

Abstract
Symbiotic bacteria of the genus Rhizobium synthesize lipo-chitooligosaccharides, called Nod factors (NFs), which act as morphogenic signal molecules on legume hosts. The common nodABC genes, present in all Rhizobium species, are required for the synthesis of the core structure of NFs. NodC is an N-acetylglucosaminyltransferase, and NodB is a chitooligosaccharide deacetylase; NodA is involved in N-acylation of the aminosugar backbone. Specific nod genes are involved in diverse NF substitutions that confer plant specificity. We transferred to R. tropici, a broad host-range tropical symbiont, the ability to nodulate alfalfa, by introducing nod genes of R. meliloti. In addition to the specific nodL and nodFE genes, the common nodABC genes of R. meliloti were required for infection and nodulation of alfalfa. Purified NFs of the R. tropici hybrid strain, which contained chitin tetramers and were partly N-acylated with unsaturated C16 fatty acids, were able to elicit nodule formation on alfalfa. Inactivation of the R. meliloti nodABC genes suppressed the ability of the NFs to nodulate alfalfa. Studies of NFs from nodA, nodB, nodC, and nodI mutants indicate that (i) NodA of R. meliloti, in contrast to NodA of R. tropici, is able to transfer unsaturated C16 fatty acids onto the chitin backbone and (ii) NodC of R. meliloti specifies the synthesis of chitin tetramers. These results show that allelic variation of the common nodABC genes is a genetic mechanism that plays an important role in signaling variation and in the control of host range.
Keywords: Rhizobium–legume symbiosis/nod genes/nodulation factors/plant–microbe interactions/oligosaccharides
The Rhizobium–legume symbiosis is an efficient system for dissecting the molecular basis of host specificity, because of the high specificity of some of these associations and the well-studied genetics of a number of the bacterial partners (Rhizobium is defined here as all bacteria that nodulate root or stem of legumes). Rhizobium meliloti strains, for example, elicit the formation of nitrogen-fixing nodules on only three genera, Medicago, Melilotus, and Trigonella. Nodulation (nod, nol, and noe) genes that control host specificity, infection, and nodulation specify the synthesis of Nod factors (NFs) that act as specific growth regulators on legume hosts (1–4). NFs from all Rhizobium species belong to the same chemical family; they are chitin tetramers or pentamers, mono-N-acylated at the nonreducing end. The chitin oligomer backbone, however, can be diversely substituted at both ends of the molecule, and the substitutions are generally characteristic of a given rhizobial species having a defined host range.
A number of genetic and biochemical mechanisms by which regulatory and structural nodulation genes control host specificity have been identified. The regulatory nodD genes encode DNA-binding proteins that activate the transcription of the other nod operons (1). These genes determine a first level of host specificity, since the NodD proteins are activated by plant signals whose nature and abundance in root exudates may vary according to the plant host (5, 6). Structural nod genes comprise both genes that are species-specific and genes that are common to all Rhizobium species. Specific nod genes are major determinants of host specificity, each Rhizobium species being characterized by a given combination of specific nod genes (3, 4, 7). These genes encode enzymes that are involved in the “decoration” of the NF chitooligosaccharide backbone with substitutions that confer plant specificity (8–14).
The common nodABC genes encode enzymes involved in the synthesis of the lipo-oligosaccharide core. NodC is an N-acetylglucosaminyltransferase that determines the synthesis of the chitin oligomer backbone (15, 16), NodB is a chitooligosaccharide deacetylase (17), and NodA is required for the N-acylation of the aminosugar backbone (18, 19). nodABC genes have been thought to be functionally equivalent between Rhizobium species (20–22). In this paper, we demonstrate, by a gain-of-function approach, that the R. meliloti nodABC genes are host-range determinants. We have shown that the R. meliloti nodABC genes are required to allow Rhizobium tropici to infect and nodulate alfalfa and to produce new NFs that elicit nodule formation on alfalfa. These new NFs had characteristics of R. meliloti factors, including a tetrameric chitooligosaccharide backbone and N-acylation with polyunsaturated C16 fatty acids.
MATERIALS AND METHODS
Strain Construction.
Bacteria, strains, and plasmids used in this study are described in Table 1 and Fig. 1. R. meliloti and R. tropici strains were grown on tryptone yeast and Bergersen agar media, respectively (29). Plasmids were introduced into R. tropici by triparental mating as described (14, 30). To determine the precise location of Tn5 insertions in the R. meliloti nodABCIJ genes, a junction fragment was amplified by PCR with Taq DNA polymerase (Life Technologies SARL, Cergy-Pontoise, France). The oligonucleotide primers were 5′-CGATGAAGAGCAGAAGT-3′ (Insertion sequence 50 specific) and 5′-GAAGAAACAACACAC-3′ (nodA specific). The amplified fragments were cloned into pUC18 and sequenced with the oligonucleotide primer internal to insertion sequence 50 (31).
