxol, growth cones at tenascin borders were not able to turn and grow along the laminin–tenascin border, and consequently stopped at the border. Growth cones were arrested at borders for as long as Taxol was present (up to 6 h) without showing any signs of drug toxicity. These effects of Taxol were reversible. Together, these results suggest that microtubule reorganization in growth cones is a necessary event in growth cone turning.
Keywords: axonal pathfinding, Taxol, tenascin
Neural development is crucially dependent on the ability of growth cones to navigate precise routes through the embryo and to recognize appropriate target cells. Growth cones are sensitive to extrinsic guidance cues, such as chemotropic factors, cell adhesion glycoproteins, and extracellular matrix molecules, that influence the direction of axon growth (1–4). These cues interact with growth cone membrane receptors and lead, via intracellular signaling events, to changes in the growth cone cytoskeleton and hence in directional motility. Despite considerable effort to identify and characterize guidance molecules, we have only a rudimentary knowledge of how they regulate the growth cone cytoskeleton.
We have shown previously in an in vitro “choice assay” that chicken dorsal root ganglion (DRG) growth cones turn when confronting a sharp substrate border between a permissive (laminin) and a nonpermissive (tenascin) substrate (5, 6). At the border, growth cones make turns and grow along the laminin side. Tenascin (tenascin-C, see ref. 7) is a member of a family of extracellular matrix glycoproteins, which may function in the development of the nervous system (but see ref. 8). Particular attention has been paid to the possible role of tenascin in axon guidance due to its presence within tissue boundaries during development (9) and its up-regulation during the period of peripheral nerve regeneration following injury (10). Several in vitro studies have pointed to the ability of tenascin to promote neurite outgrowth when offered as a uniform substrate (11–15). We, and others, have shown that tenascin can act as a barrier to growth cones advancing on permissive substrates when it is offered as a substrate boundary (6, 16). In contrast to other molecules shown to be repulsive or inhibitory for advancing growth cones, this is not associated with growth cone collapse and neurite retraction. Instead, growth cones approaching tenascin borders change their direction of growth to avoid advancing into the tenascin-containing territory. These studies support the idea that tenascin might guide growth cones by repulsive or inhibitory mechanisms, as has been suggested for a variety of other molecules found within the developing and adult nervous system (17, 18). The response of the growth cone in turning at the border clearly involves intracellular signaling events that eventually lead to changes in the growth cone cytoskeleton that underlies the turning event. In the experiments reported here we have sought to understand what those changes might be.
The major components of the cytoskeleton of growth cones are microtubules and microfilaments (19–21). Microtubules are prominent in the central domain whereas microfilaments are concentrated in the peripheral, motile regions. In the axon, microtubules provide the substrate for fast axonal transport and are necessary for axon elongation. The function of microtubules in growth cones is less clear, whereas microfilaments are involved in growth cone motility (22). In the axon shaft, microtubules are bundled into fascicles but splay out on entering the proximal region of the growth cone (23–25). Occasionally, a few of these microtubules extend into the distal, motile domain of filopodia and lamellipodia, and may even enter filopodia (26, 27) where they lie alongside the filopodial actin filament bundle, suggesting a specific interaction (28).
Microtubules in these distal regions are extending and retracting, either by polymer sliding or by polymerization and depolymerization (29, 30). We have suggested that actin filament bundles in those filopodia that have interacted with extrinsic guidance cues may capture extending microtubules and stabilize them (26, 30). In our model, the activation of filopodial membrane receptors by extrinsic guidance cues may influence actin filament dynamics, either directly or through intermediate proteins. This may change actin filament stability and hence filopodial lifetime and the probability of microtubule capture. Such a mechanism may underlie vectorial growth because it would allow material moving into the growth cone along captured microtubules to be incorporated differentially into a particular region of the growth cone. To test this hypothesis we have used our in vitro choice assay.
If our hypothesis is correct, we might expect the turning of growth cones at tenascin borders to be mediated by a reorganization of the microtubule-based cytoskeleton. We found that the most obvious change in the growth cone cytoskekton in growth cones at tenascin borders was an asymmetric arrangement of the microtubules, compared with the more evenly distributed, fan-like arrangement of microtubules in growth cones on laminin. To test the functional role of microtubules in growth cone turning we used substoichiometric (nanomolar) concentrations of Taxol that can reversibly block microtubule dynamics in cells without altering their numbers (20, 31, 32). We found that growth cones in the presence of Taxol were unable to turn and grow along tenascin borders, supporting the idea that the dynamic instability and rearrangement of growth cone microtubules is important for growth cone turning.
MATERIALS AND METHODS
Preparation of Proteins for Substrate Coating.
Tenascin was prepared from the brains of 1- to 15-day-old mice by immunoaffinity chromatography (11, 33). Laminin was purchased from Boehringer Mannheim or Sigma. Colloidal gold-labeled bovine serum albumin (BSA) was prepared as described (5, 6).
Substrate Preparation.
