

Abstract
T cells recognize antigen by formation of a trimolecular complex in which the T-cell receptor (TCR) recognizes a specific peptide antigen within the groove of a major histocompatibility complex (MHC) molecule. It has generally been assumed that T-cell recognition of two distinct MHC–antigen complexes is due to similarities in the three-dimensional structure of the complexes. Here we report results of experiments examining the crossreactivity of TCRs recognizing the myelin basic protein peptide MBPp85–99 and several of its analogs in the context of MHC. We demonstrate that single conservative amino acid substitutions of the antigenic peptide at the predominant TCR contact residues at positions 91 and 93 totally abrogate reactivity of specific T-cell clones. Yet, when a conservative substitution is made at position 91 concomitant with a substitution at position 93, the T-cell clones regain reactivity equivalent with that of the original stimulating peptide. Thus, the exact nature of the amino acid side chains engaging one TCR functional pocket may change the apparent selectivity of the other predominant TCR functional pocket, thus suggesting a remarkable degree of receptor plasticity. This ability of the TCR–MHC–peptide complex to undergo conformational changes provides a conceptual framework for reconciling the apparent paradox of the extreme selectivity of the TCR and its remarkable crossreactivity with different MHC–peptide complexes.
T cells recognize a complex formed by peptide antigens and major histocompatibility complex (MHC) molecules (1–3). In recent years, information has been produced examining this complex. Based on x-ray crystallography, the structures of several MHC–peptide complexes have been solved (4–9). Functionally, the determination of MHC allelic motifs, the sequencing of naturally processed MHC ligands, and the evaluation of MHC site directed mutants have all helped in the elucidation of the three dimensional orientation of this complex (reviewed in ref. 10). By contrast, although an x-ray crystallographic structure of the T-cell receptor (TCR) structure engaging the MHC–peptide complex has been solved (11), it is unknown what TCR–MHC structure would allow recognition of two distinct peptides that share little sequence homology, yet would allow, at the same time, the TCR the specificity to differentiate individual atoms on a molecule (12).
The autoantigen myelin basic protein (MBP) is an extensively studied autoantigen in experimental models of autoimmunity as well as in the autoimmune disease multiple sclerosis (13, 14). It is one of the few antigens whose immunodominant epitopes have been mapped in humans (15–17). Moreover, as crossreactivity of T cells to MBP and viral/bacterial antigens has been implicated in the pathogenesis of multiple sclerosis (18–21), it was of interest to study the degree of flexibility in the T-cell response to this antigen.
T-cell clones were generated against either the immunodominant region 85–99 of human MBP or an altered peptide of MBP, in which one of the predominant TCR contact residues, that at position 93, was changed from lysine to arginine. T-cell proliferation and cytokine secretion of the T-cell clones in response to a series of peptides substituted at either one or both of the predominant TCR contact residues (positions 91 and 93) were examined. Any single amino acid substitution at these predominant TCR contact positions abrogated reactivity as measured by proliferation or cytokine secretion. However, a conservative substitution at position 93 of the peptide induced a change in the receptor such that now a substitution of the amino acid side chain at position 91 could induce T-cell activation equivalent to that elicited by the original stimulating peptide. These data are best explained by a model in which the TCR possesses at least two functional pockets (FPs); the exact nature of the side chain engaging one TCR FP may change the apparent selectivity of the other TCR FP. This suggests a remarkable degree of TCR–MHC plasticity.
MATERIALS AND METHODS
Peptide Binding to MHC Molecules.
Peptide HFFR (ENPVVHFFRNIVTPR) and other altered peptides were synthesized in the Biopolymer Laboratory, Harvard Medical School, by automated solid phase methods using fluorenylmethoxycarbonyl (Fmoc)-protected amino acid precursors and purified by reverse-phase HPLC. Peptides were >98% pure on the basis of HPLC and mass spectrometry analysis. The binding of MBP peptides to purified DRβ1*1501 molecules was determined as previously described (22).
Split Well Assay and T-Cell Cloning.
T-cell clones were generated as previously described (17). Briefly, whole mononuclear cells (WMNCs) from the blood were cultured in 96-well plates at 2 × 105 cells per well with peptide HFFR (ENPVVHFFRNIVTPR) for individual Ho and peptide HFFK for individuals Hy and O6, both at a final concentration of 10 μM. Seven days later, the cultures were restimulated with 3.3 × 104 cells per well of irradiated antigen-pulsed antigen-presenting cells (APCs) prepared by incubating 2 × 106 WMNCs with peptide HFFR (10 μM) in 0.2 ml of medium for 2 hr at 37°C. Two days after restimulation (day 9 of culture), interleukin (IL)-2 (Human T-Stim; Collaborative Biomedical Products, Bedford, MA) was added to a final concentration of 5%. Cells were provided with fresh media with IL-2 every 3–4 days. On day 14, a split well assay was performed on each individual well (17). All lines showing reactivity (stimulation index > 2 and Δcpm > 500) were single cell cloned (17).