Table 1.
Bacteria and plasmids used in this study
| Designation | Relevant characteristics | Ref./source |
|---|---|---|
| R. meliloti | ||
| GMI51 | RCR2011 = SU47, wild type, Nod+, Fix+ on M. sativa | 23 |
| R. tropici | ||
| GMI9776 | CFN299, wild type, NalR, Brazil | 24 |
| GMI3067 | CFN299 (pGMI149)(pGMI1962) | This study |
| GMI3066 | CFN299 (pGMI1962) | This study |
| Plasmids | ||
| pGMI149 | pRK290 prime (IncP), TcR | 25 |
| pGMI1962 | = pnodL, pMP190-prime (IncQ), CmR, | 12 |
| pGMI353 | pGMI149 nodA::Tn5#2208 | 25 |
| pGMI318 | pGMI149 nodB::Tn5#154 | 25 |
| pGMI362 | pGMI149 nodC::Tn5#2217 | 25 |
| pGMI377 | pGMI149 nodI::Tn5#2111 | 25 |
| pGMI1984 | pGMI149 nodJ::Tn5#2412 | 25 |
| pGMI41589 | pGMI149 nodP::Tn5 | 9 |
| pGMI306 | pGMI149 nodQ::Tn5#115 | 25 |
| pGMI336 | pGMI149 nodH::Tn5#2313 | 25 |
| pGMI5625 | pGMI149 Δ(nodF)6 | 13 |
| pGMI334 | pGMI149 nodE::Tn5#2309 | 25 |
| pGMI930 | pGMI149 nodD1::Tn5 | 26 |
| pGMI860 | pGMI149 nodD3::MudII lac#V40 | 26 |
| pMH682 | pWB5a-prime (IncP), 8-kb insert carrying R. meliloti nodD3 and syrM, TcR | 14 |
| pXLGD4 | pGD499-prime (IncP) carrying a hemA–lacZ fusion, TcR | 27 |
| pRK2013 | Helper plasmid for mobilization of IncP and IncQ plasmids, KmR | 28 |
Figure 1.
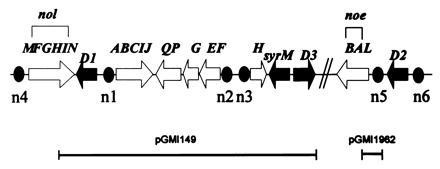
Genetic map of the R. meliloti nodulation region. The open arrows represent the structural nod, nol, and noe genes and operons, and black arrows represent the regulatory nodD and syrM genes. Filled ovals identify the nod boxes. Below the map, the two segments represent the pGMI149 and pGMI1962 plasmids.
NF Production and Purification.
Production and extraction of NFs were carried out as described by Poupot et al. (32). The nod gene inducer was luteolin (10 μM), and the NF yield was approximately 2 mg/liter. NFs were then purified by reverse phase HPLC as described (13). Further separation of tetrameric and pentameric NFs was achieved by analytical reverse phase HPLC chromatography on a C18 column (4.6 × 250 nm, ODS2, 5 μm, ColoChrom, Spherisorb) in the presence of 50 mM ammonium acetate as indicated below. Compounds were eluted from the column at a flow rate of 1 ml/min under the following conditions: solvent A, 7:3 H2O/CH3CN; solvent B, 3:7 H20/CH3CN; both solvent A and solvent B were supplemented with 50 mM ammonium acetate. After 15 min in isocratic conditions with solvent A, NFs were eluted with a linear gradient to solvent B in 30 min.
Structural Analysis of NFs.
Fatty acids were released from NFs by acid methanolysis and analyzed by GC as described (13). Fatty acids were characterized by coinjection with authentic standards and by GC/MS analysis, for which a Hewlett Packard gas chromatogram 5890A/mass spectrometer 5970A/Data System with electron impact MS was used. Methyl ester fatty acids were chromatographed on a DB1 column (30 m). Column temperature was first programmed from 80 to 180°C at 20°C/min, then from 180 to 260°C at 6°C/min, and finally from 260°C to 300°C at 20°C/min.
Matrix-assisted laser desorption ionization-time of flight (MALDI) mass spectra were recorded on a TOFSPEC instrument (Fisons, VG Analytical, Manchester, U.K.) equipped with an N2 laser (337 nm, 18 kV extraction voltage) and a reflectron. These experiments were conducted with 2,5-dihydroxybenzoic acid as the matrix. The targets, which consisted of flat, stainless-steel plates, were loaded with 1 μl of the matrix solution [7.5 mg of matrix in 1 ml of water/acetonitrile (2:1 vol/vol) with 0.1% trifluoroacetic acid] and 1 μl of the aqueous sample solution (15–75 ng/μl sample from HPLC-purified NFs) and allowed to dry. Measurements were performed with negative and positive detection modes. Liquid secondary ion mass spectrometry mass spectra of NFs were obtained as described (14).
