

Abstract
By examining the front of virus invasion in immature pea embryos infected with pea seed-borne mosaic virus (PSbMV), the selective control of different host genes has been observed. From our observations, the early responses to PSbMV replication can be grouped into three classes, inhibited host gene expression, induced host gene expression, and no effect on a normal host function. The expression of two heat-inducible genes encoding HSP70 and polyubiquitin was induced coordinately with the onset of virus replication and the down-regulation of two other genes encoding lipoxygenase and heat shock cognate protein. The down-regulation was part of a general suppression of host gene expression that may be achieved through the degradation of host transcripts. We discuss the possibilities of whether the induction of HSP70 and polyubiquitin genes represents a requirement for the respective protein products by the virus or is merely a consequence of the depletion of other host transcripts. The former is feasible, as the induction of both genes does result in increased HSP70 and ubiquitin accumulation. This also indicates that, in contrast to some animal virus infections, there is not a general inhibition of translation of host mRNAs following PSbMV infection. This selective control of host gene expression was observed in all cell types of the embryo and identifies mechanisms of cellular disruption that could act as triggers for symptom expression.
For maximum efficiency, a virus must balance the exploitation of host cellular processes against consequential cellular damage. How this is achieved is not well understood. However, examples from a wide range of animal viruses (reviewed in ref. 1) and our earlier work with the plant virus pea seed-borne mosaic virus (PSbMV; ref. 2) suggest that a major component in the control exerted by the virus is the shutoff of transcription and/or translation of host mRNAs. Because the virus must use the host machinery for polynucleotide and protein synthesis, either the control must be highly selective or virus replication must depend on long-lived host mRNAs and/or proteins. A similar argument may also apply to the use of host processes for the correct folding and turnover of viral proteins.
The host shutoff phenomenon has been studied in most detail for poliovirus in which viral proteins interfere with DNA polymerase I, II, and III transcription (3–5) and translation (6, 7) and for herpes simplex virus 1 in which host translation is inhibited and there is a rapid degradation of capped mRNAs by a viral protein with riboexonuclease activity (8). For PSbMV replicating in pea embryonic tissues, there is a transient host shutoff associated with a loss of host gene transcripts associated with diverse areas of metabolism (2).
In common with all viruses, PSbMV replication is associated with the translation of viral RNA and the multifarious activities of virus-encoded proteins. Hence, the basic translational machinery and processes required to assist in the correct folding and turnover of proteins should be in place. We know that viral RNAs are translated and there are animal virus precedents for the presence of protein chaperones (e.g., refs. 9–11) and the ubiquitin protein turnover pathway (12) in infected cells. For plant viruses, there is less information. Closteroviruses encode a 65-kDa protein with strong homology to the HSP70 class of molecular chaperones (13), and ubiquitin has been shown to be associated with the tobacco mosaic virus replicase protein in vivo (14), whereas there is evidence for the activation of the ubiquitin gene in tobacco mosaic virus-infected Nicotiana sylvestris protoplasts (15).
PSbMV (family Potyviridae) has a positive sense RNA genome of 9.8 kb (16). The RNA is polyadenylylated but not capped, having a virus-linked protein (Vpg) at the 5′ end. Potyviral genomes are expressed by translation of a large ORF into a polyprotein that is processed by viral proteases into functional proteins (17). PSbMV is transmitted through seeds, infecting the embryo early in development (18). As the infection proceeds, the virus invades the compact tissues of the cotyledon on an advancing front. Using this system, we correlated the host shutoff phenomenon with active virus replication (2). Here we report an extension to these studies to show that there is a coordinated induction of particular representatives of the chaperone and ubiquitin classes of genes with the down-regulation of other genes. However, there is no detectable effect on rRNA or the translation of host mRNAs. In addition, we show that these effects are not cell-type specific as they occur within the area of PSbMV replication throughout the developing embryo.
MATERIALS AND METHODS
Methods for obtaining PSbMV-infected pea seeds and for their fixation, embedding, and processing for in situ hybridization and immunocytochemistry were as described (2, 18). The localization data were observed consistently over 5–10 experiments using pea embryos harvested at different times of the year.
In Situ Hybridization Probes.