Proliferation Assay.
The T cells were washed twice and added to U-bottomed 96-well plates (1 × 105cells per well). The APCs were incubated with varying concentrations (0–50 μM) of peptide for 2 hr at 37°C. The EBV-transformed B-cell line MGAR, which is homozygous for DR2-DRβ1*1501, was used as the antigen-presenting cell line for clones Ho.A226, Ho.A240, and 3Ob.1. The EBV-transformed B-cell line 9016, which expresses DR2-DRβ1*1602, was used as the antigen-presenting cell line for clone Hy.152. The APCs were irradiated (5000 rad), washed, and added to the 96-well plate (104 cells per well) in triplicate. After 48 hr, plates were pulsed with 1 μCi of 3H-thymidine per well to measure proliferation. Plates were harvested 18 hr later.
ELISA.
For measurement of interferon (IFN)-γ, IL-10, and IL-4, and IL-5, supernatants of duplicated cultures were collected 40 hr after stimulation using identical conditions as for proliferation assays. Primary and secondary antibodies were purchased from PharMingen and used following the manufacturer’s suggestions. Assays were done in duplicate.
Determination of TCR.
TCR usage was determined as previously described by Southern blotting and hybridization (23) and fluorescence staining. PCR products were ligated into pCRII vectors (TA cloning system; Invitrogen) and sequenced (Sequenase version 2.0 DNA sequencing kit; United States Biochemical) following manufacturer’s suggestions.
RESULTS
Reactivity of T Cells Cloned Against an Altered Peptide Ligand to MBP.
The peptide MBPp85-99 (ENPVVHFFKNIVTPR) is the immunodominant epitope that binds HLA-DR2 (DRβ1*1501; refs. 22, 24, and 25), and the amino acid residues H(90), F(91), and K(93) have been shown to be TCR contact residues (25). T cells from a healthy individual with the DR2 phenotype were cloned by in vitro stimulation with an altered peptide ligand to MBPp85-99 in which the predominant T-cell contact residue at position 93 was changed from a lysine (K) to an arginine (R; ENPVVHFFRNIVTPR). Subsequent single cell cloning was performed with PHA.
Two T-cell clones (Ho.A226 and Ho.A240) were derived independently from separate wells. Both were highly reactive with the HFFR-altered peptide ligand, as measured by proliferation or cytokine secretion (Fig. 1). These clones were not reactive with the native MBPp85-99 peptide, HFFK. To confirm that arginine at position 93 was a predominant TCR contact residue for these clones, a series of peptides were synthesized with substitutions at this position. Ho.A226 and Ho.A240 could not recognize any of the peptides with amino acid substitutions at arginine (93), including the conservative lysine substitution, indicating that position 93 was one of the predominant TCR contact sites (Fig. 1A). The ability of these clones to proliferate in response to a panel of MBPp85-99 peptides substituted at the other presumed TCR contact residue at position 91 was then examined. Similarly, reactivity was lost with both conservative and nonconservative changes in this position, including the substitution of a tyrosine for a phenylalanine, indicating that these T-cell clones could absolutely differentiate between a single hydroxyl molecule on the peptide antigen (Fig. 1B). These results indicate that position 91 is the second predominant contact site for the TCR.
Figure 1.
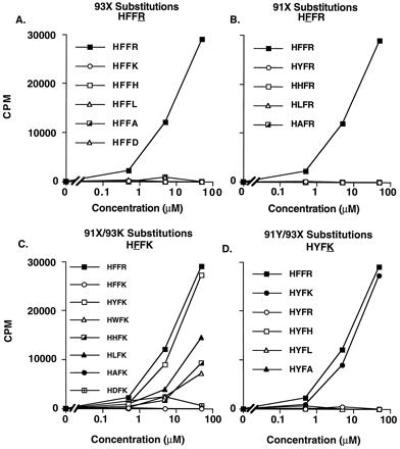
Reactivity of Ho.A226 to a panel of MBPp85-99 peptide antigens. (A) Reactivity to a series of MBPp85-99 peptides substituted at position 93. (B) Reactivity to a series of MBPp85-99 peptides substituted at position 91. (C) Reactivity to a series of MBPp85-99 peptides with a lysine (K) at position 93 and an amino acid substitution at position 91. (D) Reactivity to a series of MBPp85-99 peptides with a tyrosine (Y) at position 91 and an amino acid substitution at position 93. A representative of three experiments using clone Ho.A226 is shown. SEs were <10%. The background proliferation of 12,177 cpm was subtracted from each value shown. Virtually identical results were obtained with clone Ho.A240, which contained an identical TCR sequence (data not shown). Peptides are identified by the core amino acids at positions 90–93.
