

Abstract
Controlled protein degradation mediated by ubiquitin/proteasome system (UPS) plays a crucial role in modulating a broad range of cellular responses. Dysregulation of the UPS often accompanies tumorigenesis and progression. Here, we report that Smad ubiquitination regulatory factor 2 (Smurf2), a HECT-domain containing E3 ubiquitin ligase, is up-regulated in certain breast cancer tissues and cells. We show that reduction of Smurf2 expression with specific short interfering RNA in metastatic breast cancer cells induces cell rounding and reorganization of the actin cytoskeleton, which are associated with a less motile and invasive phenotype. Overexpression of Smurf2 promotes metastasis in a nude mouse model and increases migration and invasion of breast cancer cells. Moreover, expression of Smurf2CG, an E3 ligase–defective mutant of Smurf2, suppresses the above metastatic behaviors. These results establish an important role for Smurf2 in breast cancer progression and indicate that Smurf2 is a novel regulator of breast cancer cell migration and invasion.
Introduction
Regulated protein degradation by the ubiquitin/proteasome system (UPS) is often a critical control step in a variety of cellular functions (1). This process is initiated by protein ubiquitination, which is carried out by three sequential enzymes: ubiquitin-activating enzyme (E1), ubiquitin-conjugase (E2), and ubiquitin ligase (E3). The E3 ligase is of particular importance because it usually comes in direct contact with protein substrates, thereby playing a crucial role in defining substrate specificity. The significance of the UPS in cancer is underscored by frequent tumor-susceptible mutations targeting the ubiquitination enzymes, and by the recent success in therapeutic use of proteasome inhibitors.
Smad ubiquitination regulatory factors 1 and 2 (Smurf1 and Smurf2), two members of the HECT family of E3 ligases, were identified based on their activities to modulate transforming growth factor (TGF)-β/BMP signaling by inducing ubiquitin modification on Smads; however, subsequent studies have expanded the repertoire of Smurf substrates, such as RhoA, Runx2, and MAPK kinase kinase 2, to both within and outside the TGF-β/BMP pathway (2). Smurf1 was further found in a RhoA-containing complex at the leading edge of cell lamelipodium, suggesting that this E3 ligase participates in controlling cell polarity and movement (3). Likewise, Smurf2 may also have a role in controlling cell movement because it can catalyze ubiquitination of Rap1B, an essential factor for the establishment of neuronal polarity (4). Moreover, elevated Smurf2 expression has been reported to correlate with poor prognosis in patients with esophageal squamous cell carcinoma (5), although it was not clear whether such correlation reflected a direct effect of Smurf2 or merely a consequence of tumor progression. Here, we report that Smurf2 expression is also elevated in certain breast cancer tissues and several cell lines and show that elevated Smurf2 expression promotes the aggressiveness of breast cancers by enhancing cell migration, invasion, and metastasis.
Materials and Methods
Cell culture
Breast cancer cell lines T47D, MDA-MB-231, HS-578T, BT-549, MCF-7, MDA-MB-435; prostate cancer cell lines PC-3 and DU145; ovarian cancer cell lines NCI/ADR-RES, OVCAR-4, OVCAR-3, OVCAR-5, OVCAR-8, IGR-OV1, A2780, and SK-OV-3 were obtained from the DCTD Tumor Repository of the National Cancer Institute. Primary human mammary epithelial cells (HMEC) were purchased from Cambrex. Immortalized normal human breast epithelial cell line MCF-10A and breast cancer cell line MDA-MB-468 were from American Type Culture Collection. Metastatic breast cancer cell line MCF10Ca1a (6) was maintained in DMEM/F-12 supplemented with 5% horse serum.
Expression plasmids and retroviral infection
Retroviral vector, pBabe-puro, was used to express FLAG-tagged Smurf2 and Smurf2C716G (7). Viral production and infection were done as described (8). Stable MCF10Ca1a cells were established after infection with empty vector, pBabe-FLAG-Smurf2, or pBabe-FLAG-Smurf2C716G retroviruses under puromycin selection (8 μg/mL).