Plant Assays.
Plant assays were performed as described (14) with the following modifications. Seeds of Medicago sativa cv. Europa, provided by Florimond Desprez (Cappelle, 59242 Templeuve, France) were used in this study. NFs were introduced into melted Fahraeus agar medium (29) before solidification. Infection was examined on plants inoculated with bacterial strains carrying a fusion of the reporter gene lacZ of Escherichia coli to the nod box 5 region of R. meliloti (12) as described (14).
RESULTS
Choice of the Recipient Strain for Gain-of-Function Experiments.
To address the question of a possible allelic variation of nodABC genes among rhizobia, we hypothesized that these genes could encode enzymes having similar catalytic activities in different species, with NodC controlling the length of the chitin oligomer backbone (four or five glucosamine residues) and NodA mediating N-acylation in a specific way (polyunsaturated C16 fatty acids or vaccenic acid). Genetic analysis of this problem required the transfer of R. meliloti nod genes into a Rhizobium species having appropriate characteristics. We have chosen R. tropici CFN299 as the recipient strain for the following reasons. (i) In spite of producing sulfated NFs, it does not nodulate alfalfa. (ii) NodRt factors are different from NodRm factors for the two relevant characteristics studied, because they are glucosamine pentamers and are N-acylated by vaccenic acid (32) (Fig. 2). (iii) The NF pattern is very simple (only two major compounds), which should facilitate studies of structural changes. (iv) NodRt factors are similar to NodRm factors in that they are sulfated and not glycosylated at C-6 of the reducing end and are not carbamoylated at the nonreducing end. (v) This strain produces NFs abundantly. (vi) R. meliloti nod genes are strongly expressed (33).
Figure 2.
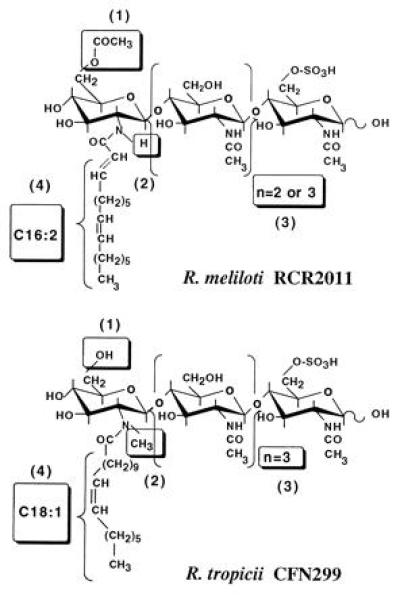
Structures of the major NFs of R. meliloti and R. tropici. Differences between the NFs from the two species are boxed. (1) ± O-acetate group; (2) ± N-methyl group; (3) number of N-acetylglucosamine residues; (4) structure of the acyl chain.
R. tropici Containing R. meliloti nod Genes Can Infect and Nodulate Alfalfa.
We first introduced into R. tropici an IncP plasmid, pGMI149, carrying the main R. meliloti nodulation region (Fig. 1). The R. tropici (pGMI149) transconjugants nodulated M. sativa very poorly (Fig. 3A). When a second plasmid, of the IncQ group (compatible with pGMI149), carrying the nodL gene (pGMI1962) was introduced into R. tropici (pGMI149), a better nodulation was observed (Fig. 3A). Root infection was then studied cytologically. The plasmid pGMI1962 carries the lacZ gene of E. coli downstream of nodL and the “associated” nod box n5, allowing the use of a β-galactosidase assay to facilitate the observation of bacteria in planta. Alfalfa seedlings were inoculated with the R. tropici (pGMI149)(pGMI1962) hybrid strain and examined 7 days after inoculation; bacteria could form infection threads (ITs; data not shown). Three weeks after inoculation, nodules were found to have a zone II containing a network of ITs, and in some nodules, bacteria were released into the cytoplasm of host cells (data not shown). However, no nitrogenase activity could be detected.
Figure 3.