A second series of peptides were synthesized with double substitutions at predominant TCR contact residues—a lysine at position 93 (HFFK) as well as modifications at positions 90 or 91 (Fig. 1C). To our surprise, the peptide HYFK with the conservative substitutions of a tyrosine for a phenylalanine at position 91 and a lysine for an arginine at position 93 totally restored reactivity of these T-cell clones. Moreover, significant T-cell reactivity was restored with a variety of amino acid substitutions at positions 90 or 91 concomitant with a lysine at position 93. These included a large, bulky side chain such as tryptophan (HWFK), the smaller noncharged aliphatic side chain of leucine (HLFK), or the basic side chain of the histidine (HHFK). The single methyl group side chain of alanine (HAFK) was able to elicit only a minimal response. The substitutions of a lysine (KFFK) or a phenylalanine (FFFK) at position 90 also partially activated the clones (data not shown).
To further determine the influence that predominant TCR contact residues at positions 91 and 93 may have on each other and in subsequent activation of the clones, a series of peptides was synthesized with a tyrosine at position 91 and different amino acids at position 93. The only peptide that was able to activate clones Ho.A226 or Ho.A240 was the HYFK peptide. Thus, the tyrosine at position 91 was only accepted when a lysine was at position 93 (Fig. 1D). These data indicate that with an arginine in position 93, no substitutions could be tolerated at the predominant TCR contact residue at position 91, including a conservative phenylalanine to tyrosine change. However, a lysine in position 93 allowed for both conservative and nonconservative substitutions at the other predominant TCR contact residues, indicating that the total loss of reactivity observed with a conservative change in one predominant TCR contact residue on the antigenic peptide could be completely restored by a second, distinct substitution at another predominant TCR contact residue (Table 1).
Table 1.
Reactivity of T-cell clones to peptide antigens
| Clone | MBPp85–99 sequence | 50% maximum,* μM | Proliferation,† Δcpm |
|---|---|---|---|
| Ho.A226 | HFFR | 12 | 29,008 |
| HFFK | — | 0 | |
| HYFR | — | 0 | |
| HYFK | 20 | 27,292 | |
| Hy.152 | HFFK | 5 | 7854 |
| HFFR | — | 501 | |
| HYFK | — | 1899 | |
| HYFR | 15 | 11,956 | |
| 3Ob.1 | HFFK | 2 | 79,968 |
| HFFR | — | 161 | |
| HYFK | — | 1960 | |
| HYFR | 5 | 49,216 |
Peptide concentrations (in μM) that give 50% maximum proliferation of the primary antigen used to generate the T-cell clone. A dash indicates that no concentration of peptide could induce 50% maximal proliferation.
cpm ([3H]thymidine incorporation) at 50 μM peptide.
An additional T-cell clone, Ho.B27, was isolated from this individual and was tested for its ability to respond when the same complementary mutations were made in the MBP peptide. Unlike clones Ho.A226 and Ho.A240, Ho.B27 only proliferated to the HFFR peptide (data not shown).
HFFR and HYFK Provide Equivalent Signals Through the TCR.
It was possible that certain altered peptide ligands could be activating the T-cell clones to produce cytokines, even though no proliferation was observed, or to switch their cytokine profile. To test this, supernatants were collected after 40 hr of stimulation with the highest concentration (50 μM) of various altered peptides, and IFN-γ, IL-4, and IL-10 levels were measured by ELISA. None of the peptides induced any appreciable IL-4 production, but IFN-γ and IL-10 production correlated specifically with proliferation (Table 2). These results are consistent with the HFFR and HYFK peptides providing an equivalent signal through the TCR.
Table 2.