RNAi experiments
Target sequences for siRNA used are as follows: nonsilencing (5′-AATTCTCCGAACGTGTCACGT-3′), Smurf1 (5′-AACCTTGCAAAGAAAGACTTC-3′; ref. 8), and Smurf2 (5′-GCAGTTAATCCGGAACATTTA-3′). Oligofectamine (Invitrogen) was used to deliver of siRNAs into cells (9). Briefly, for 1 well of a 12-well plate, 1.68 μg siRNA duplex (6 μL of 20 μmol/L siRNA duplex) in 100 μL of Opti-MEM were mixed with 6 μL of OLIGOFECTAMINE Reagent in 100 μL of Opti-MEM. After incubating for 20 to 25 min at room temperature, the 200 μL of siRNA-OLIGOFECTAMINE was added to cultured cells (40−50% confluent) in 400 μL of Opti-MEM. Regular medium (600 μL) containing 20% fetal bovine serum (FBS) was added to the transfected cells at 3 h after transfection. Two days posttransfection, cells were harvested for Westernblot or starved overnight before migaration or invasion assay.
Western blot, immunofluorescence,and immunohistochemistry
Breast cancer tissue array BR801 with matching normal tissues was from US BioMax, Inc. Immunohistochemistry of the array was analyzed by a board certified pathologist (Dr. Anver). Methods for Western blot, immunofluorescence, and immunohistochemistry were described previously (7, 8, 10). Antibodies used were as follows: Smurf1 (H-60), Smurf2 (H-50), Smad5, Smad8 (R64), and TβRI (V-22) (Santa Cruz Biotech); Smad1 (EP565Y), Smad2 (EP784Y), and Smad3 (EP568Y) (Epitomics); c-terminally phosphorylated Smad1/5/8 and phosphorylated Smad2 (Cell Signaling Technology); BMPRII (R&D) Smad2/3, E-cadherin, N-cadherin, and β-catenin (BD Biosciences); Texas-Red–conjugated-phalloidin; glyceraldehyde-3-phosphate dehydrogenase (Biodesign); and FLAG M2 monoclonal antibody (Sigma).
Growth inhibition assay
Cells (2−4 × 104) seeded in 24-well plates were cultured with or without TGF-β treatment for 24 h, and 5 μCi/mL [3H]-thymidine (PerkinElmer) was added during the last 3 h. Incorporation of [3H]-thymidine was measured as described (10).
Cell migration and invasion assay
Cell invasion assay was performed in Matrigel Invasion Chambers (BD Biosciences). Cells were starved overnight in serum-free medium before the experiment. MDA-MB-231 (2.5 × 104) or MCF10ACa1a (1 × 105) in 0.2 mL serum-free medium containing 0.1% bovine serum albumin (BSA) or 0.1% horse serum, respectively, were added to the upper well of transwell chamber. Conditional medium (0.6 mL) from NIH3T3 cells or complete medium containing 10% FBS were added to the bottom well for MDA-MB-231 or MCF10ACala cells, respectively. Conditional medium from NIH3T3 cells was generated by incubating NIH3T3 cells at 90% confluence for 2 d with DMEM containing 10% FBS. After incubation for 40 h at 37°C, cells remaining in the top well were removed by swiping with cotton swabs thrice to allow for specific staining of cells on the underside of the membrane using the Diff-Quik stain kit (Dade Boehringer). All cells on the membrane were counted under microscope.
Transwell migration assays were performed essentially as the invasion assays above except that a 6-mm Biocoat Cell Culture Control Inserts (BD Biosciences) was used instead of the Matrigel inserts. Migrating MDA-MB-231 or MCF10ACa1a cells were counted after incubation at 37°C for 5 or 25 h, respectively. Cells on 50% of the membrane area were counted and extrapolated to 100% of the membrane surface. All migaration and invasion experiments described in the article were performed in triplicates and repeated three to five times.