PSbMV +ve and −ve sense RNA in sections of pea tissue were detected using the complementary RNA probes as described (2, 18). Hybridization probes for host RNAs were prepared as follows. (i) A lipoxygenase (lox1:Ps:2) cDNA clone, p320 (19, 20), was the complete cDNA cloned into the SmaI site of pBluescript KS+ (Stratagene). The clone was linearized with KpnI or BamHI for transcription of +ve or −ve sense RNAs with T7 or T3 RNA polymerase, respectively. (ii) HSP70 (PsHSP71.2) cDNA (21) was subcloned as an EcoRI fragment (cDNA nt 1–788) into pBluescript SK+ (Stratagene). The clone was linearized with KpnI or XbaI for transcription of +ve or −ve sense RNAs with T3 or T7 RNA polymerase, respectively. (iii) HSC70 (PsHSC71.0) cDNA (21) was subcloned as an EcoRI fragment (cDNA nt 1–1075) into pBluescript SK+. The clone was linearized with KpnI or BamHI for transcription of +ve or −ve sense RNAs with T3 or T7 RNA polymerase, respectively. (iv) Polyubiquitin (pCU1) cDNA (22) was subcloned as an EcoRI fragment (1.5 kb) into pBluescript SK+. The clone was linearized with HindIII or XbaI for transcription of +ve or −ve sense RNAs with T7 or T3 RNA polymerase, respectively. (v) The 18S rRNA clone (pF4) was a subcloned EcoRI–BamHI fragment (1 kb) from an rDNA clone (pHA2; ref. 23) into pBluescript SK+. The clone was linearized with HindIII or XbaI for transcription of +ve or −ve sense RNAs with T7 or T3 RNA polymerase, respectively.
Digoxygenin-11-UTP-labeled RNA (Boehringer Mannheim) was prepared, hybridized to tissue sections, and detected using an alkaline phosphatase-conjugated anti-digoxygenin antibody as described previously; 5-bromo-4-chloro-3-indoylphosphate and nitro-blue tetrazolium were used as chromogenic substrates (2, 18). Host RNA positive sense probes gave no detectable signal (data not shown).
Processed sections were mounted in Entellan (Merck) without (Figs. 1, 2, 3) or with (Fig. 4) prior to counterstaining in 0.1% Calcofluor (Sigma), and photographed using transmitted light microscopy (Figs. 1, 2, 3) or combined epifluorescence (UV; Calcofluor staining of cell walls) and transmitted light microscopy (Fig. 4).
Figure 1.
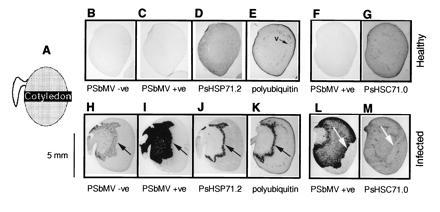
Selective regulation of host gene expression following PSbMV infection of pea cotyledonary tissues. The orientation of the pea cotyledon sections is shown in (A). B–E, F and G, H–K, and L and M were from four separate embryos. Sections B–G and H–M are from healthy and PSbMV-infected cotyledons, respectively. The sections were subjected to in situ hybridization with probes for PSbMV −ve RNA (B and H), PSbMV +ve RNA (C, F, I, and L), PsHSP71.2 RNA (D and J), polyubiquitin RNA (E and K), and PsHSC71.0 RNA (G and M). The zone of HSP70 and polyubiquitin induction correspond with the front of virus invasion and the onset of virus replication (H–K, arrows). In contrast, HSC70 expression is reduced at the front of virus invasion (L and M, white arrows).
Figure 2.
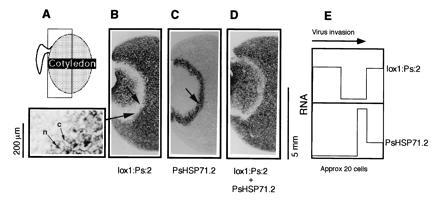
Spatial correlation between the zones of transcript depletion and HSP70 induction. B–D were cut from the same PSbMV-infected cotyledon in the area shown in A; B and C are adjacent sections. Sections were subjected to in situ hybridization with probes for lox1:Ps:2 RNA (B), PsHSP71.2 RNA (C) and both probes together (D). The transcript depletion seen for lox1:Ps:2 appears to involve a rapid loss of transcript from the cytoplasm and the nucleus (B, Inset: compare the signal in the cytoplasm, c, and the nucleus, n, on the right and left of the panel, i.e., outside and inside the infection front, respectively). The zone of PsHSP71.2 induction corresponds with the first few cells of the zone of lox1:Ps:2 RNA depletion (B and C, arrows). When both probes are combined (D) the transcript-depleted zone gets correspondingly narrower. These data are summarized graphically (E).