The preparation of sharp substrate borders of laminin and tenascin has been described in detail elsewhere (5, 6). Briefly, colloidal gold-labeled BSA was applied to the middle of glass coverslips, incubated at 37°C for 30 min, and washed off with Ca2+- and Mg2+-free PBS. This produced a visible spot with phase-contrast microscopy. Tenascin (50 μg/ml) or, as a control, heat-inactivated, fatty acid-free BSA (Boehringer Mannheim; 50 μg/ml in PBS) was applied directly on top of the colloidal gold-labeled BSA spot. After 2 h at 37°C, the tenascin or BSA-coated spot was washed with PBS, after which the entire coverslip was coated with laminin (20 μg/ml in PBS) at 37°C for 2 h. Coverslips were washed with Ham’s F-14 or DMEM and then left in the CO2 incubator, covered with culture medium (see below), until use.
Tissue Culture.
DRGs were dissected from the lumbrosacral region of 7- or 8-day old chicken embryos and explanted about 1 mm from the border. After the explants had attached to the substrate they were flooded with culture medium (DMEM; GIBCO) supplemented with 10% fetal calf serum (GIBCO), penicillin (100 units/ml), streptomycin (100 μg/ml), and 7S nerve growth factor (50 ng/ml; Boehringer Mannheim). Cultures were maintained for 8–24 h before fixation (see below), after which time growth cones had arrived at the tenascin border. In some experiments, cultures were maintained in the presence of Taxol, at concentrations between 10 and 75 nM. Taxol was stored at −20°C as a stock solution in dimethyl sulfoxide (DMSO). For each experiment, DMSO was added to some cultures as control.
Video Time-Lapse Microscopy.
Video time-lapse microscopy was used to record growth cone behavior. Growth cones were considered to have reached the border when more than one filopodium had contacted the tenascin. Cultures were viewed under an inverted microscope (Axiovert; Zeiss) fitted with an oil-immersion objective (×100) and maintained at 37°C in humidified air containing 5% CO2 inside a climatized box surrounding the microscope stage (Zeiss). Images were recorded using a silicon intensified (SIT) video camera, at defined intervals (typically one image every 2–3 sec) which were determined by means of a pacemaker (built in the laboratory of F. Bonhoeffer, Tübingen, Germany). After image processing on an arithmetic frame grabber (Data Translation, Marlboro, MA) using opto-tech software, images were stored on video tape using a Grundig (Fürth, Germany) video recorder modified for video time-lapse recording. In some experiments the cultures were photographed on Ilford HP5 film.
Antibodies.
Rat mAb YL 1/2, specific for C terminally tyrosinated α-tubulin (34, 35) was purchased from Sera-Lab (Crawley Down, Sussex, U.K.). Rabbit antiserum against detyrosinated α-tubulin (SUP GLU; ref. 36) was a gift from J. C. Bulinski (Columbia University, New York). Anti-actin antibody (N350) and anti-α-tubulin antibody (N356) were purchased from Amersham. Secondary antibodies (fluorescein or rhodamine anti-rat and anti-rabbit) were purchased from Sigma or Milan Analytical (LaRoche, Switzerland). Rhodamine or fluorescein-labeled phalloidin was purchased from Molecular Probes.
Fixation and Double Labeling for Tubulin and F-Actin.
After 8–14 h of culture, or following video time-lapse microscopy, cultures were fixed by removing most of the culture medium while maintaining the cultures at 37°C, and replacing it with 3% (wt/vol) formaldehyde, 0.2% (vol/vol) glutaraldehyde, 0.2% Triton X-100, and 10 mM EGTA in PBS (26) at 37°C for 15 min. Cultures were washed five times with PBS, incubated in blocking buffer (5% horse and goat serum/50 mM l-lysine/0.2% Triton X-100 in PBS), for 1 h, followed by YL 1/2 (1:2), SUP GLU (1:1000), N350 (1:300), or N356 (1:300) alone or in combination in blocking buffer for 2 h, washed with PBS, and incubated with the appropriate secondary antibody (1:100) and rhodamine or fluorescein-conjugated phalloidin (5 μg/ml) in blocking buffer for 30 min. Controls omitted the primary antibody or the two secondary antibodies were exchanged to give the inappropriate combination. All controls were negative. Coverslips were mounted in Citifluor (City University, London) and viewed with phase contrast or fluorescence optics using an Olympus BX50 microscope and photographed on Kodak T-Max 400 film.
Image Analysis.
To quantify the arrangement of microtubules within growth cones, growth cones that had been fixed and stained for actin and tubulin were bisected with a straight line that divided the area of the growth cone into two equal halves using the ibas image analysis system (Kontron Instruments, Zurich). The number of microtubules terminating in each half of the growth cone were counted. An asymmetric distribution of microtubules within the growth cone was scored when >60% of microtubules terminated in one half of the growth cone.
RESULTS
Growth Cone Behavior on Laminin and at Tenascin Borders.
Chicken DRG growth cones were examined by video time-lapse microscopy to assess their behavior before fixation and to monitor the fixation process. Growth cones on laminin, which had not arrived at tenascin borders, had a similar morphological appearance and behavior to that described previously (6). They were well spread and displayed prominent lamellipodia and several filopodia (between 5 and 20 per growth cone) which extended and retracted from the growth cone margin. The average rate of growth cone advance on laminin measured in living cultures was 55.2 ± 5.6 μm/h (mean ± SD, n = 5). After addition of fixative to the cultures, growth cone motility ceased within a few seconds, and growth cone morphology remained relatively unchanged during the period of fixation (data not shown; this is in agreement with ref. 21).