Metastasis assay
Four- to 6-week-old female athymic nu/nu mice were injected i.v. with 5 × 105 cells in 0.2 mL of DMEM/F12 through the tail vein. Five weeks after injection, mice were euthanized and examined grossly at necropsy for the presence of metastases in internal organs. Animal care was provided according to the US Public Health Policy and Guide on the Care and Use of Laboratory Animals. Quantification of metastases in the lung was performed as described (11). Statistical analyses were performed using a two-tailed Student's t test.
Results
Expression of Smurf2 is elevated in human cancer tissues and cell lines
To explore the possibility that Smurf2 plays a role in carcinogenesis, we first screened for Smurf2 expression in breast cancer tissue arrays by immunohistochemistry. Like Smurf1, immunostaining of Smurf2 was detected in normal mammary gland in both cytoplasm and nucleus of luminal epithelial cells (Fig. 1A, A4, D6, and E10). Comparing to their matching normal tissues in the surrounding, 17.5% or 30% of mammary ductal carcinomas in the array displayed elevated Smurf1 or Smurf2, respectively, and the Smurf1 and Smurf2 in the tumor cells appeared to be exclusively in the cytoplasm (Supplementary Table; Fig. 1A, A3, D5, and E9). Interestingly, only two carcinomas have elevated expression of both Smurf1 and Smurf2, others showed either elevated Smurf2 or Smurf1 expression with the other Smurf unchanged or down-regulated (Supplementary Table; Fig. 1A).
Figure 1.
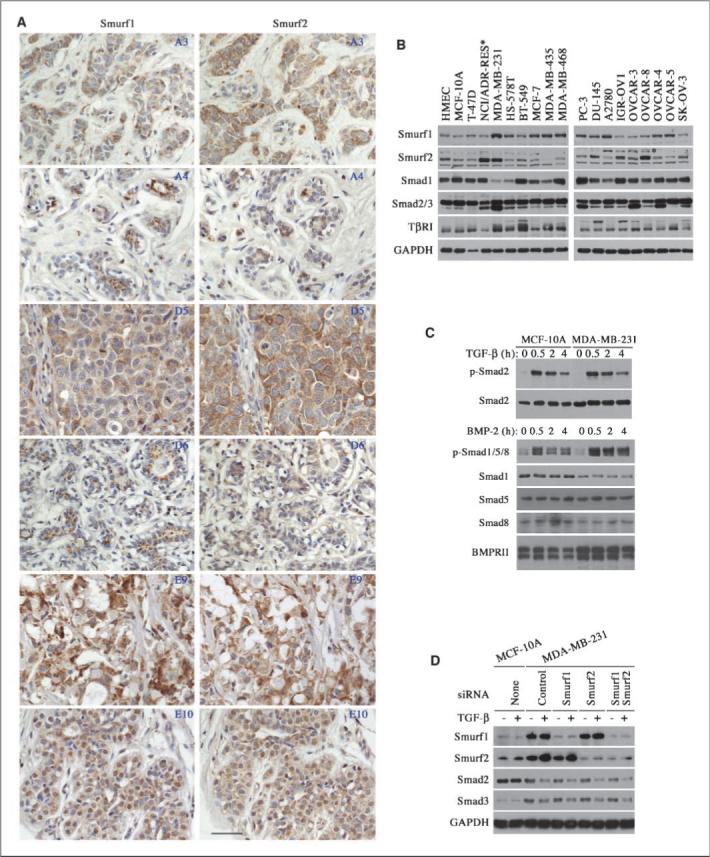
Up-regulation of Smurf2 in breast cancers. A, immunohistochemical staining of tumor (A3, D5, E9) and matching normal (A4, D6, E10) tissues in a breast cancer tissue array with anti-Smurf1 or anti-Smurf2 antibody. Three pairs of representative images are shown here and coordinates of each sample in the array are given in the top right corner. Scale bar, 4 μm. B, expression of Smurfs, Smads, and TβRI receptor in various cancer and normal control cells. *, a nonspecific band above the Smurf2 band. C, normal activation of Smads in MCF-10A and MDA-MB-231 cells. D, attenuated Smad expression induced by prolonged exposure (2 d) to TGF-β is resistant to RNAi-mediated knockdown of Smurf1 and/or Smurf2 in MDA-MB-231 cells. Left, protein levels in normal MCF-10A cells.