Figure 3.
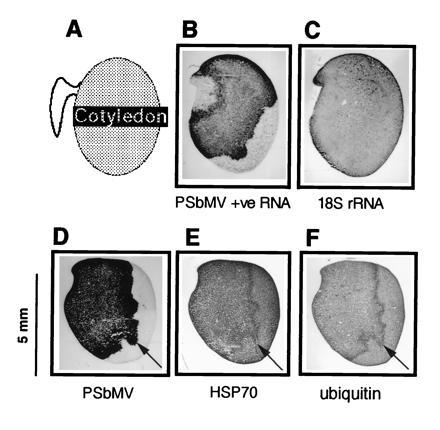
Accumulation of viral and host proteins at the front of virus invasion. B and C and D–F, respectively, were cut from two different infected embryos. The orientation of the sections is illustrated in A. Sections were subjected to in situ hybridization with probes for PSbMV +ve RNA (B) or 18S rRNA (C), or immunohistochemistry with antibodies for PSbMV coat protein (D), HSP70 (E), or ubiquitin (F). rRNA was uniformly distributed throughout the infected cotyledon. Cells at the front of virus invasion (arrows) accumulated viral and induced host proteins.
Figure 4.
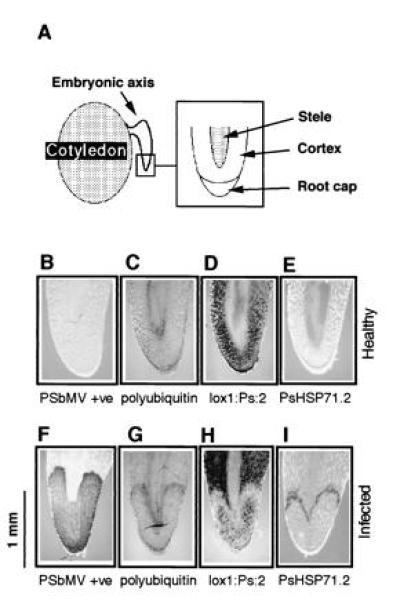
Selective control of host gene expression in the infected embryonic radicle. B–E and F–I were sectioned from single healthy and infected embryos, respectively, as illustrated in A. Sections were subjected to in situ hybridization with probes for PSbMV +ve RNA (B and F), polyubiquitin RNA (C and G), lox1:Ps:2 RNA (D and H) or PsHSP71.2 RNA (E and I). As observed in cotyledons, coordinate induction of polyubiquitin and HSP70 and down-regulation of lipoxygenase expression were observed. Note, to visualize the tissue structure in the absence of hybridization signal, these sections were photographed under joint-transmitted and epifluorescent light after staining the tissue sections with calcofluor.
Antisera.
Polyclonal rabbit antisera for HSP70 homologs (21) and ubiquitin (Sigma) were used at dilutions of 1/100 and 1/250, respectively. The immunohistochemical procedures with anti-HSP70, anti-ubiquitin, and anti-PSbMV coat protein (18) were as described previously except that antibody incubations were carried out at 4°C overnight.
RESULTS
Detection of HSP70 and Polyubiquitin RNA in Healthy and PSbMV-Infected Cotyledons.
HSP70 represents one of the families of heat-inducible chaperones. Three cytoplasmic HSP70 homologs have been identified in pea (21). PsHSP71.2 is heat-inducible, nonconstitutive in leaves, but constitutively expressed late in seed development. PsHSC71.0 is constitutive and nonheat inducible in leaves, and is expressed continuously during seed development. PsHSP70.b represents an intermediate situation, being constitutive and heat inducible in leaves (21). Ubiquitin is a highly conserved small (8.5 kDa) polypeptide that is intimately involved in targeting protein for degradation, but has other cellular roles, including responses to a variety of biological and physical stress conditions (reviewed in ref. 24). Ubiquitin is encoded by a gene family of natural gene fusions, termed polyubiquitin, for which two distinct cDNA clones (pCU1 and pCU2) have been isolated from pea (22). There are no data for the patterns of expression of these genes in pea embryos.