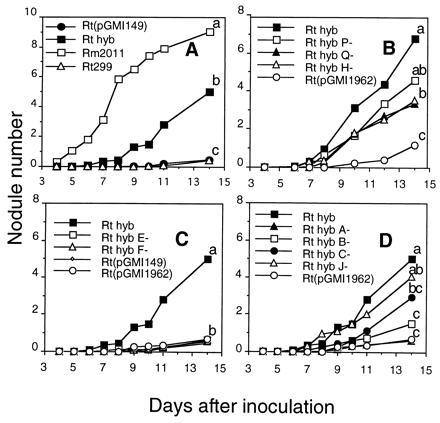
Induction of nodule formation on M. sativa cv. Europa by living bacterial cells of R. tropici CFN299 carrying various combinations of R. meliloti nod genes. (A) R. meliloti, Rm2011; R. tropici, Rt299; R. tropici (pGMI149), Rt (pGMI149); R. tropici (pGMI149)(pGMI1962), Rt hyb. (B–D) R. tropici (pGMI149)(pGMI1962) with Tn5 insertions in the R. meliloti nod genes carried in pGMI149. (B) Mutations in the sulfation genes nodP, nodQ, and nodH. (C) Mutations in the nodFE and nodL genes. (D) Mutations in the nodA, nodB, nodC, and nodIJ genes. Rt hyb, R. tropici (pGMI149)(pGMI1962); Rt hyb D1-, R. tropici (pGMI149nodD1)(pGMI1962) and so on. Nodule numbers represent the average of 20 tubes, each containing two seedlings. Statistical analysis of nodule numbers 2 weeks after inoculation: treatments with letters in common do not differ significantly at the P = 0.05 level. Analysis of variance with Fisher’s test.
The R. tropici Hybrid Strain Produces a New Class of NFs.
We prepared NFs from R. tropici, R. meliloti (with pMH682 for increasing NF production), and the R. tropici (pGMI149)(pGMI1962) hybrid strain. We then tested the ability of these NFs to form nodules on alfalfa (Fig. 4). The mixture of NFs from wild-type R. tropici did not induce nodule formation at concentrations of 10−9 M and 10−8 M. In contrast, NFs produced by the R. tropici hybrid strain were able to induce nodule formation in the same concentration range as those produced by R. meliloti (10−8 M to 10−9 M). The presence of R. meliloti nodulation genes therefore enables R. tropici to produce new NFs that can induce nodule formation on M. sativa.
Figure 4.
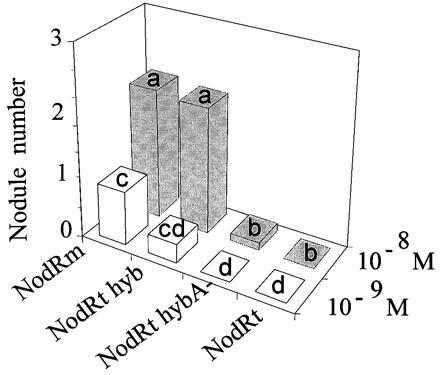
Nodules induced on M. sativa by addition of purified NFs at 10−8 M and 10−9 M. NFs were purified from the following strains. NodRm, R. meliloti RCR2011(pMH682); NodRt, R. tropici wild type; NodRt hyb, R. tropici (pGMI149)(pGMI1962); NodRt hybA−, R. tropici (pGMI149nodA)(pGMI1962). Nodules were scored 25 days after addition of the NFs. Ten tubes were used for each dilution, with two seedlings per tube. For details on the statistical analysis of nodule numbers, see the Fig. 3 legend.
NFs from the different strains were characterized chemically. Two major compounds have been described in the R. tropici wild-type strain, NodRt-V(Me, C18:1) and NodRt-V(Me, C18:1,S), having molecular weights of 1269 and 1349, respectively (33). In addition to these major NFs acylated with vaccenic acid, less abundant NFs acylated with C16:0 or C18:0 were also found (Fig. 5a). The HPLC chromatographic profile of the purified extracts from the hybrid R. tropici (pGMI149)(pGMI1962) strain was more complex than that of R. tropici (data not shown). NFs from the hybrid strain, collected as a single HPLC fraction, were first analyzed by positive liquid secondary ion mass spectrometry for structure control. Several (M + H)+ ions were obtained, all of them accompanied by ions 80 mass units lower, due to the loss of a SO3 group. The characteristic fragmentation sequence (203 mass units intervals) indicated a chitin backbone. This series ended down at oxenium ions corresponding to the last glucosamine residue at the nonreducing end. This glucosamine residue was partially acetylated or methylated, or carried both substitutions, and was acylated by a variety of fatty acids.
Figure 5.
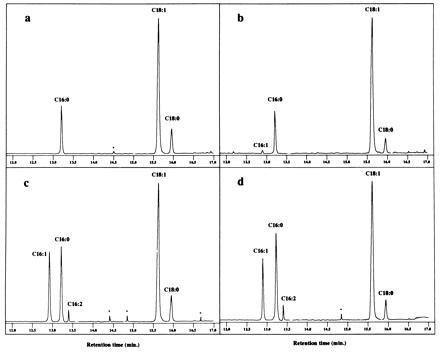
Gas chromatographic analysis of NF acyl chains. Fatty acids were released after methanolysis of NFs from the following R. tropici strains. (a) CFN299 wild type. (b) R. tropici (pGMI149nodA)(pGMI1962). (c) R. tropici (pGMI149)(pGMI1962). (d) R. tropici (pGMI149nodB)(pGMI1962).