We next examined the expression of Smurf1, Smurf2, and several components of the TGF-β pathway in 7 breast cancer cell lines to substantiate the above finding. As controls, we included a primary and an untransformed but immortalized HMEC lines, HMEC and MCF-10A, respectively, in the study. Smurf2 expression was decreased in MCF10A cells comparing to that of HMEC cells, which is consistent with a previous observation that immortalization prevents induction of Smurf2 (12). Surprisingly, Smurf2 was strongly up-regulated in MDA-MB-231 cells (Fig. 1B). In addition, a high level of Smurf2 was also detected in NCI/ADR-RES cells (Fig. 1B), which were recently reclassified as a cell line derived from OVCAR-8 ovarian cancer cells (13). In light of this, we expanded our study to several ovarian and prostate cancer cell lines, and found a high level of Smurf2 in three ovarian lines, IGR-OV1, OVCAR-3, and OVCAR-8, and in one prostate cancer line, DU145 (Fig. 1B). Interestingly, in most of the cell lines examined in Fig. 1B, Smurf1 and Smurf2 exhibited a mutually exclusive up-regulation; that is whenever Smurf1 level was up-regulated, Smurf2 was not, and vice versa. Taken together, these results suggest that Smurf1 and Smurf2 may have redundant function in breast cancer cells.
To our surprise, the up-regulation of Smurfs in the above cancer cell lines was not followed by a concomitant change in the protein levels of Smad1, Smad2/3, or TGF-β type I receptor (Fig. 1B); all are established targets of Smurfs. Therefore, we compared the TGF-β signaling response between normal MCF-10A cells that express low level of Smurf2 and breast cancer MDA-MB-231 cells that express high Smurf2, and found that the TGF-β–induced Smad2 phosphorylation proceeded at a similar intensity and duration between these two cell lines (Fig. 1C). In addition, the BMP signaling response was not decreased in MDA-MB-231 cells, although Smad1 protein level was lower in this line of cells, possibly due to the elevated expression of BMP type II receptor (Fig. 1C). Previously, it was reported that prolonged TGF-β treatment leads to a decrease in total Smad2 and Smad3 levels in MDA-MB-231 but not in MCF-10A cells (14). This regulation does not seem to be mediated by the elevated Smurf1 and/or Smurf2, as knockdown Smurf1 and/or Smurf2 expression using siRNA did not recuperate Smad2 and Smad3 levels in MDA-MB-231 cells after exposure to TGF-β for 2 days (Fig. 1D). These results suggest that the level of Smurfs is probably not a rate-limiting factor in determining TGF-β/BMP responses in breast cancer cells.
Elevated Smurf2 is required for the enhanced migration and invasiveness of MDA-MB-231 cells
It was reported previously that reducing Smurf1 expression induced cell rounding and resulted in a loss of membrane protrusion and cell motility in highly invasive and metastatic MDA-MB-231 breast cancer cells (15). We observed a similar morphologic change in these cells after knockdown of Smurf2 expression. Staining the MDA-MB-231 cells that were transfected with Smurf-specific siRNAs for filamentous (F)-actin illuminated cortical actin instead of the typical actin-rich lamelipodial extension that could be readily detected in the control cells (Fig. 2A). These findings suggest that Smurfs may have a role in regulating tumor cell migration and invasion. To confirm this, we assessed the ability of MDA-MB-231 cells to migrate through multiporous polycarbonate membrane or invade into Matrigel-coated membrane in transwell Boyden chambers, respectively. We found that reducing either Smurf1 or Smurf2 expression via siRNA greatly diminished the ability of MDA-MB-231 cells to migrate and invade (Fig. 2B–C). These results suggest that the elevated Smurfs may be required for preparing cancer cells to migrate and invade, thereby promoting metastasis.