Growth cones at tenascin borders had a much slower rate of advance than those on homogeneous laminin substrates. In most cases (≈90%), growth cones did not cross into the tenascin-containing territory, and growth cone filopodia that did were rapidly withdrawn within seconds. The lamellipodia of growth cones at tenascin borders tended to collapse away from the border (6). However, in some cases the morphology of growth cones at tenascin borders appeared little different to those on laminin.
Microtubule Organization in Growth Cones on Laminin and at Tenascin Borders.
The organization of microtubules in growth cones was monitored by labeling fixed growth cones with tubulin antibodies. We used mAb YL 1/2, which recognizes C terminally tyrosinated α-tubulin, because this form of tubulin is the major form in growth cones (37, 38). The microtubule organization in chicken DRG growth cones on laminin as revealed by staining with mAb YL 1/2 was similar to that described previously (26). Microtubules are bundled within the axon, but on entering the growth cone they splay out within the central domain. In some cases microtubules reached into the peripheral domain of the growth cone, occasionally entering filopodia (Fig. 1 A and B) (26).
Figure 1.
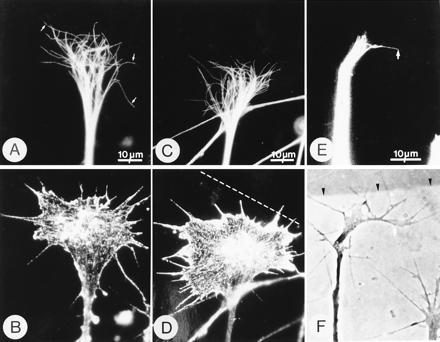
Microtubules become reoriented in growth cones at tenascin borders. Phase-contrast and immunofluorescence micrographs of growth cones immunostained with mAb YL 1/2, which stains microtubules (A, C, and E) and mAb N350, which stains actin (B and D). (F) Phase-contrast image of the living growth cone shown in E. In growth cones growing on laminin (A and B), microtubules are splayed out in the central domain and extend, individually, far into the peripheral domain and occasionally insert into filopodia (arrows in A). In this growth cone, 52% of the microtubules had terminations in the left-hand area of the growth cone and therefore, by the definition given in Materials and Methods, the distribution is symmetrical. However, in growth cones at laminin-tenascin borders, microtubules become asymmetrically distributed (C and E). In C and D, the actin staining (D) reveals the growth cone morphology which suggests that the axon approached the border at a slight angle and that the right hand side of the growth cone has contacted the border, indicated by the broken line. The microtubules in this growth cone (C) are asymmetrically distributed (65% of the microtubules terminate in the right-hand half). In micrographs E and F the growth cone has clearly turned to the right and the microtubule asymmetry is obvious (arrow in E). Arrows in E and F are at the same location. The laminin–tenascin border is visualized with colloidal gold (arrowheads).
In growth cones at tenascin borders the microtubules became reorganized so that their distal terminations were no longer symmetrically distributed within the growth cone (Fig. 1 C–F). We found examples where microtubules were asymmetrically arranged (Fig. 1C) and the growth cone appeared to have initiated a turn, as judged by phase contrast microscopy and the fact that the actin staining indicated that the mass of the growth cone was unevenly distributed (Fig. 1D). We did not find any clear-cut examples where microtubules were asymmetrically distributed but the growth cone was symmetrical. However, we may have missed such an event because growth cones become asymmetrical soon after contacting borders. In other cases, the growth cone was clearly turning as seen by phase contrast microscopy (Fig. 1 E and F). However, in fixed cultures, predicting growth cone behavior at borders, particularly in which direction the growth cone will turn, is not always possible. Furthermore, we do not have a direct way of precisely determining when a growth cone has decided to turn. For these reasons we cannot say whether microtubule reorganization precedes growth cone turning.
Quantification of the arrangement of microtubules within growth cones on laminin revealed that the vast majority (82%; 37/45 growth cones analyzed) displayed a symmetrical arrangement of microtubules. In contrast, in growth cones at tenascin borders, only 34% (24/70 growth cones analyzed) displayed a symmetrical microtubule arrangement. In this analysis we counted between 10 and 40 microtubules per growth cone on laminin and at the border. These results suggest that in living growth cones turning at the tenascin border there is a correlation with reorganization of the microtubules in fixed growth cones that display signs of turning.
Growth Cone Behavior During Taxol Treatment.
The results described above suggest that microtubule reorganization in growth cones at tenascin borders might be an early consequence of growth cone contact with tenascin and hence underlie growth cone turning. To test this hypothesis, we examined the behavior of growth cones at tenascin borders in the presence of the microtubule stabilizing compound, Taxol. Previous work has shown that stoichiometric concentrations (micromolar) of Taxol inhibits axon growth and increases microtubule mass and bundling in growth cones (20, 38–43). More recent work with substoichiometric concentrations (nanomolar) of Taxol has revealed more subtle effects in nonneuronal cells; principally a reduction in dynamic instability without an appreciable increase in polymer mass (31, 32). We wondered whether substoichiometric concentrations of Taxol might prove useful in probing microtubule function in growth cones turning at tenascin borders.