To better estimate the relative proportion of each species, negative ion MALDI MS was performed. The absence of fragmentations and the lesser influence of hydrophobic effects on ionization would allow a better determination of the ratio between tetrameric and pentameric species. Two groups of molecules could be detected (Fig. 6A). In the group corresponding to the largest molecules, the major molecular ions corresponded to the following pentameric NFs: Nod-V(Ac, S,C18:1), Nod-V(Ac, S,C16:0), Nod-V(Ac, S,C16:1), Nod-V(S, C18:1), Nod-V(S, C16:1), and Nod-V(Ac, Me, S,C18:1). The second group represented the corresponding tetrameric NFs (Fig. 6A). MALDI spectra showed ratios of tetramer/pentamer ion abundances of about 30:50. Thus the presence of R. meliloti nod genes results in the synthesis of tetrameric NFs. GC analysis of the fatty acids released from the hybrid strain NFs after methanolysis showed that approximately 22% and 4%, respectively, were C16:1 and C16:2. These could not be detected in the R. tropici control (Fig. 5, compare a and c). The introduction of R. meliloti nod genes thus results in the synthesis of NFs N-acylated by unsaturated C16 fatty acids.
Figure 6.
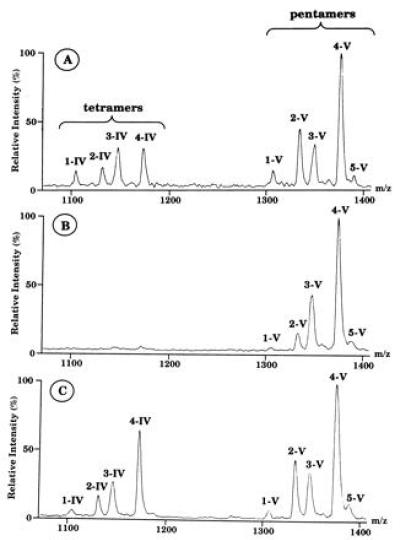
Negative ion MALDI mass spectra of NFs from the following R. tropici (pGMI149)(pGMI1962) derivatives. (A) The control strain with unmodified pGMI149. (B) A derivative with a nodC::Tn5 insertion in pGMI149. (C) A derivative with a nodI::Tn5 insertion in pGMI149. Tetrameric (IV) and pentameric (V) NFs were found with the following substitutions (nomenclature as proposed in ref. 9). 1-IV: m/z 1105 (IV, C16:0,S), m/z 1103 (IV, C16:1,S); 2-IV: m/z 1131 (IV, C18:1,S); 3-IV: m/z 1147 (IV, Ac, C16:0,S), m/z 1145 (IV, Ac, C16:1,S), m/z 1145 (IV, Me, C18:1,S); 4-IV: m/z 1173 (IV, Ac, C18:1,S); 1-V: m/z 1308 (V, C16:0,S), m/z 1306 (V, C16:1,S); 2-V: m/z 1334 (V, C18:1,S); 3-V: m/z 1350 (V, Ac, C16:0,S), m/z 1348 (V, Ac, C16:1,S), m/z 1348 (IV, Me, C18:1,S); 4-V: m/z 1376 (V, Ac, C18:1,S); 5-V: m/z 1390 (V, Me, Ac, C18:1,S).
R. meliloti Genes Required for the Infection and Nodulation of Alfalfa.
pGMI1962 is required for nodulation of alfalfa, showing the role of nodL (Fig. 3). To identify genes on pGMI149 involved in transferring the ability to nodulate alfalfa, we constructed different derivatives of CFN299 (pGMI149)(pGMI1962), each carrying a Tn5 insertion in one of the R. meliloti nod genes carried on pGMI149, and tested the ability of these different mutants to nodulate alfalfa. Mutations in the regulatory genes nodD1 and nodD3 did not result in a detectable decrease in nodulation (data not shown); thus these genes are either not involved in the extension to alfalfa of the host range, or the presence of one of them is sufficient. Mutations in the nodPQ and nodH sulfation genes caused only a 35–50% decrease in nodulation (Fig. 3B), in agreement with the previous finding that the sulfation machinery of R. tropici is active in the hybrid strain (33).