Figure 2.
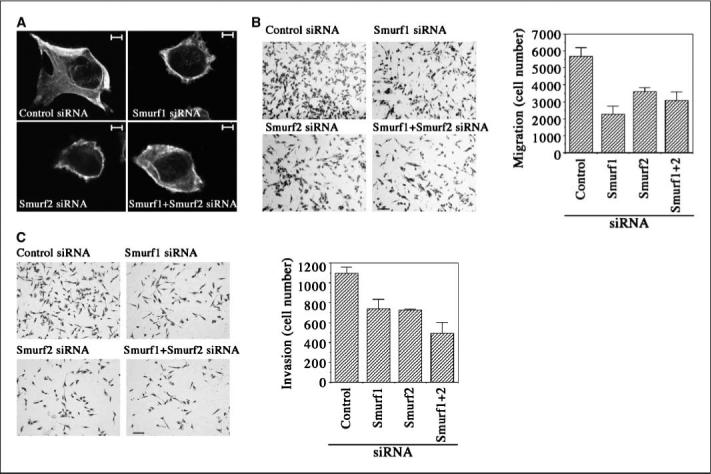
Requirement of elevated Smurf expression for the migration and invasion of MDA-MB-231 cells. Transient transfection of siRNAs was used to attenuate Smurfs expression in MDA-MD-231 cells. A, immunofluorescent staining of F-actin showing disruption of cell protrusion as the result of interfering with Smurfs expression. Scale bar, 5 μm. B and C, cell migration and invasion assays in transwell chambers. Scale bar, 10 μm.
Smurf2 promotes metastasis in vivo
To determine if indeed Smurfs play a role in the metastasis of breast cancer cells, we utilized a metastatic MCF10Ca1a cell system that was derived from a carcinoma arising from xenografts of activated HRas-transformed MCF-10A cells (6). MCF10Ca1a cells formed rapidly growing carcinomas in 7 to 10 days on xenografting and metastasized to the lung upon injection into tail vein (6, 11). Expression of endogenous Smurf1 and Smurf2 in MCF10Cala cells did not change appreciably from the low levels in the parental cells (Fig. 3A). To alter expression and activity of Smurf2, we created a stable line of MCF10Cala cells expressing NH2-terminal FLAG-tagged Smurf2 and a line expressing an E3 ligase–defective mutant of Smurf2 that carries a C716G point mutation (Fig. 3B). The metastatic potential of these stable cells was tested in nude mice. Our results indicated that 88% (22 of 25) of mice injected with Smurf2-expressing cells developed lung metastases, compared with 68% (17 of 25) injected with cells carrying the empty vector (Fig. 3C), and the numbers of histologically confirmed metastases per lung were also significantly higher in the experimental group with Smurf2-expressing (Fig. 3D). In contrast, the percentage of animals with lung metastases was decreased in the group injected with Smurf2CG-expressing cells, with only 52% (13 of 25) of animals developing lung metastases (Fig. 3C). In addition, there was a significant reduction in the number of metastases in the affected lungs of the group of animals injected with Smurf2CG-expressing cells (Fig. 3D), suggesting a dominant negative inhibition. These results indicate that overexpressing Smurf2 enhances the ability of tumor cells to metastasize, whereas inhibiting Smurf2 function has the opposite effect.
Figure 3.