First we determined the effects of low concentrations of Taxol on the organization of microtubules in growth cones growing on laminin and at tenascin borders (Fig. 2). To do this we added Taxol, at concentrations of 10 and 75 nM, to DRG cultures and prepared them for immunofluorescence at various times after exposure to the drug. Because Taxol at high concentrations is known to stabilize microtubules, and this is associated with an increase in the proportion of microtubules containing detyrosinated α-tubulin in growth cones (43), we used an antibody (SUP GLU) that recognizes this form of α-tubulin as well as mAb YL 1/2, which recognizes tyrosinated α-tubulin. This approach ensured that all microtubules in growth cones were observed. In control cultures, the majority of the microtubules in growth cones contained tyrosinated α-tubulin (i.e., were YL 1/2+), confirming previous observations (Fig. 2A; refs. 26 and 44). There is a set of microtubules that contained detyrosinated α-tubulin (SUP GLU+), but these were less numerous, more proximally located in the growth cone and tended to be highly curved, often describing hair-pin loops (Fig. 2B). After Taxol treatment, the tyrosinated microtubules were fewer in number, did not extend as far distally (Fig. 2 C and D) as they normally did in control cultures (Fig. 1 A and B and Fig. 2 A and B) and were tightly bundled (Fig. 2 D, E, and G). In Taxol-treated growth cones, the detyrosinated microtubules were either highly curved or, more usually, tightly bundled, and extended as far forward as the tyrosinated microtubules (compare Fig. 2 F and H with E and G). Furthermore, their numbers had increased (Fig. 2 F and H compared with Fig. 1B). To quantitate the microtubule bundling effect of Taxol, we measured the lateral spread of microtubules in growth cones from micrographs. In control growth cones growing on laminin, the microtubule spread was 21.07 ± 9.97 μm (mean ± SD, n = 10), whereas after Taxol (75 nM) it was 5 ± 2.28 μm (mean ± SD, n = 18). These effects of Taxol on growth cones were seen with varying severity depending on the concentration of Taxol used. To confirm that the combination of YL 1/2 and SUP GLU double labeling revealed all microtubules we labeled some cultures with an anti-α-tubulin antibody (N356) that recognizes all forms of tubulin and confirmed the Taxol effects on microtubules in growth cones (not shown).
Figure 2.
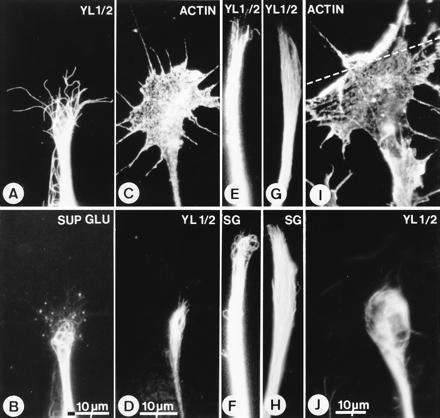
Substoichiometric concentrations of Taxol stabilizes and bundles microtubules. Immunofluorescence micrographs of growth cones grown on laminin (A–H) and at a tenascin border (I and J) in the absence (A and B) and presence of Taxol (C, D, I, and J, 75 nM; E–H, 10 nM). Cultures were immunostained with mAb YL 1/2, which stains tyrosinated α-tubulin (A, D, E, G, and J), SUP GLU (SG in F and H), which stains detyrosinated α-tubulin (B, F, and H), and mAb N350, which stains actin (C and I). The microtubules in growth cones growing on laminin in control cultures are predominantly YL 1/2+—i.e., contain tyrosinated α-tubulin (A)—whereas the SUP GLU+ microtubules form a small, separate group that are often highly curved and more proximally located (B). In the presence of Taxol the microtubules in growth cones become more bundled and do not extend as far distally as normal (C and D; I and J; see A). Furthermore, the proportion of SUP GLU+ microtubules increases (F and H versus E and G). At laminin-tenascin borders, growth cones growing in Taxol are arrested and their microtubules remain bundled and proximally located (I and J). The position of the border is indicated by the broken line. (Magnification in E–H is as in B.)
At Taxol concentrations of 10 and 75 nM, axons were still able to extend from DRG explants, albeit at a reduced speed. In the presence of 10 nM Taxol, the speed in living cultures was 23.8 ± 4.6 μm/hr (mean ± SD, n = 5), which is significantly different (P < 0.001) from control cultures (see above; Figs. 3 and 4). We also determined the effect of Taxol on axon length in fixed cultures. In control cultures, axons had an average length of 144.12 ± 31.03 μm (mean ± SD, n = 100), whereas after 75 nM Taxol the average axon length had decreased to 57.71 ± 17.32 μm (mean ± SD, n = 100). We also noticed that axons became thicker and did not branch in the presence of Taxol (see also ref. 40). In control cultures, 12.26% of axons had branched (n = 302), whereas after 75 nM Taxol the proportion of axons with branches had reduced to 0.82% (n = 491). Video time-lapse microscopy of growth cones showed that there was a transient (10–15 min) response to Taxol in which the lamellipodia partially collapsed and the filopodia became longer (Fig. 4B).
Figure 3.