In contrast, mutations in nodFE provoked a very strong (more than 80%) decrease in nodulation, similar to the loss of the nodL gene or of the whole pGMI149 plasmid (Fig. 3C), suggesting that N-acylation with unsaturated C16 fatty acids is required to allow R. tropici to nodulate alfalfa. Interestingly, mutations in the common nodABCIJ operon also affected nodulation. The effect followed the polarity of the operon, suggesting that each of the common nodABC genes contributed to alfalfa nodulation, with a mutation in nodA having the greatest effect and a mutation in nodJ having no significant effect (Fig. 3D). Indeed, a Tn5 insertion in nodA, which probably also resulted in the inactivation of the downstream genes, had a drastic effect, similar to the inactivation of nodFE or nodL. Therefore, the “common” nod genes of R. meliloti are required for the nodulation of alfalfa. That the effect of Tn5 mutations was not due to a decrease in the amount of NF produced by the various mutants was indicated by the fact that Tn5 insertions in the various genes of the R. meliloti nodABCIJ operon did not result in any detectable decrease of the global NF production, which remained at approximately 2 mg per liter of culture. That the R. meliloti nodABC genes control the alfalfa nodulating ability of the hybrid strain by mediating qualitative changes in NFs was then shown directly. Purified NFs from the R. tropici hybrid carrying a Tn5 insertion in the R. meliloti nodA gene had lost the ability to elicit nodule formation on alfalfa roots at 10−8 and 10−9 M (Fig. 4).
To determine whether these mutations altered early stages of infection, ITs in root hairs and the root cortex were counted at 7 days, and data were statistically analyzed. Mutations in nodA, nodB, nodC, and nodE provoked a very strong decrease in IT formation (significant at P = 0.01). In contrast, mutations in nodI and nodJ did not result in any detectable reduction of IT numbers. These results indicate that in the nodABCIJ operon, the nodABC genes are clearly more important than the nodIJ genes for early infection and that the effect of the nodC mutation is not due to a polar effect on nodIJ.
Influence of the R. meliloti Common nod Genes on NF Structure.
The role of the R. meliloti nodABC genes on the structure of NFs produced by the R. tropici (pGMI149)(pGMI1962) hybrid strain was then investigated. NFs from derivatives carrying mutated nodA, nodB, nodC, nodI, or nodJ genes in pGMI149 were purified and studied by MS. MALDI MS showed that mutations in any of the three nodABC genes result in the disappearance of tetrameric NFs (Fig. 6B). In contrast, inactivation of nodI and nodJ genes did not modify the proportion of pentameric and tetrameric NFs (Fig. 6C). These results suggest that the inactivation of the R. meliloti nodC gene is sufficient to suppress the synthesis of tetrameric NFs. Fatty acids were then analyzed by GC. Whereas a mutation in nodA strikingly decreased the proportion of unsaturated C16 fatty acids (no C16:2 detectable and C16:1 reduced to 2%), a mutation in nodB did not significantly modify the proportion of these fatty acids (16% of C16:1 and 4% of C16:2) (Fig. 5 b and d). These results suggest that the inactivation of R. meliloti nodA is sufficient to alter the synthesis of NFs N-acylated with C16 unsaturated fatty acids.
DISCUSSION
The introduction of R. meliloti nodulation genes into R. tropici enables this bean symbiont to form ITs and nodules on alfalfa. Similarly, it had already been shown that the introduction of R. meliloti nod genes into R. leguminosarum bv. viciae and bv. trifolii allows these bacteria to infect and nodulate alfalfa (34, 35). This provides evidence for a major role of nod genes and NFs in the infection process and namely in IT formation. NF structural requirements are very stringent for bacterial entry and IT initiation. The N-acylation of NodRm factors with the cognate polyunsaturated C16 fatty acids and the presence of an O-acetyl group have already been shown to be required for IT initiation in alfalfa (14). These stringent NF structural requirements are confirmed in the gain-of-function experiments described in this study.
Various genetic and biochemical mechanisms by which nodulation genes control host-specificity have already been identified (36). Regulatory nodD genes encode transcriptional activators that are activated by specific plant signals (5, 6). Specific structural nod genes control the substitutions modifying the chitooligosaccharide backbone to make NFs plant-specific. Each species possesses a given combination of genes that encode enzymes involved either in the synthesis of activated substituents or for transferases that transfer the activated groups onto specific atoms of the NF backbone. Genetic variation is basically nonallelic with combinations of genes that are present in some species and absent in others; however, some species-specific nod genes such as nodFE add the possibility of allelic variation by encoding a family of enzymes that have different characteristics in different Rhizobium species.
In this paper, we have identified a novel mechanism of control of host specificity: allelic variation of the common nodABC genes. These genes, which are present in all Rhizobium species, were initially called “common,” because they were believed to be functionally equivalent in various species (20–22). This was true to some extent. nodABC-negative mutants are unable to produce NFs and thus cannot elicit plant responses and nodule formation. The introduction of an active heterologous nodABC clone restores the possibility to produce NFs and thus the ability to induce some infection and nodulation, suggesting a functional complementation. A close look at the symbiotic behavior of transconjugants, however, reveals that the complementation was often partial (37) and that if nodABC genes are common in the sense that they are present in all rhizobia, they can also be important determinants of host specificity. In this paper, we have clearly shown that the R. meliloti nodABC genes are required for, and are as important as the species-specific nodL and nodFE genes for effective infection and nodulation of alfalfa. Our results suggest that nodABC genes have evolved in different rhizobial species to contribute to the adaptation to specific legume hosts, together with species-specific nod genes.