Peptide binding to MHC DR2 and cytokine secretion by clones in response to different altered peptides
| Substitutions | Peptide binding DRβ1*1501 (IC50%, nM) | Ho.A226 Δ IFN-γ secretion, pg/ml | Ho.A226 Δ IL-10 secretion, pg/ml | ||||
|---|---|---|---|---|---|---|---|
| ⇑ | ⇑ | ⇓ | ⇑ | ||||
| Native | H | F | F | R | 2.0 | 816 | 201 |
| 93X | H | F | F | K | 4.1 | 0 | 0 |
| H | F | F | H | 6.3 | 0 | 0 | |
| H | F | F | L | 2.8 | 0 | 0 | |
| H | F | F | A | 3.5 | 0 | 0 | |
| H | F | F | D | 343 | 0 | 0 | |
| 91X | H | Y | F | R | 22 | 0 | 0 |
| H | H | F | R | 16 | 0 | 0 | |
| H | L | F | R | 5.5 | 0 | 0 | |
| H | A | F | R | 19 | 0 | 0 | |
| 90X/93K | K | F | F | K | 7.0 | 474 | 169 |
| A | F | F | K | 5.2 | 239 | 123 | |
| F | F | F | K | 2.0 | 407 | 157 | |
| D | F | F | K | 76 | 0 | 0 | |
| 91X/93K | H | Y | F | K | 2.1 | 685 | 175 |
| H | W | F | K | 20 | 254 | 115 | |
| H | H | F | K | 7.9 | 237 | 138 | |
| H | L | F | K | 7.6 | 471 | 121 | |
| H | A | F | K | 2.1 | 20 | 123 | |
| H | D | F | K | 423 | 0 | 0 | |
| 91Y/93X | H | Y | F | H | 13 | 0 | 0 |
| H | Y | F | L | 18 | 0 | 0 | |
| H | Y | F | A | 18 | 0 | 0 | |
| 92A* | H | F | A | R | 924 | ||
The binding of MBP peptides to purified DRβ1*1501 molecules and cytokine secretion values after 40 hr of stimulation with antigen (at 50 μM) are reported. The background secretion of IFN-γ, IL-10, or IL-4 by clone Ho.226 in the presence of APCs without antigen was undetectable. There was no measurable IL-4 production. A representative of three experiments is shown. ⇑, TCR contact; and ⇓, MHC contact.
Binding of the peptide HFAR, which has an alanine at residue 92, to MHC DR2 molecules.
Reactivity of T Cells Cloned Against the Native MBP Peptide.
The T-cell clones Ho.A226 and Ho.A240 were generated by stimulating T cells with an altered peptide to the native MBPp85-99 sequence. To further investigate the generality of these observations and to examine the reactivity to native self MBP antigen, T-cell clones reactive with the native MBPp85-99 (HFFK) from another individual expressing HLA-DR2 were generated. A stable T-cell clone, Hy.152, which reacted with the native peptide, was similarly examined for its ability to react to changes in the MBP peptide at positions 93 and 91. As expected, Hy.152 could not recognize peptides in which there was a substitution at position 93, even with a conservative change of an arginine for a lysine, and it showed minimal reactivity to peptides in which position 91 was altered (Fig. 2 A and B). However, when the clone was tested for its ability to tolerate peptides in which there were two amino acid changes, a reciprocal pattern in terms of amino acid substitutions was seen as compared with the clones Ho.A226 and Ho.A240. That is, the substitution of an arginine for a lysine at position 93 allowed for both conservative and nonconservative substitutions at the other predominant TCR contact residue 91 (Fig. 2C). However, with a tyrosine at position 91, only minimal reactivity was observed to peptides not having an arginine at position 93 (Fig. 2D). IFN-γ secretion again correlated with proliferation (data not shown). These data are summarized in Table 1.
Figure 2.
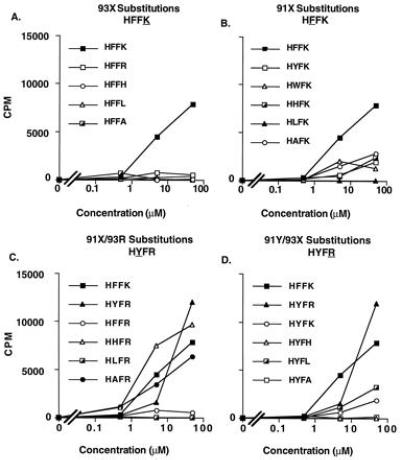
Reactivity of Hy.152 to a panel of MBPp85-99 peptide antigens. (A) Reactivity to a series of MBPp85-99 peptides substituted at position 93. (B) Reactivity to a series of MBPp85-99 peptides substituted at position 91. (C) Reactivity to a series of MBPp85-99 peptides with an arginine (R) at position 93 and an amino acid substitution at position 91. (D) Reactivity to a series of MBPp85-99 peptides with a tyrosine (Y) at position 91 and an amino acid substitution at position 93. A representative of three experiments is shown. SEs were <10%. The background proliferation of 1719 cpm was subtracted from each value shown.