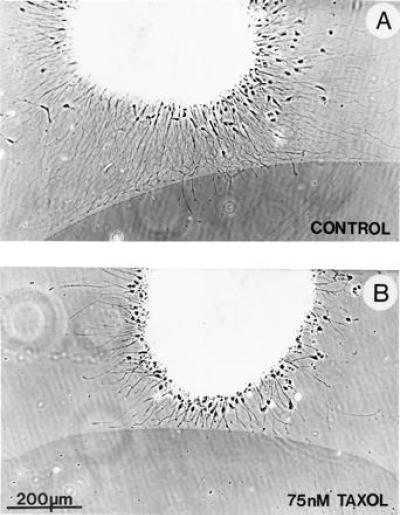
Phase contrast photomicrographs of DRG explants at tenascin borders, grown in the presence of DMSO only (A) or in Taxol (75 nM) in DMSO (B). Taxol, or DMSO, was added at the time of explantation and the cultures were left overnight. The tenascin region, marked by colloidal gold, is the dark area with the sharp border in the lower half of the micrographs. Typically, in the control culture (A), most growth cones turned and grew along the tenascin border. A small proportion of growth cones, particularly those that approached the border perpendicularly, have crossed over. In the presence of Taxol (B), growth cones that approached the border at a shallow angle turned and grew along the border, but those that approached perpendicularly to the tenascin border did not turn or cross. Some of these latter group of growth cones were in contact with the border for at least 5 h. Note that in Taxol (B) the rate of axon elongation is slowed and axon outgrowth appears less profuse, compared with controls (see A).
Figure 4.
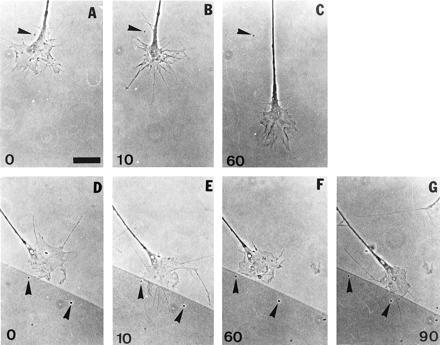
Taxol fails to block growth cone advance on laminin but reversibly blocks growth cone turning at tenascin borders. The first sequence of time-lapse phase-contrast video images (A–C) shows the effects of Taxol (10 nM) on a growth cone advancing on laminin. Numbers indicate time in minutes. Taxol was added shortly after time 0 min (A). There is a transient increase in filopodial length and a partial lamellipodial collapse within 10 min following Taxol addition (B) but growth cone advance is not inhibited; note the stationary piece of debris on the coverslip (arrowheads). In the second sequence (D–G), Taxol was added shortly after time 0 min and replaced with fresh medium, without Taxol, after 60 min. At the tenascin border the growth cone halts its advance following Taxol addition (E and F; note stationary pieces of debris indicated by arrowheads) but resumes turning along the border after the Taxol-containing medium is removed (G). In this sequence also, the growth cone extends several, long filopodia within 10 min following Taxol addition (E) but this phenomenon is transient (F). (Bar in A = 10 μm.) The tenascin covered area is marked by colloidal gold, which appears darker in phase contrast (D–G).
Whereas in the absence of Taxol the majority of growth cones that contacted tenascin borders turned to grow along the border (Fig. 3), in Taxol-treated cultures, many of the growth cones that were at the border were not able to turn, particularly if the approach was perpendicular (Figs. 3 and 4). We cultured 16 DRG explants in normal medium and found that all cultures had examples of growth cones turning at tenascin borders, whereas in 25 DRG explants cultured in the presence of 75 nM Taxol, growth cones failed to turn at the border in all cases. In only two of these explants were growth cones found that had turned. Out of a total of 307 axons that had reached the border, 23 had turned.
Growth cones that had reached the tenascin border in the presence of Taxol were arrested in their advance and remained stationary at the border, although they continued to extend and retract filopodia and lamellipodia. Those growth cones which approached the tenascin border at shallow angles, however, were occasionally able to grow along the border (Fig. 3). When growth cones were unable to turn at the tenascin borders in the presence of Taxol, they remained at the border until the Taxol was removed. We observed individual growth cones arrested at tenascin borders for over 6 h (three growth cones observed), whereas normally they would spend between 30 and 60 min at the border (seven growth cones timed). Thus the reduced growth cone speed seen on laminin in the presence of Taxol (see above) is not sufficient to account for the increase in time that arrested growth cones spend at the border. The microtubules in growth cones arrested at tenascin borders remained tightly bundled and proximally located, as on laminin alone (Fig. 2 I and J). When Taxol was washed out, after maximally 6 h in the culture medium, and replaced by fresh medium, growth cones that had previously arrested at the border were once again able to turn and grow along the laminin side of the border (Fig. 4).
DISCUSSION
Previously we have characterized a growth cone turning event in which chicken DRG growth cones respond to a sharp substrate border between the extracellular matrix proteins laminin and tenascin in vitro (5, 6). At the border, growth cones turn and grow along the laminin side of the border. Growth cone turning is a fundamental event observed during growth cone pathfinding, for example at the optic chiasm (45). Our choice assay provided us with an ideal opportunity to investigate the underlying role of the cytoskeleton in growth cone turning.