Using in situ hybridization of embryo sections, we have studied the expression of PsHSP71.2, PsHSC71.0, and a polyubiquitin gene (pCU1) in healthy and PSbMV-infected pea cotyledons. Near-consecutive sections were also treated with strand-specific probes for PSbMV RNA. In sections of healthy cotyledons, host PsHSP71.2 transcripts are detected weakly and with a uniform distribution (Fig. 1D). When sections were probed for polyubiquitin or PsHSC71.0 RNAs, a stronger hybridization signal was observed, particularly in the provascular tissues and surface layers of the cotyledon (Fig. 1 E and G). In virus-infected cotyledons, a different pattern of expression was seen. For PsHSP71.2 and polyubiquitin, there was a marked zone of increased transcript accumulation (Fig. 1 J and K). This zone, which was two to four cells wide, lay close to the front of virus invasion, as revealed by probing for PSbMV +ve RNA (Fig. 1I), and appeared to overlie the zone of viral RNA replication (Fig. 1H). In contrast, the weak expression of PsHSC71.0 was marginally reduced within the zone of viral RNA replication (Fig. 1M) in a pattern similar to that seen previously for suppressed host gene expression (2). This reduction was less dramatic than that seen for other genes but was consistently observed in all the experiments with the PsHSC71.0 probe. The expression of PsHSP71.2 and polyubiquitin showed further differences from the other genes and from each other. Whereas previously suppression of host gene expression was followed by a complete recovery to levels seen in healthy tissue as viral RNA replication declined (ref. 2; Fig. 1M), expression of PsHSP71.2 was abolished after the induction phase behind the infection front (Fig. 1, compare D and J) and was only sometimes seen to recover a long time after the wave of infection had passed (i.e., close to the embryonic axis; not shown). In contrast, polyubiquitin showed no significant reduction in expression over that in healthy tissue after the induction phase except perhaps in the provascular tissue (Fig. 1, compare E and K).
Comparison of the Spatial Patterns of PsHSP71.2, lox1:Ps:2, and PSbMV RNA Accumulation.
The expression of genes associated with seed storage protein synthesis and starch metabolism (2) and PsHSC71.0 (Fig. 1M) was suppressed in cells replicating PSbMV RNA. The pea lipoxygenase gene, lox1:Ps:2 (19, 20), was further selected as a gene expressed to intermediate levels in the middle to late stages of pea embryo development, and as a gene representative of a different area of host metabolism. When sections of virus-infected pea cotyledons were probed for lox1:Ps:2 expression, a transcript-depleted zone (Fig. 2B) that correlated with the area of viral RNA replication (not shown) was seen. As observed previously (2), the front of virus invasion, indicated by transcript depletion, was seen as a sharp demarcation between uninfected and infected areas at which point host mRNAs disappeared throughout the cell. In the case of lox1:Ps:2, this included a loss of RNA from nuclei and from the cytoplasm (Fig. 2B, Inset).
The induction of genes in infected tissues could occur as a response to a signal in advance of the infection front, to the onset of viral replication, or to the accumulation of viral products in the cells. To determine more precisely the spatial relationship between the suppression and induction phenomena, consecutive sections (15 μm thick; cotyledonary cells have an approximate size of 50 μm) were analyzed. In Fig. 1, the sections shown in panels H–K are strictly consecutive. Measurement of the position of the induction and the virus-infected zone (Fig. 1, arrows) showed that the induction zone was at the extreme outer edge but overlying the infected area indicative of a response to the onset of virus replication. To assess the relationship between the induction and suppression of gene expression, consecutive sections (Fig. 2 B and C) were analyzed for lox1:Ps:2 and PsHSP71.2, respectively. This showed that these different response zones also coincided at the outer edge of virus invasion. These data were supported when a near-consecutive section was analyzed jointly for both mRNAs. In this case, the area with reduced hybridization signal was narrower than with lox1:Ps:2 mRNA alone (Fig. 2D). These data are summarized graphically in Fig. 2E.
Detection of HSP70 and Ubiquitin in PSbMV-Infected Cotyledons.
The translation of viral RNAs would require that the host translational machinery should remain intact even against a background of host mRNA degradation. Accordingly, when sections of infected cotyledons were probed for the presence of 18S rRNA, no difference in the accumulation of rRNA was observed between infected and uninfected areas (Fig. 3 B and C). It is possible however that in the area of viral RNA replication, translation could be restricted to viral RNAs, possibly through an interference from viral proteins in the translation of host capped mRNAs. A consequence of this would be that the induced PsHSP71.2 and polyubiquitin mRNAs might not result in an increased accumulation of protein. To test this, sections of infected cotyledons were analyzed for protein accumulation using immunocytochemistry and the respective antibodies. Since these genes are members of large gene families and the antibodies recognize many of the protein products, the level of antibody binding is high in the uninfected and infected regions in both cases (Fig. 3 E and F). Nevertheless, the increased accumulation of the respective proteins at the front of virus invasion (Fig. 3D) is clear (arrows).