An additional T-cell clone 3Ob.1 was isolated from a third individual expressing HLA-DR2, which reacted with the native peptide MBPp85-99 (HFFK). Clone 3Ob.1 was tested for its reactivity to native peptide as well as a panel of altered peptides. It was not found to react with any peptides in which a single amino acid substitution was made at positions 90, 91, or 93, except for minimal reactivity to an alanine substitution at position 93 (Table 3). Yet, a conservative substitution of an arginine for a lysine at position 93 allowed for both conservative and nonconservative substitutions at position 91 (Table 1). For this clone, a tyrosine at position 91 did allow for some flexibility at position 93. Unlike the previous clones, 3Ob.1 did not secrete IFN-γ in response to stimulation, but IL-4 and IL-5 correlated specifically with proliferation (Table 3).
Table 3.
Reactivity of 3Ob.1 to a panel of MBPp85-99 peptide antigens
| Antigen | Peptide | Antigen, cpm | IL-4 secretion, pg/ml | IL-5 secretion, pg/ml | IL-10 secretion, pg/ml | IFN-γ secretion, pg/ml |
|---|---|---|---|---|---|---|
| No Antigen | — | 1,746 | 0 | 0 | 0 | 0 |
| Native | HFFK | 81,714 | 1,676 | 10,500 | 0 | 83 |
| 90 substitutions | AFFK | 2,125 | 0 | 0 | 0 | 0 |
| FFFK | 5,020 | 0 | 280 | 0 | 0 | |
| KFFK | 3,130 | 0 | 0 | 0 | 0 | |
| 91 substitutions | HYFK | 3,706 | 0 | 290 | 0 | 0 |
| HAFK | 2,898 | 0 | 290 | 0 | 0 | |
| HLFK | 3,585 | 0 | 0 | 0 | 0 | |
| HHFK | 3,349 | 0 | 0 | 0 | 0 | |
| HWFK | 2,952 | 0 | 0 | 0 | 0 | |
| 93 substitutions | HFFR | 1,907 | 0 | 0 | 0 | 0 |
| HFFA | 15,361 | 196 | 2,400 | 0 | 0 | |
| HFFL | 1,633 | 0 | 0 | 0 | 0 | |
| HFFH | 1,671 | 0 | 0 | 0 | 0 | |
| 91 Y/93X substitutions | HYFH | 95,305 | 2,349 | 11,000 | 0 | 0 |
| HYFL | 2,537 | 0 | 280 | 0 | 0 | |
| HYFA | 19,324 | 243 | 2,400 | 0 | 0 | |
| 93 R/91X substitutions | HYFR | 50,962 | 740 | 6,800 | 0 | 0 |
| HAFR | 83,858 | 972 | 10,000 | 0 | 0 | |
| HLFR | 71,215 | 1,049 | 10,000 | 0 | 0 | |
| HHFR | 88,287 | 1,604 | 11,000 | 0 | 0 |
A representative of two experiments using 3Ob.1 is shown. Standard errors were <10%. T cells were tested for their reactivity to peptides at a concentration of 50 μM.
Seven other distinct clones from individual Hy and two other distinct clones from individual Ob using different TCRs were isolated and tested for their ability to respond when the same complementary mutations were made in the MBP peptide. These clones were not able to respond to any peptides with double mutations (data not shown).
Reactivity of Cells Is Due to a Single Clone with a Single TCR.
The apparent crossreactivity observed could be explained by the presence of contaminating T cells recognizing a different peptide in a presumed clonal population or by a T cell possessing two distinct TCRs. These were excluded by the following experiments. First, the T cells were originally isolated at limiting dilution, and then they were recloned twice at 0.3 cells per well. Secondly, we used three methods to exclude contaminating T-cell populations with different TCR sequences. First, the α chain of the TCR was determined by PCR amplification (23) and subsequent Southern blotting. Hybridization with a primer to the α constant chain showed that Ho.A226, Ho.A240, Hy.152, and 3Ob.1 each expressed only a single α chain (Vα 18 for the Ho clones, Vα 1 for the Hy clone, and Vα6 for the Ob clone). The β chain was determined by staining with a panel of antibodies to the variable region. Each clone stained positive for a single β chain (Vβ 8 for the Ho clones, Vβ 5.1 for the Hy clone, and Vβ2 for the Ob clone). Lastly, we directly sequenced the clones to determine whether they were clonally related and to provide further evidence that there were no contaminating T-cell populations. Only one sequence was found for both the α and β chains for clones Ho.A226 and Ho.A240, and they were identical TCRs. Similarly, Hy.152 and 3Ob.1 each expressed only a single, yet, distinct TCR α and β chain. Thus, the reactivities of the clones were due to a single TCR on an individual T-cell clone.