Here we show that growth cone microtubules become reorganized in growth cones showing morphological signs of turning at tenascin borders. The reorganization takes the form of a redistribution of the distal terminations of microtubules into one-half of the growth cone. We cannot be completely sure that the reorganization predicts the direction of growth cone turning because we looked at fixed cultures. Recent experiments by Tanaka and Kirschner (46) in living growth cones also indicate that microtubules are reorganized during a turning event. However, they were also unable to say whether microtubule reorganization precedes growth cone turning or is a consequence of it.
To test the role of microtubule reorganization in growth cone turning we applied Taxol, a microtubule stabilizing compound, to growth cones. Previous work has shown that micromolar concentrations of Taxol inhibit axon growth and increase microtubule mass and bundling in growth cones (20, 38–43). More recent work with nanomolar concentrations of Taxol has revealed more subtle effects in nonneuronal cells; principally a reduction in dynamic instability without an increase in polymer mass (31, 32). We found that nanomolar concentrations of Taxol increased microtubule bundling and restricted the distribution of microtubules in growth cones. Microtubule stability in growth cones was also increased, as judged by the increase in the amount of detyrosinated α-tubulin, a marker for microtubule stability (47, 48). Despite these dramatic alterations in microtubule organization in growth cones, axons continued to elongate, albeit at a reduced rate. This is in marked contrast to previous experiments with high concentrations of Taxol—and drugs that depolymerize microtubules—in which axon elongation was inhibited (20, 40, 41, 43, 49). It has been shown that growth cones are the principal site of action of these drugs, and it was proposed that the polymerization of microtubules at the growth cone is essential for axon elongation (49). At high concentrations of Taxol or microtubule depolymerizing drugs it is likely that microtubules are either inhibited from extending forward into the growth cone, by sliding or polymerization, or are completely lost from the growth cone. In our experiments with nanomolar concentrations of Taxol, although microtubules are still present in growth cones, microtubule polymerization is probably severely reduced or even abolished (31). Since axon elongation continued under such conditions, it is likely that elongation is not entirely dependent upon microtubule polymerization, as originally suggested (49). Alternatively, axon elongation might be sustained both by microtubules extending into the growth cone by polymer sliding and by polymerization (50).
When growing in the presence of Taxol, growth cones at tenascin borders were not able to turn and grow along the laminin–tenascin border, and consequently halted their advance. Growth cones were arrested at borders for as long as Taxol was present (up to 6 h) and their microtubules remained bundled and proximally located. These effects of Taxol were completely reversible. Together, these results suggest that microtubule reorganization in growth cones is a necessary event in growth cone turning. Microtubules may function in this important aspect of growth cone behavior to stabilize the turn and also to direct the movement of organelles by fast axonal transport into the appropriate region of the growth cone. We also observed that Taxol abolishes branching of axons, and because this normally occurs by growth cone splitting in chicken DRG cultures (25, 40), this effect may be related to the failure of growth cones to turn at borders in the presence of Taxol.
Experiments in grasshopper embryos in which Ti1 neurons were microinjected with fluorescent-tubulin conjugates suggest that microtubules may either be selectively stabilized or polymerized in regions of the growth cone that will become new axon (51). In these experiments, microtubules selectively invaded branches that developed from stabilized filopodia and eventually became new neurite. Growth cones that contact guidance cues accumulate F-actin at these local contact points and microtubules become redistributed toward these regions afterwards (46, 52, 53). In some cases, microtubule reorganization occurred before growth cone mass became unevenly distributed during turning, suggesting that microtubule reorganization may precede turning. Local stabilization of F-actin may transmit guidance information within the growth cone, possibly by capturing and stabilizing microtubules by microtubule-associated proteins.
What are the signal transduction mechanisms involved in growth cone turning behavior at tenascin borders? Intracellular changes of Ca2+ (54) and the heterotrimeric G proteins (55) have been shown to be involved in complete growth cone collapse (56). These events, however, are different from the partial growth cone collapse seen at tenascin borders (6). The cell surface receptors in the growth cone mediating these different behaviors have yet to be identified. Likely candidates include integrins (57–59) and the F3/11 immunoglobulin superfamily adhesion molecule (60, 61). Knowledge of the receptor and signal transduction mechanisms underlying growth cone turning elicited by tenascin will provide the basis for further understanding of the involvement of the cytoskeleton in response to extrinsic guidance cues.
Acknowledgments
We thank J. C. Bulinski for SUP GLU antibody and Elly Tanaka for very helpful comments on the manuscript. T.W. was supported by a Science and Engineering Research Council studentship and a European Molecular Biology Organization fellowship. The National Cancer Institute generously provided Taxol.
Footnotes
Abbreviations: DMSO, dimethyl sulfoxide; DRG, dorsal root ganglion.