Note Added in Proof.
Further studies are required to analyze in detail the mechanisms by which the NodABC proteins control the length of the chitooligosaccharide backbone and the specificity of the acyl transfer. In a followup of the present study, we have recently shown that the R. meliloti NodA protein is involved in the specific transfer not only of the polyunsaturated C16 fatty acids but also of the series of (ω-1)-hydroxylated fatty acids which also N-acylate R. meliloti Nod factors (38). Similarly, in a concurrent study, it has been shown that R. leguminosarum bv. viciae and Bradyrhizobium sp. ANU289 have evolved NodA proteins with different specificities (39). It has also been reported that R. meliloti NodAB may act preferentially on backbones of at least four residues (18), implying that NodAB may act together with NodC, as part of an enzymatic complex, to determine chitooligosaccharide chain length and specificity of N-acylation.
Acknowledgments
We thank Arlette Savagnac for her help in GC analysis, and Clare Gough for her critical review of the manuscript. This work was supported by the Human frontier Science Programme (Grant RG-378/92) and from the European Communities BIOTECH Programme (Grant PTP CT93-0400).
Footnotes
Abbreviations: NF, Nod factor; IT, infection thread; MALDI, matrix-assisted laser desorption ionization-time of flight.
References
- 1.Fisher R F, Long S R. Nature (London) 1992;357:655–660. doi: 10.1038/357655a0. [DOI] [PubMed] [Google Scholar]
- 2.Dénarié J, Cullimore J. Cell. 1993;74:951–954. doi: 10.1016/0092-8674(93)90717-5. [DOI] [PubMed] [Google Scholar]
- 3.Schultze M, Kondorosi E, Ratet P, Buire M, Kondorosi A. Int Rev Cytol. 1994;156:1–75. [Google Scholar]
- 4.Spaink H P. Annu Rev Phytopathol. 1995;33:345–368. doi: 10.1146/annurev.py.33.090195.002021. [DOI] [PubMed] [Google Scholar]
- 5.Spaink H P, Wijffelman C A, Pees E, Okker R J H, Lugtenberg B J J. Nature (London) 1987;328:337–340. [Google Scholar]
- 6.Horvath B, Bachem C W B, Schell J, Kondorosi A. EMBO J. 1987;6:841–848. doi: 10.1002/j.1460-2075.1987.tb04829.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Dénarié J, Debellé F, Rosenberg C. Annu Rev Microbiol. 1992;46:497–531. doi: 10.1146/annurev.mi.46.100192.002433. [DOI] [PubMed] [Google Scholar]
- 8.Schwedock J, Long S R. Nature (London) 1990;348:644–647. doi: 10.1038/348644a0. [DOI] [PubMed] [Google Scholar]
- 9.Roche P, Debellé F, Maillet F, Lerouge P, Faucher C, Truchet G, Dénarié J, Promé J C. Cell. 1991;67:1131–1143. doi: 10.1016/0092-8674(91)90290-f. [DOI] [PubMed] [Google Scholar]
- 10.Ehrhardt D W, Atkinson E M, Faull K F, Freedberg D I, Sutherlin D P, Armstrong R, Long S R. J Bacteriol. 1995;177:6237–6245. doi: 10.1128/jb.177.21.6237-6245.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Schultze M, Staehelin C, Rohrig H, John M, Schmidt J, Kondorosi E, Schell J, Kondorosi A. Proc Natl Acad Sci USA. 1995;92:2706–2709. doi: 10.1073/pnas.92.7.2706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ardourel M, Lortet G, Maillet F, Roche P, Truchet G, Promé J C, Rosenberg C. Mol Microbiol. 1995;17:687–699. doi: 10.1111/j.1365-2958.1995.mmi_17040687.x. [DOI] [PubMed] [Google Scholar]
- 13.Demont N, Debellé F, Aurelle H, Dénarié J, Promé J C. J Biol Chem. 1993;268:20134–20142. [PubMed] [Google Scholar]
- 14.Ardourel M, Demont N, Debellé F, Maillet F, De Billy F, Promé J C, Dénarié J, Truchet G. Plant Cell. 1994;6:1357–1374. doi: 10.1105/tpc.6.10.1357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Geremia R A, Mergaert P, Geelen D, Van Montagu M, Holsters M. Proc Natl Acad Sci USA. 1994;91:2669–2673. doi: 10.1073/pnas.91.7.2669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Spaink H P, Wijfjes A H M, van der Drift K M G M, Haverkamp J, Thomas-Oates J E, Lugtenberg B J J. Mol Microbiol. 1994;13:821–831. doi: 10.1111/j.1365-2958.1994.tb00474.x. [DOI] [PubMed] [Google Scholar]