Altered Peptides in the Same Register as the Native MBP.
It was also possible that the HFFR peptide bound in a different register in the DR2 molecule as compared with the native MBPp85-99 and thus would not be structurally similar. This was examined by measuring the MHC binding affinity of the peptides studied, in particular that of the HFFR peptide with a substitution at the presumed MHC contact residue at position 92 (Table 2). As was similar for the native MBPp85-99 peptide, the phenylalanine at position 92 was critical for MHC binding, while substitutions at positions 90, 91, and 93, except that of the highly charged aspartic acid, did not affect MHC binding. In total, these data indicate that all of the peptides bound in the same register to the MHC molecule with approximately the same affinity.
DISCUSSION
We demonstrate that a conservative amino acid substitution at one major TCR contact residue, which totally abrogates T-cell reactivity, can allow for both conservative and nonconservative changes in other TCR contact residues that can fully restore antigen recognition (Table 4). These data lead us to postulate there are at least two functional pockets in TCR recognition of MBPp85–99; FP′1 contacts the amino acid at position 93 while FP′2 contacts the amino acid at position 91. The amino acid side chain engaging one TCR FP changes the selectivity of the other TCR FP, suggesting a remarkable degree of receptor plasticity.
Table 4.
Complementary mutations
| Clone | FP′1 (93) | FP′2 (91) | |
|---|---|---|---|
| ENPVVHFFRNIVTPR* | ENPVVHFFRNIVTPR | ||
| Ho.A226 | R | ⇒ | F |
| K | ⇒ | Y,L,H | |
| K | ⇐ | Y | |
| R | ⇐ | F | |
| ENPVVHFFKNIVTPR† | ENPVVHFFKNIVTPR | ||
| Hy.152 | K | ⇒ | F |
| R | ⇒ | Y,H,A | |
| K | ⇐ | F | |
| R,L | ⇐ | Y | |
| 3Ob.1 | K | ⇒ | F |
| R | ⇒ | Y,L,H,A | |
| K | ⇐ | F | |
| H,R | ⇐ | Y |
The table summarizes the allowable amino acid residues that are able to activate T-cell clones >30% of the original stimulating peptide. The arrows point to which amino acids are allowed in one FP given the corresponding amino acid in the other FP.
Ho.A226 was generated against MBPp85-99 (93R) peptide (HFFR).
Hy.152 and 3Ob.1 were generated against MBPp85-99 peptide (HFFK), the native sequence of MBP.
The crystal structure of a TCR recognizing peptide antigen in the context of class I MHC has recently been solved (11). This TCR combining site was relatively flat except for a deep hydrophobic cavity between the hypervariable CDR3s of the α and β chains. The Vα and Vβ CDR3s straddled the peptide between the helices around the central position of the antigenic peptide. Assuming that the crystal structures of other TCRs recognizing antigen in the context of class II MHC will be similar, we postulate that the large aromatic side chain (Y or F) at position 91 of MBPp85–99 is accommodated in this large hydrophobic pocket, which we assigned as FP′2, and the R or K at position 93 interacts with the undulating surface of the TCR CDR3s, which would be FP′1. This would suggest that the specific amino acid side chains that can engage the hydrophobic cavity between the hypervariable CDR3s of TCR α and β chains and supply the major binding energy for the specific TCR-MHC-peptide interaction can be dictated by charged amino acids interacting with the relatively flat FP′1 of the TCR surface.
It has long been hypothesized that T-cell recognition of two distinct MHC–antigen complexes is due to similarities in the three-dimensional structure of the complexes (26, 27). It has been shown that one can design de novo whole proteins having a predetermined three dimensional topology that mimics the conformation recognized by antibodies of a specific protein by varying multiple amino acids (28). Others have found antibodies that can recognize antigens that appear to have no sequence homology to the original antigen against which they were made (29, 30). Likewise, T cells have been shown to recognize peptides that share little sequence homology (12). Yet, unlike these studies in which many amino acids are changed, our system examines T-cell reactivity to peptides that have had only one or two conservative substitutions made. As a result, one is better able to assess the distinct three-dimensional structure of the peptides.