Expression of PsHSP71.2, Polyubiquitin, and lox1:Ps:2 in the Embryonic Axis.
In our earlier experiments, we used sections of pea cotyledons to analyze host responses to virus replication. The cotyledon is a specialized organ composed predominantly of cortical cells dedicated to the accumulation of storage products. To test whether the responses to virus replication were cell-type specific, parts of the embryonic axis were subjected to the same in situ analysis. The embryonic axis is a highly complex structure with cell types representative of all the vegetative components of the mature plant. Fig. 4 shows the data obtained for sections through the embryonic radicle; equivalent results were obtained for the hypocotyl and epicotyl regions (not shown).
Sections of the radicle from uninfected seeds probed for lox1:Ps:2, PsHSP71.2, or polyubiquitin expression showed that lox1:Ps:2 was expressed in the root cortex and root cap but barely at all in the stele (Fig. 4D), polyubiquitin was expressed in the cortex and the stele (Fig. 4C), and PsHSP71.2 was not expressed in the radicle at all (Fig. 4E). In contrast, sections of the radicle from infected embryos showed very different patterns of expression (Fig. 4 G–I). Because of the narrow dimension of the tip of the radicle, it is not possible to correlate precisely the spatial distribution of the expression. However, the general features of the responses seen in cotyledons remain the same in the radicle. Virus invasion into the embryo follows a precise pathway, originating at the tip of the radicle from where it is subsequently excluded, to become reinfected later in embryo development (unpublished data). Following reinfection, the virus spreads through the cortical tissues of the radicle and more slowly through the stele (Fig. 4F). Correlated with the front of virus invasion, there is decreased expression of lox1:Ps:2 (Fig. 4H) and increased expression of polyubiquitin and PsHSP71.2 (Fig. 4 G and I). The increased expression was seen in both the cortex and the stele and was especially notable for PsHSP71.2, for which there is no constitutive basal expression (Fig. 4I).
DISCUSSION
We showed previously that virus replication was associated with a depletion of host transcripts for nine genes associated with two distinct areas of metabolism, seed storage protein accumulation and starch metabolism. The observations have been extended in this work to include lipoxygenase and a constitutively expressed putative chaperone, HSC70. The abrupt loss of in situ hybridization signal at the front of virus invasion suggests that host RNAs are degraded in the cytoplasm and nucleus and that there is a block to further accumulation until virus replication declines. This block could result from continued efficient degradation or from an inhibition of transcription. If from an inhibition of transcription, then this could be under direct virus control, as observed for poliovirus, or could result from a loss of translation products essential for transcription. It is not clear at present which of these possibilities applies.
Not all host genes are down-regulated during virus replication. We have identified two genes (encoding HSP70 and polyubiquitin) that are induced, but it is likely that there will be others. These three phenomena associated with PSbMV replication (host mRNA depletion, induction of HSP70 and induction of polyubiquitin) have been observed before for some animal viruses but not for plant viruses, and never have all three phenomena been studied in the same infection. This system provides special opportunities not only to characterize the influence of the virus on host gene expression but also to study the coordinate expression of HSP70 and polyubiquitin in a background of mRNA degradation.
Our data show that these changes in host gene expression coincide with the onset of virus replication. The changes can be interpreted in three ways: (i) as positive controls by the virus to avoid competition for macromolecular synthesis; (ii) as part of an unsuccessful defense response; or (iii) as a series of separate events where one or more is consequential upon the effect of another. In the first case, it could be argued that the selective regulation of different genes could be beneficial to viral protein function and hence to virus accumulation. The second option will need to await more detailed observations on the success of virus invasion when these phenomena are either enhanced or diminished. The third point is interesting in the context of the control of gene expression for HSP and polyubiquitin.