References
- 1.Hynes R O, Lander A D. Cell. 1992;68:303–322. doi: 10.1016/0092-8674(92)90472-o. [DOI] [PubMed] [Google Scholar]
- 2.Goodman, C. & Shatz, C. (1993) Cell 72, Suppl., 65–75. [DOI] [PubMed]
- 3.Culotti J G. Curr Opin Genet Dev. 1994;4:587–595. doi: 10.1016/0959-437x(94)90077-g. [DOI] [PubMed] [Google Scholar]
- 4.Tessier-Lavigne M. Curr Opin Genet Dev. 1994;4:596–601. doi: 10.1016/0959-437x(94)90078-h. [DOI] [PubMed] [Google Scholar]
- 5.Taylor J. In: NeuroProtocols, Axon Growth and Guidance in Vitro. Gordon-Weeks P R, editor. Vol. 4. New York: Academic; 1994. pp. 158–166. [Google Scholar]
- 6.Taylor J, Pesheva P, Schachner M. J Neurosci Res. 1993;35:347–362. doi: 10.1002/jnr.490350402. [DOI] [PubMed] [Google Scholar]
- 7.Erickson H P. Curr Opin Cell Biol. 1993;5:869–876. doi: 10.1016/0955-0674(93)90037-q. [DOI] [PubMed] [Google Scholar]
- 8.Saga Y, Yagi T, Ikawa Y, Sakakura T, Aizawa S. Genes Dev. 1992;6:1821–1831. doi: 10.1101/gad.6.10.1821. [DOI] [PubMed] [Google Scholar]
- 9.Steindler D A. Annu Rev Neurosci. 1993;16:445–470. doi: 10.1146/annurev.ne.16.030193.002305. [DOI] [PubMed] [Google Scholar]
- 10.Martini R, Schachner M, Faissner A. J Neurocytol. 1990;19:601–616. doi: 10.1007/BF01257247. [DOI] [PubMed] [Google Scholar]
- 11.Husmann K, Faissner A, Schachner M. J Cell Biol. 1992;116:1475–1486. doi: 10.1083/jcb.116.6.1475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lochter A, Schachner M. J Neurosci. 1993;13:3986–4000. doi: 10.1523/JNEUROSCI.13-09-03986.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Lochter A, Vaughan L, Kaplony A, Prochiantz A, Schachner M, Faissner A. J Cell Biol. 1991;113:1159–1171. doi: 10.1083/jcb.113.5.1159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Wehrle B, Chiquet M. Development (Cambridge, UK) 1990;110:401–415. doi: 10.1242/dev.110.2.401. [DOI] [PubMed] [Google Scholar]
- 15.Wehrle-Haller B, Chiquet M. J Cell Sci. 1993;106:597–610. doi: 10.1242/jcs.106.2.597. [DOI] [PubMed] [Google Scholar]
- 16.Perez R G, Halfter W. Dev Biol. 1993;156:278–292. doi: 10.1006/dbio.1993.1076. [DOI] [PubMed] [Google Scholar]
- 17.Schwab M E, Kapfhammer J P, Bandtlow C E. Annu Rev Neurosci. 1993;16:565–595. doi: 10.1146/annurev.ne.16.030193.003025. [DOI] [PubMed] [Google Scholar]
- 18.Dodd J, Schuchardt A. Cell. 1995;81:471–474. doi: 10.1016/0092-8674(95)90066-7. [DOI] [PubMed] [Google Scholar]
- 19.Yamada K M, Spooner B S, Wessells N K. Proc Natl Acad Sci USA. 1970;66:1206–1212. doi: 10.1073/pnas.66.4.1206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Letourneau P C, Ressler A H. J Cell Biol. 1984;98:1355–1362. doi: 10.1083/jcb.98.4.1355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Forscher P, Smith S J. J Cell Biol. 1988;107:1505–1516. doi: 10.1083/jcb.107.4.1505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Smith S J. Science. 1988;242:708–715. doi: 10.1126/science.3055292. [DOI] [PubMed] [Google Scholar]
- 23.Yamada K M, Spooner B S, Wessells N K. J Cell Biol. 1971;49:614–635. doi: 10.1083/jcb.49.3.614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Bunge M B. J Cell Biol. 1973;56:713–735. doi: 10.1083/jcb.56.3.713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Letourneau P C. J Cell Biol. 1983;97:963–973. doi: 10.1083/jcb.97.4.963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Gordon-Weeks P R. NeuroReport. 1991;2:573–576. doi: 10.1097/00001756-199110000-00005. [DOI] [PubMed] [Google Scholar]
- 27.DiTella F, Feiguin F, Morfini G, Cácares A. Cell Motil Cytoskeleton. 1994;29:117–130. doi: 10.1002/cm.970290204. [DOI] [PubMed] [Google Scholar]
- 28.Bush M S, Goold R, Moya F, Gordon-Weeks P R. Eur J Neurosci. 1996;8:235–248. doi: 10.1111/j.1460-9568.1996.tb01208.x. [DOI] [PubMed] [Google Scholar]
- 29.Mitchison T, Kirschner M. Neuron. 1988;1:761–772. doi: 10.1016/0896-6273(88)90124-9. [DOI] [PubMed] [Google Scholar]
- 30.Gordon-Weeks P R. J Neurocytol. 1993;22:717–725. doi: 10.1007/BF01181317. [DOI] [PubMed] [Google Scholar]
- 31.Jordan M A, Toso R J, Thrower D, Wilson L. Proc Natl Acad Sci USA. 1993;90:9552–9556. doi: 10.1073/pnas.90.20.9552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Wilson L, Jordan M A. In: Microtubules. Hyams J, Lloyd C W, editors. New York: Wiley–Liss; 1994. pp. 59–83. [Google Scholar]