There are two possible mechanisms to explain the ability of a T cell to accommodate different side chains in one TCR FP when specific conservative changes are made in the peptide at another predominant TCR contact residue. The TCR may undergo a conformational change to accommodate the distinct MHC–peptide complexes. This ability of the TCR to change shape upon ligation to MHC–antigen complexes has already been hypothesized (31). It is also possible that the peptides themselves are able to cause a structural change in the MHC, thereby leading to identical MHC–peptide complexes. Fremont et al. (32) determined the structure of I-Ek with the hemoglobin 64–76 peptide. The P6 residue that was was thought to be a TCR contact residue by virtue of functional studies (33) was in fact found to be buried in the MHC molecule. It is thus possible that amino acids that appear to be TCR contact residues by functional analysis may in fact bind MHC and, in the process, alter the structure of the MHC molecule itself. In the present study, this would seem a less likely possibility since almost all of the peptides tested bind with equal affinity to the MHC. Without x-ray diffraction crystallization data, though, we cannot be certain which receptor is altering its structure. Nevertheless, the ability of double mutated peptides to induce complementary changes in the TCR or the MHC is of importance in understanding the potential flexibility of this trimolecular complex.
As we have only examined one peptide in such detail, it is not possible to determine a general motif for complementation pairs for all peptides; we can only determine a general motif that could be used to identify complementations in peptides with similar amino acid sequences. Specifically, as both large bulky aromatic amino acids and charged amino acids are frequently observed to be TCR contact residues, we postulate that these residues (F/Y amino acid, space, K/R) may form a complementary pair in other antigenic systems as well. Moreover, it would appear likely that this is not the only complementary pair of amino acids that can be recognized by T cells, and a search should continue to identify others.
Our data suggest a remarkable degree of plasticity of the TCR–MHC structure and can potentially explain a number of immunologic phenomena. First, the ability of peptide antigens to induce structural changes in the TCR or MHC provides an explanation for the ability of a T-cell clone to recognize two distinct peptides that share little sequence homology, yet, simultaneously to differentiate individual atoms on the same antigenic peptide. This concept further suggests that an individual TCR can recognize more antigenic complexes than previously thought and indicates that potentially vast numbers of exogenous antigens may be capable of crossreacting with self antigens. This is of obvious importance in understanding the development of autoimmunity. Moreover, it is also possible that there are structural changes in the TCR that allow a single T cell to recognize both allogeneic MHC–peptide and self MHC–peptide complexes. Lastly, it would appear likely that this capacity of recognizing multiple ligands may better explain how a limited number of peptides in the thymus can select for T cells that react to the wide variety of peptide antigens possible.
Acknowledgments
We would like to thank M. Farzan, S. Schreiber, and D. Wiley for helpful discussions. L.J.A. is a Howard Hughes Medical Institute Predoctoral Fellow. This work was supported by a grant from the National Multiple Sclerosis Society and National Institutes of Health Grant RO1-NS24247 to D.A.H.
Footnotes
Abbreviations: MHC, major histocompatibility complex; TCR, T-cell receptor; MBP, myelin basic protein; IL, interleukin; FP, functional pocket; APC, antigen-presenting cells; IFN, interferon.
References
- 1.Zinkernagel R, Doherty P. Nature (London) 1974;251:547–548. doi: 10.1038/251547a0. [DOI] [PubMed] [Google Scholar]
- 2.Katz D H, Hamaoka T, Benacerraf B. J Exp Med. 1973;137:1405–1418. doi: 10.1084/jem.137.6.1405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Rosenthal A J, Shevach E. J Exp Med. 1973;138:1194–1212. doi: 10.1084/jem.138.5.1194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bjorkman P J, Saper M A, Samraoui B, Bennett W S, Strominger J L, Wiley D C. Nature (London) 1987;329:506–512. doi: 10.1038/329506a0. [DOI] [PubMed] [Google Scholar]