The complex control of HSP expression has been reviewed recently (25). Briefly, the current model suggests that in the absence of heat induction, the HSP gene has a paused transcriptional complex. Transcriptional induction is mediated by the binding of a transcriptional activator, heat shock factor, to a short DNA regulatory sequence, the heat shock element. The availability and activity of heat shock factors is controlled by the formation of complexes with HSP, and by phosphorylation. The diversion of HSP into chaperone activity to correct heat shock damage to cellular proteins releases heat shock factor in trimeric form for HSP transcriptional activation. Thus, HSP can negatively control its own expression. The upstream region of eukaryotic polyubiquitin genes also contain heat shock elements (e.g., ref. 23), and, following heat treatment, polyubiquitin and HSP expressions are both induced. Heat-induced stress also results in the degradation of host mRNAs against which HSP (and presumably polyubiquitin) RNAs are protected (26). Hence, many of the features of the response to PSbMV replication mimic those resulting from heat stress.
These observations raise many questions. Particularly, in avoiding competition in the cell through a host shutoff mechanism does the virus trigger incidentally the induction of heat responsive genes, albeit to advantage? This could occur through a block to host protein synthesis since induced HSP expression has been observed when protein synthesis is inhibited (27). The abrupt induction of HSP at the infection front gives no indication of a time delay between it and mRNA depletion. The equally abrupt stop to HSP expression within the zone of mRNA depletion presumably results from negative feedback regulation from HSP itself. The coordinate induction of HSP70 and polyubiquitin supports the idea that they may both be regulated through heat shock elements. However, there must be differences in the subtleties of their regulation as there were differences in the pattern of recovery of basal expression after virus replication had ceased. For HSP70, expression declined and failed to recover to the constitutive level seen in uninfected tissue whereas polyubiquitin expression rapidly returned to “normal” levels.
Although PSbMV behaves like many animal viruses in promoting a host shutoff mechanism, it differs from poliovirus in particular in not blocking the translation of host mRNAs, or at least those representative of heat-responsive genes. It would be interesting to test whether HSP and polyubiquitin were induced following poliovirus infection. The absence of induction would suggest that the gene products were not always essential during virus replication. Recently, evidence was obtained to indicate that induction of members of the HSP70 family in cells infected with canine distemper virus could enhance viral polymerase activity (28).
The observation that PSbMV host shutoff and heat shock protein induction occurred throughout the developing embryo supports the idea that this is a general mechanism rather than one peculiar to the specialized cell type within the cotyledon. Why then have these phenomena not been observed before? The infection of plants by viruses is a completely asynchronous process and observed phenomena are very transient, probably only a few hours within a single cell. The only plant single cell system available for synchronized infections are plant protoplasts. These are subjected to osmotic trauma and the toxic effects of relatively crude cell-wall degrading enzymes and, at best, only approximate a synchronous response. The mechanisms underlying symptom expression in virus infected plants are very poorly understood. The magnitude of the responses to virus replication observed in this work could represent the primary triggers for a sequence of events that lead to symptom production. We hope that by manipulating the growth conditions of the plant (e.g., by differential temperature treatment; ref. 29) we can obtain sufficient synchrony for a more detailed analysis of the molecular events governing these responses.
Acknowledgments
We thank R.Casey for plox1:Ps:2; F. Watts for pCU1; P. Shaw for pF4; and E. Vierling for pPsHSP71.2, pPsHSC71.0, and HSP70 polyclonal antibodies. We also thank M. Boulton, J. W. Davies, and R. Casey for reviewing the manuscript before submission. The work was supported by the Ministerio de Educacion y Ciencia, Spain (Fellowship to M.A.), the Biotechnology and Biological Research Council (D.W.), and the John Innes Foundation (M.E.).
Footnotes
Abbreviations: PSbMV, pea seed-borne mosaic virus; +ve RNA, encapsidated +ve sense viral RNA; −ve RNA, replicative −ve sense viral RNA (nonencapsidated).