- 33.Faissner A, Kruse J. Neuron. 1990;5:627–637. doi: 10.1016/0896-6273(90)90217-4. [DOI] [PubMed] [Google Scholar]
- 34.Kilmartin J V, Wright B, Milstein C. J Cell Biol. 1982;93:576–582. doi: 10.1083/jcb.93.3.576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Wehland J, Schroder H C, Weber K. EMBO J. 1984;3:1295–1300. doi: 10.1002/j.1460-2075.1984.tb01965.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Gundersen G G, Kalnoski M H, Bulinski J C. Cell. 1984;38:779–789. doi: 10.1016/0092-8674(84)90273-3. [DOI] [PubMed] [Google Scholar]
- 37.Gordon-Weeks P R, Lang R D A. Dev Brain Res. 1988;42:156–160. doi: 10.1016/0165-3806(88)90213-1. [DOI] [PubMed] [Google Scholar]
- 38.Gordon-Weeks P R, Mansfield S G, Curran I. Dev Brain Res. 1989;49:305–310. doi: 10.1016/0165-3806(89)90032-1. [DOI] [PubMed] [Google Scholar]
- 39.Peterson E R, Crain S M. Science. 1982;217:377–379. doi: 10.1126/science.6124041. [DOI] [PubMed] [Google Scholar]
- 40.Letourneau P C, Shattuck T A, Ressler A H. J Neurosci. 1986;6:1912–1917. doi: 10.1523/JNEUROSCI.06-07-01912.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Letourneau P C, Shattuck T A, Ressler A H. Cell Motil Cytoskeleton. 1987;8:193–209. doi: 10.1002/cm.970080302. [DOI] [PubMed] [Google Scholar]
- 42.Gordon-Weeks P R. Neuroscience. 1987;21:977–987. doi: 10.1016/0306-4522(87)90052-2. [DOI] [PubMed] [Google Scholar]
- 43.Mansfield S G, Gordon-Weeks P R. J Neurocytol. 1991;20:654–666. doi: 10.1007/BF01187067. [DOI] [PubMed] [Google Scholar]
- 44.Fan J, Mansfield S G, Redmond T, Gordon-Weeks P R, Raper J A. J Cell Biol. 1993;121:867–878. doi: 10.1083/jcb.121.4.867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Godement P. Semin Dev Biol. 1994;5:381–389. [Google Scholar]
- 46.Tanaka E, Kirschner M W. J Cell Biol. 1995;128:127–137. doi: 10.1083/jcb.128.1.127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Kreis T E. EMBO J. 1986;6:2597–2606. doi: 10.1002/j.1460-2075.1987.tb02550.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Khawaja S, Gundersen G F, Bulinski J C. J Cell Biol. 1988;106:141–149. doi: 10.1083/jcb.106.1.141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Bamburg J R, Bray D, Chapman K. Nature (London) 1986;321:788–790. doi: 10.1038/321788a0. [DOI] [PubMed] [Google Scholar]
- 50.Joshi H C, Baas P W. J Cell Biol. 1993;121:1191–1196. doi: 10.1083/jcb.121.6.1191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Sabry J H, O’Connor T P, Evans L, Toroian-Raymond A, Kirschner M. J Cell Biol. 1991;115:381–395. doi: 10.1083/jcb.115.2.381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Lin C-H, Forscher P. J Cell Biol. 1993;121:1369–1383. doi: 10.1083/jcb.121.6.1369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.O’Connor T P, Bentley D. J Cell Biol. 1993;123:935–948. doi: 10.1083/jcb.123.4.935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Bandtlow C E, Schmidt M F, Hassinger T D, Schwab M E, Kater S B. Science. 1993;259:80–83. doi: 10.1126/science.8418499. [DOI] [PubMed] [Google Scholar]
- 55.Igarashi M, Strittmater S M, Vartanian T, Fishman M C. Science. 1993;259:77–79. doi: 10.1126/science.8418498. [DOI] [PubMed] [Google Scholar]
- 56.Fawcett J W. Trends Neurosci. 1993;16:165–167. doi: 10.1016/0166-2236(93)90141-8. [DOI] [PubMed] [Google Scholar]
- 57.Yokosaki Y, Palmer E L, Prieto A L, Crossin K L, Bourdon M A, Pytela R, Sheppard D. J Biol Chem. 1994;269:26691–26696. [PubMed] [Google Scholar]
- 58.Varnum-Finney B, Venstrom K, Muller U, Kypta R, Backus C, Chiquet M, Reichardt L F. Neuron. 1995;14:1213–1222. doi: 10.1016/0896-6273(95)90268-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Husmann K, Schachner M. Cell Adhes Commun. 1995;3:293–310. doi: 10.3109/15419069509081015. [DOI] [PubMed] [Google Scholar]
- 60.Zisch A H, D’Alessandri L, Ranscht B, Falchetto R, Winterhalter K H, Vaughan L. J Cell Biol. 1992;119:203–213. doi: 10.1083/jcb.119.1.203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Pesheva P, Gennarini G, Goridis C, Schachner M. Neuron. 1993;10:69–82. doi: 10.1016/0896-6273(93)90243-k. [DOI] [PubMed] [Google Scholar]