- 5.Madden D R, Gorga J C, Strominger J L, Wiley D C. Cell. 1992;70:1035–1048. doi: 10.1016/0092-8674(92)90252-8. [DOI] [PubMed] [Google Scholar]
- 6.Fremont D H, Matsumura M, Stura E A, Peterson P A, Wilson I A. Science. 1992;257:919–927. doi: 10.1126/science.1323877. [DOI] [PubMed] [Google Scholar]
- 7.Matsumura M, Fremont D H, Peterson P A, Wilson I A. Science. 1992;257:927–934. doi: 10.1126/science.1323878. [DOI] [PubMed] [Google Scholar]
- 8.Zhang W Y, Young A C, Imarai M, Nathenson S G, Sacchettini J C. Proc Natl Acad Sci USA. 1992;89:8403–8407. doi: 10.1073/pnas.89.17.8403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Brown J H, Jaretzsky T S, Gorga J C, Stern L J, Urban R G, Strominger J L, Wiley D C. Nature (London) 1993;364:33–39. doi: 10.1038/364033a0. [DOI] [PubMed] [Google Scholar]
- 10.Chien Y, Davis M M. Immunol Today. 1993;14:597–602. doi: 10.1016/0167-5699(93)90199-u. [DOI] [PubMed] [Google Scholar]
- 11.Garcia K C, Degano M, Stanfield R L, Brunmerk A, Jackson M R, Peterson P A, Teyton L, Wilson I A. Science. 1996;274:209–219. [PubMed] [Google Scholar]
- 12.Evavold B D, Sloan-Lancaster J, Wilson K J, Rothbard J B, Allen P M. Immunity. 1995;2:655–663. doi: 10.1016/1074-7613(95)90010-1. [DOI] [PubMed] [Google Scholar]
- 13.Zamvil S, Nelson P, Trotter J, Mitchell D, Knobler R, Fritz R, Steinman L. Nature (London) 1985;317:355–358. doi: 10.1038/317355a0. [DOI] [PubMed] [Google Scholar]
- 14.Mokhtarian F, McFarlin D E, Raine C S. Nature (London) 1984;309:356–358. doi: 10.1038/309356a0. [DOI] [PubMed] [Google Scholar]
- 15.Pette M, Fujita K, Wilkinson D, Altmann D M, Trowsdale J, Giegerich G, Hinkkanen A, Epplen J T, Kappos L, Wekerle H. Proc Natl Acad Sci USA. 1990;87:7968–7972. doi: 10.1073/pnas.87.20.7968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Martin R, Howell M D, Jaraquemada D, Flerlage M, Richert J, Brostoff S, Long E O, McFarlin D E, McFarland H F. J Exp Med. 1991;173:19–24. doi: 10.1084/jem.173.1.19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ota K, Matsui M, Milford E L, Mackin G A, Weiner H L, Hafler D A. Nature (London) 1990;346:183–187. doi: 10.1038/346183a0. [DOI] [PubMed] [Google Scholar]
- 18.Damian R T. Am Nat. 1964;98:129. [Google Scholar]
- 19.Oldstone M B A. Cell. 1987;50:819–820. doi: 10.1016/0092-8674(87)90507-1. [DOI] [PubMed] [Google Scholar]
- 20.Fujinami R S, Oldstone M B A. Science. 1985;230:1043–1046. doi: 10.1126/science.2414848. [DOI] [PubMed] [Google Scholar]
- 21.Wucherpfennig K W, Strominger J L. Cell. 1995;80:695–705. doi: 10.1016/0092-8674(95)90348-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Valli A, Sette A, Kappos L, Oseroff C, Sidney J, Miescher G, Hochberger M, Albert E D, Adorini L. J Clin Invest. 1993;91:616–628. doi: 10.1172/JCI116242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Wucherpfennig K W, Newcombe J, Li H, Keddy C, Cuzner M L, Hafler D. J Exp Med. 1992;175:993–1002. doi: 10.1084/jem.175.4.993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Jaraquemada D, Martin R, Rosen-Bronson S, Flerlage M, McFarland H, Long E O. J Immunol. 1990;145:2880–2885. [PubMed] [Google Scholar]
- 25.Wucherpfennig K W, Sette A, Southwood S, Oseroff C, Matsui M, Strominger J L, Hafler D A. J Exp Med. 1994;179:279–290. doi: 10.1084/jem.179.1.279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Goverman J, Hunkapiller T, Hood L. Cell. 1986;45:475–484. doi: 10.1016/0092-8674(86)90279-5. [DOI] [PubMed] [Google Scholar]
- 27.Nanda N K, Arzoo K K, Geysen H M, Sette A, Sercarz E E. J Exp Med. 1995;182:531–539. doi: 10.1084/jem.182.2.531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Desjarlais J R, Handel T M. Protein Sci. 1995;4:2006–2018. doi: 10.1002/pro.5560041006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.McConnell S J, Kendall M L, Reilly T M, Hoess R H. Gene. 1994;151:115–118. doi: 10.1016/0378-1119(94)90640-8. [DOI] [PubMed] [Google Scholar]
- 30.O’Neil K T, Hoess R H. Curr Opin Struct Biol. 1995;5:443–449. doi: 10.1016/0959-440x(95)80027-1. [DOI] [PubMed] [Google Scholar]
- 31.Janeway J C A. Immunol Today. 1995;16:223–225. doi: 10.1016/0167-5699(95)80163-4. [DOI] [PubMed] [Google Scholar]
- 32.Fremont D H, Hendrickson W A, Marrack P, Kappler J. Science. 1996;272:1001–1004. doi: 10.1126/science.272.5264.1001. [DOI] [PubMed] [Google Scholar]
- 33.Evavold B D, Allen P M. Science. 1991;252:1308–1310. [PubMed] [Google Scholar]