References
- 1.Knipe D M. In: Fundamental Virology. 3rd Ed. Fields B N, Knipe D M, Howley P M, editors. Philadelphia: Lippincott–Raven; 1996. pp. 239–265. [Google Scholar]
- 2.Wang D, Maule A J. Science. 1995;267:229–231. doi: 10.1126/science.267.5195.229. [DOI] [PubMed] [Google Scholar]
- 3.Rubinstein S J, Hämmerle T, Wimmer E, Dasgupta A. J Virol. 1992;66:3062–3068. doi: 10.1128/jvi.66.5.3062-3068.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Yalamanchili P, Harris K, Wimmer E, Dasgupta A. J Virol. 1996;70:2922–2929. doi: 10.1128/jvi.70.5.2922-2929.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Clark M E, Hämmerle T, Wimmer E, Dasgupta A. EMBO J. 1991;10:2941–2947. doi: 10.1002/j.1460-2075.1991.tb07844.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Novoa I, Cotten M, Carrasco L. J Virol. 1996;70:3319–3324. doi: 10.1128/jvi.70.5.3319-3324.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Irurzun A, Sánchez-Palomino S, Novoa I, Carrasco L. J Virol. 1995;69:7453–7460. doi: 10.1128/jvi.69.12.7453-7460.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Zelus B D, Stewart R S, Ross J. J Virol. 1996;70:2411–2419. doi: 10.1128/jvi.70.4.2411-2419.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Sedger L, Ruby J. J Virol. 1994;68:4685–4689. doi: 10.1128/jvi.68.7.4685-4689.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Yang J, DeFranco D B. Mol Cell Biol. 1994;14:5088–5098. doi: 10.1128/mcb.14.8.5088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hu J, Seeger C. Proc Natl Acad Sci USA. 1996;93:1060–1064. doi: 10.1073/pnas.93.3.1060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Latchman D S, Estridge J K, Kemp L M. Nucleic Acids Res. 1987;15:7283–7293. doi: 10.1093/nar/15.18.7283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Dolja V V, Karasev A V, Koonin E V. Annu Rev Phytopathol. 1994;32:261–285. [Google Scholar]
- 14.Gaspar J O, Dunigan D D, Zaitlin M. Mol Plant–Microbe Interact. 1990;3:182–187. [Google Scholar]
- 15.Genschik P, Parmentier Y, Durr A, Marbach J, Criqui M C, Jamet E, Fleck J. Plant Mol Biol. 1992;20:897–910. doi: 10.1007/BF00027161. [DOI] [PubMed] [Google Scholar]
- 16.Johansen E, Rasmussen O F, Heide M, Borkhardt B. J Gen Virol. 1991;72:2625–2632. doi: 10.1099/0022-1317-72-11-2625. [DOI] [PubMed] [Google Scholar]
- 17.Lindbo J A, Dougherty W G. In: Encyclopedia of Virology. Webster R G, Granoff A, editors. Vol. 3. New York: Academic; 1994. pp. 1148–1153. [Google Scholar]
- 18.Wang D, Maule A J. Plant Cell. 1994;6:777–787. doi: 10.1105/tpc.6.6.777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Forster C, Knox M, Domoney C, Casey R. Plant Physiol. 1994;106:1227–1228. doi: 10.1104/pp.106.3.1227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Domoney C, Firmin J L, Sidebottom C, Ealing P M, Slabas A, Casey R. Planta. 1990;181:35–43. doi: 10.1007/BF00202322. [DOI] [PubMed] [Google Scholar]
- 21.DeRocher A, Vierling E. Plant Mol Biol. 1995;27:441–456. doi: 10.1007/BF00019312. [DOI] [PubMed] [Google Scholar]
- 22.Watts F Z, Moore A L. Nucleic Acids Res. 1989;17:10100. doi: 10.1093/nar/17.23.10100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Jorgensen R A, Cuellar R E, Thompson W F, Kavanagh T A. Plant Mol Biol. 1987;8:3–12. doi: 10.1007/BF00016429. [DOI] [PubMed] [Google Scholar]
- 24.Monia B P, Ecker D J, Crooke S T. Biotechnology. 1990;8:209–215. [Google Scholar]
- 25.Mager W H, De Kruijff A J J. Microbiol Rev. 1995;59:506–531. doi: 10.1128/mr.59.3.506-531.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Pitto L, Gallie D R, Walbot V. Plant Physiol. 1992;100:1827–1833. doi: 10.1104/pp.100.4.1827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ovelgonne J H, Souren J, Wiegant F, Vanwijk R. Toxicology. 1995;99:19–30. doi: 10.1016/0300-483x(94)02990-c. [DOI] [PubMed] [Google Scholar]
- 28.Oglesbee M J, Liu Z, Kenney H, Brooks C L. J Gen Virol. 1996;77:2125–2135. doi: 10.1099/0022-1317-77-9-2125. [DOI] [PubMed] [Google Scholar]
- 29.Dawson W O, Schlegel D E, Lung M C Y. Virology. 1975;65:565–573. doi: 10.1016/0042-6822(75)90061-6. [DOI] [PubMed] [Google Scholar]