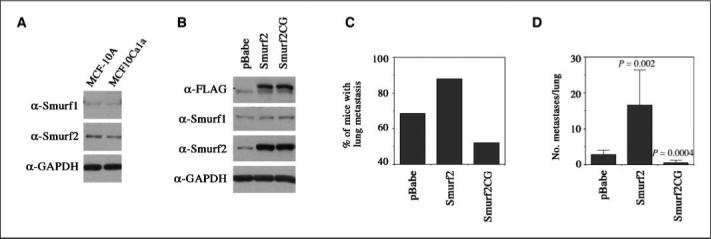
Elevated Smurf2 expression promotes breast cancer cell metastasis. A, comparable endogenous Smurf1 and Smurf2 levels between normal MCF-10A and metastatic MCF10Ca1a cells. B, elevated Smurf2 and Smurf2CG expression in the MCF10Ca1a cell–derived stable lines. C, percentage of animals with lung metastases (n = 25), and (D) number of histologically confirmed lung metastasis foci per lung in mice inoculated with stable MCF10Ca1a cells bearing pBabe vector, pBabe-Smurf2, or pBabe-Smurf2CG (n = 25). Columns, mean; bars, SD. Statistically significant differences (P) between Smurf2 or Smurf2CG-expressing cells versus pBabe-puro vector control cells are indicated.
Smurf2 promotes migration and invasiveness of MCF10Cala cells
Epithelial-to-mesenchymal transition (EMT) is a transient change in cell structure often associated with weaker cell-cell interactions and acquisition of motile and invasive properties of the cells (16). The ability of tumor cells to metastasize in vivo is correlated with EMT, and increased migration and invasiveness, in which TGF-β has an enhancing role (16, 17). MCF10Cala cells are highly malignant metastatic cancer cells, and TGF-β treatment did not cause any changes in E-cadherin or β-catenin expression in these cells (Fig. 4A). However, the level of N-cadherin was significantly increased in response to TGF-β treatment (Fig. 4A), consistent with a role of this nonepithelial cadherin in promoting breast cancer cell migration and invasion (18). Overexpressing Smurf2 increased the level of N-cadherin in MCF10Cala cells even without TGF-β treatment, whereas overexpressing the C716G mutant had an opposite effect (Fig. 4A). These results suggest that Smurf2 may promote EMT by up-regulating N-cadherin expression. Interestingly, these stable cells maintained a normal phospho-Smad2 response to TGF-β stimulation (Fig. 4A), and altering Smurf2 expression did not affect TGF-β–mediated growth inhibition response as determined by thymidine incorporation assay (Fig. 4B).
Figure 4.
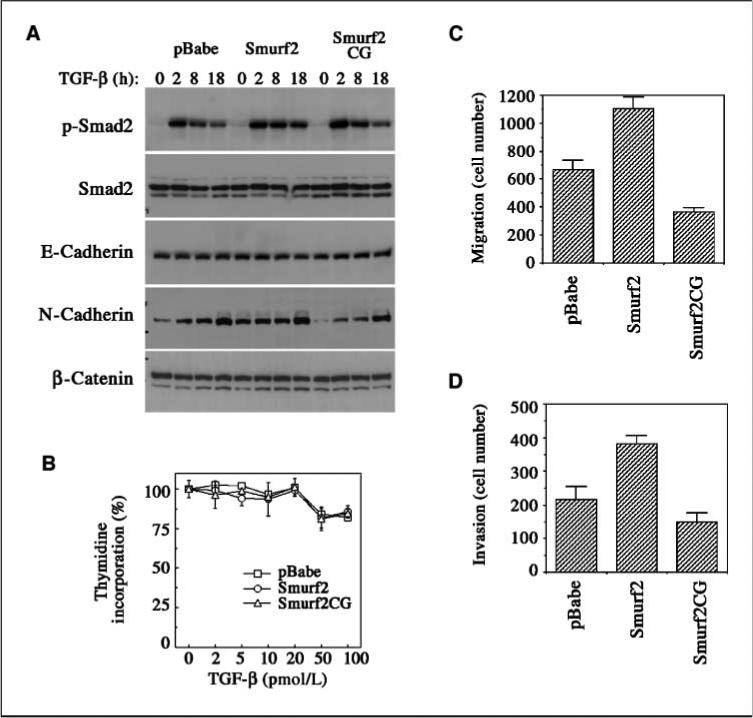
Enhanced N-cadherin expression, migration, and invasiveness of MCF10Ca1a cells expressing elevated Smurf2. A, Western analyses of stable MCF10Cala cells for Smads and cadherin expression. B, thymidine incorporation assay for TGF-β–induced growth inhibition. C to D, transwell migration and invasion assays of stable MCF10Ca1a cells.