- 17.John M, Röhrig H, Schmidt J, Wieneke U, Schell J. Proc Natl Acad Sci USA. 1993;90:625–629. doi: 10.1073/pnas.90.2.625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Atkinson E M, Palcic M M, Hindsgaul O, Long S R. Proc Natl Acad Sci USA. 1994;91:8418–8422. doi: 10.1073/pnas.91.18.8418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Röhrig H, Schmidt J, Wieneke U, Kondorosi E, Barlier I, Schell J, John M. Proc Natl Acad Sci USA. 1994;91:3122–3126. doi: 10.1073/pnas.91.8.3122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kondorosi E, Banfalvi Z, Kondorosi A. Mol Gen Genet. 1984;193:445–452. [Google Scholar]
- 21.Fisher R F, Tu J K, Long S R. Appl Environ Microbiol. 1985;49:1432–1435. doi: 10.1128/aem.49.6.1432-1435.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Marvel D J, Kuldau G, Hirsch A, Richards E, Torrey J G, Ausubel F M. Proc Natl Acad Sci USA. 1985;82:5841–5845. doi: 10.1073/pnas.82.17.5841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Rosenberg C, Boistard P, Dénarié J, Casse-Delbart F. Mol Gen Genet. 1981;184:326–333. doi: 10.1007/BF00272926. [DOI] [PubMed] [Google Scholar]
- 24.Martinez-Romero E, Segovia L, Mercante F M, Franco A A, Graham P, Pardo M A. Int J Syst Bacteriol. 1991;41:417–426. doi: 10.1099/00207713-41-3-417. [DOI] [PubMed] [Google Scholar]
- 25.Debellé F, Rosenberg C, Vasse J, Maillet F, Martinez E, Dénarié J, Truchet G. J Bacteriol. 1986;168:1075–1086. doi: 10.1128/jb.168.3.1075-1086.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Maillet F, Debellé F, Dénarié J. Mol Microbiol. 1990;4:1975–1984. doi: 10.1111/j.1365-2958.1990.tb02047.x. [DOI] [PubMed] [Google Scholar]
- 27.Leong S A, Williams P H, Ditta G S. Nucleic Acids Res. 1985;13:5965–5976. doi: 10.1093/nar/13.16.5965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ditta G, Stanfield S, Corbin D, Helinski D R. Proc Natl Acad Sci USA. 1980;77:7347–7351. doi: 10.1073/pnas.77.12.7347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Somasegaran P, Hoben H J. Handbook for Rhizobia. Berlin: Springer; 1994. [Google Scholar]
- 30.Truchet G, Debellé F, Vasse J, Terzaghi B, Garnerone A M, Rosenberg C, Batut J, Maillet F, Dénarié J. J Bacteriol. 1985;164:1200–1210. doi: 10.1128/jb.164.3.1200-1210.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Sambrook J, Fritsch E F, Maniatis T. Molecular Cloning: A Laboratory Manual. 2nd Ed. Plainview, NY: Cold Spring Harbor Lab. Press; 1989. [Google Scholar]
- 32.Poupot R, Martinez-Romero E, Promé J C. Biochemistry. 1993;32:10430–10435. doi: 10.1021/bi00090a019. [DOI] [PubMed] [Google Scholar]
- 33.Poupot R, Martinez-Romero E, Maillet F, Promé J C. FEBS Lett. 1995;368:536–540. doi: 10.1016/0014-5793(95)00737-t. [DOI] [PubMed] [Google Scholar]
- 34.Faucher C, Camut S, Dénarié J, Truchet G. Mol Plant–Microbe Interact. 1989;2:291–300. [Google Scholar]
- 35.Debellé F, Maillet F, Vasse J, Rosenberg C, De Billy F, Truchet G, Dénarié J, Ausubel F M. J Bacteriol. 1988;170:5718–5727. doi: 10.1128/jb.170.12.5718-5727.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Dénarié J, Debellé F, Promé J C. Annu Rev Biochem. 1996;65:503–535. doi: 10.1146/annurev.bi.65.070196.002443. [DOI] [PubMed] [Google Scholar]
- 37.Krishnan H B, Pueppke S G. Mol Plant–Microbe Interact. 1991;4:512–520. doi: 10.1094/mpmi-4-512. [DOI] [PubMed] [Google Scholar]
- 38.Debellé F, Plazanet C, Roche P, Pujol C, Savagnac A, Rosenberg C, Promé J C, Dénarié J. Mol Microbiol. 1996;22:303–314. doi: 10.1046/j.1365-2958.1996.00069.x. [DOI] [PubMed] [Google Scholar]
- 39.Ritsema T, Wijfjes A H M, Lugtenberg B J J, Spaink H P. Mol Gen Genet. 1996;251:44–51. doi: 10.1007/BF02174343. [DOI] [PubMed] [Google Scholar]