We then examined the effects of Smurf2 and Smurf2CG on the migration and invasiveness of MCF10Cala cells in vitro. We found that significantly more MCF10Cala cells expressing Smurf2 migrated through the transwell membrane and invaded through Matrigel than control cells expressing empty vector, whereas much fewer cells expressing Smurf2CG migrated and invaded through (Fig. 4C–D). These results correlate with the results of in vivo metastasis experiments, indicating that Smurf2 has an ability to promote breast cancer metastasis through increasing cell motility and invasiveness.
Discussion
Smurf2 was originally identified as the Smad ubiquitin ligase that induces the ubiquitination and degradation of Smad1 and Smad2 (7, 19). Smurf2 can also target TGF-β type I receptor for degradation (20). However, the effect of Smurf2 in the TGF-β/Smad pathway seems to be lost in advanced metastatic breast cancer cells. It was also reported that Smurf1 does not interfere with Smad signaling in MDA-MB-231 cells (15). Therefore, the regulation of Smurf1 or Smurf2 of breast cancer cell migration or invasion may be independent of TGF-β/Smad signaling. Interestingly, during neuronal differentiation, Smurf1 and Smurf2 coordinately regulate neurite extension and neuronal polarity through targeting RhoA and Rap1B, respectively (4). RhoA was also identified as a target of Smurf1 in breast cancer cells (15). However, we did not observe significant down-regulation of RhoA or Rap1B activity in Smurf2-overexpressing MCF10Ca1a cells (data not shown). Studies are under way to define the target proteins regulated by Smurf2 in breast cancer cells. The identification of these targets should provide new insights into the function of Smurf2 in breast cancer progression and metastasis.
Supplementary Material
Acknowledgments
Grant support: Intramural Research Program of the Center for Cancer Research, National Cancer Institute, and in part by funds from the National Cancer Institute under contract N01-CO-12400.
We thank Dr. F. Miller for providing MCF10Ca1a cells, and Drs. B. Tang and L. Wakefield for advice in culturing these cells.
Footnotes
Note: Supplementary data for this article are available at Cancer Research Online (http://cancerres.aacrjournals.org/).
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
References
- 1.Hershko A, Ciechanover A. Annu Rev Biochem. 1998;67:425–79. doi: 10.1146/annurev.biochem.67.1.425. [DOI] [PubMed] [Google Scholar]
- 2.Zhang YE, Yamashita M. Smurfs in TGF-β signaling and regulation of bone homeostasis and cancer. In: Jakowlew SB, editor. Transforming growth factor-β in cancer therapy. Vol. 1. Humana Press; 2008. pp. 155–68. [Google Scholar]
- 3.Wang HR, Zhang Y, Ozdamar B, et al. Regulation of cell polarity and protrusion formation by targeting RhoA for degradation. Science. 2003;302:1775–9. doi: 10.1126/science.1090772. [DOI] [PubMed] [Google Scholar]
- 4.Schwamborn JC, Muller M, Becker AHM, Puschel AW. Ubiquitination of the GTPase Rap1B by the ubiquitin ligase Smurf2 is required for the establishment of neuronal polarity. EMBO J. 2007;26:1410–22. doi: 10.1038/sj.emboj.7601580. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 5.Fukuchi M, Fukai Y, Masuda N, et al. High-level expression of the Smad ubquitin ligase Smurf2 correlates with poor prognosis in patients with esophageal squamous cell carcinoma. Cancer Res. 2002;62:7162–5. [PubMed] [Google Scholar]
- 6.Santner SJ, Dawson PJ, Tait L, et al. Malignant MCF10CA1 cell lines derived from premalignant human breast epithelial MCF10AT cells. Breast Cancer Res Treat. 2001;65:101–10. doi: 10.1023/a:1006461422273. [DOI] [PubMed] [Google Scholar]
- 7.Zhang Y, Chang C, Gehling DJ, Hemmati-Brivanlou A, Derynck R. Regulation of Smad degradation and activity by Smurf2, an E3 ubiquitin ligase. Proc Natl Acad Sci U S A. 2001;98:974–9. doi: 10.1073/pnas.98.3.974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ying SX, Hussain ZJ, Zhang YE. Smurf1 facilitates myogenic differentiation and antagonizes the bone morphogenetic protein-2-induced osteoblast conversion by targeting Smad5 for degradation. J Biol Chem. 2003;278:39029–36. doi: 10.1074/jbc.M301193200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Elbashir SM, Harborth J, Lendeckel W, Yalcin A, Weber K, Tuschl T. Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells. Nature. 2001;411:494–8. doi: 10.1038/35078107. [DOI] [PubMed] [Google Scholar]
- 10.Yang YA, Zhang GM, Feigenbaum L, Zhang YE. Smad3 reduces susceptibility to hepatocarcinoma by sensitizing hepatocytes to apoptosis through down-regulation of Bcl-2. Cancer Cell. 2006;9:445–57. doi: 10.1016/j.ccr.2006.04.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Tang B, Vu M, Booker T, et al. TGF-β switches from tumor suppressor to prometastatic factor in a model of breast cancer progression. J Clin Invest. 2003;112:1116–24. doi: 10.1172/JCI18899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Zhang H, Cohen SN. Smurf2 up-regulation activates telomere-dependent senescence. Genes Dev. 2004;18:3028–40. doi: 10.1101/gad.1253004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Liscovitch M, Ravid D. A case study in misidentification of cancer cell lines: MCF-7/Adr cells (redesignated NCI/ADR-RES) are derived from OVCAR-8 human ovarian carcinoma cells. Cancer Lett. 2007;245:350–2. doi: 10.1016/j.canlet.2006.01.013. [DOI] [PubMed] [Google Scholar]
- 14.Brown KA, Aakre ME, Gorska AE, et al. Induction by transforming growth factor-β1 of epithelial to mesenchymal transition is a rare event in vitro. Breast Cancer Res. 2004;6:R215–31. doi: 10.1186/bcr778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Sahai R, Garcia-Medina R, Pouyssegur J, Vial E. Smurf1 regulates tumor cell plasticity and motility through degradation of RhoA leading to localized inhibition of contractility. J Cell Biol. 2007;176:35–42. doi: 10.1083/jcb.200605135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Thiery JP. Epithelial-mesenchymal transitions in tumour progression. Nat Rev Cancer. 2002;2:442–54. doi: 10.1038/nrc822. [DOI] [PubMed] [Google Scholar]
- 17.Oft M, Peli J, Rudaz C, et al. TGF-β1 and Ha-Ras collaborate in modulating the phenotypic plasticity and invasiveness of epithelial tumor cells. Genes Dev. 1996;10:2462–77. doi: 10.1101/gad.10.19.2462. [DOI] [PubMed] [Google Scholar]
- 18.Nieman MT, Prudoff RA, Johnson KR, et al. N-cadherin promotes motility in human breast cancer cells regardless of their E-cadherin expression. J Cell Biol. 1999;147:631–44. doi: 10.1083/jcb.147.3.631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Lin X, Liang M, Feng XH. Smurf2 is a ubiquitin E3 ligase mediating proteasome-dependent degradation of Smad2 in transforming growth factor-β signaling. J Biol Chem. 2000;275:36818–22. doi: 10.1074/jbc.C000580200. [DOI] [PubMed] [Google Scholar]
- 20.Kavsak P, Rasmussen RK, Causing CG, et al. Smad7 binds to Smurf2 to form an E3 ubiquitin ligase that targets the TGF β receptor for degradation. Mol Cell. 2000;6:1365–75. doi: 10.1016/s1097-2765(00)00134-9. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.