

Abstract
To formally test the hypothesis that the granulocyte/macrophage colony-forming unit (GM-CFU) cells can contribute to early hematopoietic reconstitution immediately after transplant, the frequency of genetically modified GM-CFU after retroviral vector transduction was measured by a quantitative in situ polymerase chain reaction (PCR), which is specific for the multidrug resistance-1 (MDR-1) vector, and by a quantitative GM-CFU methylcellulose plating assay. The results of this analysis showed no difference between the transduction frequency in the products of two different transduction protocols: “suspension transduction” and “stromal growth factor transduction.” However, when an analysis of the frequency of cells positive for the retroviral MDR-1 vector posttransplantation was carried out, 0 of 10 patients transplanted with cells transduced by the suspension method were positive for the vector MDR-1 posttransplant, whereas 5 of 8 patients transplanted with the cells transduced by the stromal growth factor method were positive for the MDR-1 vector transcription unit by in situ or in solution PCR assay (a difference that is significant at the P = 0.0065 level by the Fisher exact test). These data suggest that only very small subsets of the GM-CFU fraction of myeloid cells, if any, contribute to the repopulation of the hematopoietic tissues that occurs following intensive systemic therapy and transplantation of autologous hematopoietic cells.
The advent of CD34 selection has produced extensive clinical testing of the correlation between the rate and completeness of engraftment and the content of CD34+ cells in a collection of autologous hematopoietic cells (1–3). The dose of CD34+ cells that will generate complete and rapid engraftment is on the order of 2 × 106 CD34+ cells per kg of total body weight (4). Most investigators agree that only 5–10% of the CD34+ cells at the most may contain engrafting cells, and that the early cells, which are CD34+CD38−CD33−, are the major contributors to engraftment. In fact, highly fractionated hematopoietic cells, such as CD34+ CD33− lineage CD38−Thy1+Lo are thought to be highly enriched in the cells that contribute to long-term engraftment (5).
Cells that are later in the myeloid maturation sequence (CD34+CD33+) are thought by most individuals not to contribute to long-term engraftment. The granulocyte/macrophage colony-forming unit (GM-CFU) is defined by the cells that grow in methylcellulose into colonies within 7–14 days after innoculation. The phenotype that is being measured by this assay is anchorage-independent growth of clonogenic myeloid hematopoietic precursor cells. The GM-CFU population itself is also a very heterogeneous population, which contains more or less mature cells, with varying proliferative potential. In collections of hematopoietic cells from patients during the recovery from conventional dose chemotherapy, the ratio of these early to late cells can vary from patient to patient, and thereby give different results in the GM-CFU assay.
Disagreement exists over how important the GM-CFU cells are for short-term posttransplant engraftment. In the past, transplantation specialists have used the GM-CFU content to test for the engraftment adequacy of a collection of autologous hematopoietic stem cells. Like the total nucleated cell count, the GM-CFU content is not a very proximate surrogate predictor of the content of hematopoietic cells. Weissman and collaborators (6) fractionated marrow into very early hematopoietic stem cells and non-stem cell fractions. They then used a competitive reconstitution assay to study the role of very early hematopoietic stem cells versus the non-hematopoietic stem cell in early and long-term hematopoietic recovery in lethally irradiated mice (6). The data developed in their mouse model suggest that the day 11–14 recovery as well as the long-term engraftment was generated by very early hematopoietic stem cells. In addition, their data suggest that the more mature hematopoietic cell fractions, to which the majority of the GM-CFU might belong, do not even contribute to short-term engraftment (6).
Clearly, the absence of a clear-cut correlation between the GM-CFU content and the rate of recovery after transplantation of autologous cells, although attributable to the varying ratio of early to late cells in patients postchemotherapy who are undergoing stem cell or marrow collections, has called into question the importance of the GM-CFU to short-term recovery (7).
To resolve this question, we decided to analyze the data derived from an ongoing MDR-1 chemoprotection trial for advanced breast and ovarian cancer patients to test whether the genetically modified cells in the GM-CFU population are actually contributing to the posttransplant repopulation of peripheral blood and marrow hematopoietic cells. We first measured the frequency of the cells that were genetically modified immediately posttransduction using two different transduction protocols: (i) one protocol involved suspension of the CD34 selected cells in the retroviral supernatant for 6 hr and then cryopreservation (this is designated “the suspension protocol”), and (ii) another transduction protocol, which involved the innoculation of the CD34 selected cells on irradiated stromal monolayers in the presence of growth factors interleukins (IL) 3 and 6 for 24 hr, and four feedings that involved replacement of one-half the volume of tissue culture medium with fresh retroviral supernatant supplemented with the IL-3 and IL-6 growth factors, and an additional 24 hr in medium, stromal cells, and growth factors but no viral particles. This is called the “stromal transduction protocol.”
The data from this study showed that the frequency of transduction (as measured by a GM-CFU plating assay in taxol) immediately posttransduction was the same in the suspension and stromal transduction protocols. In contrast, the genetically modified cells posttransplant (as measured by PCR assay) were found only in the patients transplanted with cells transduced by the stromal but not the suspension method. These data suggest that the GM-CFU cells from the suspension method are not contributing to the hematopoietic recovery immediately posttransplantation.
MATERIALS AND METHODS
Selection of Patients and Treatment Protocol.
The MDR-1 genetic chemoprotection trials were as described (8, 9). Patients who relapsed after surgery who were then only partially responsive to salvage conventional dose chemotherapy were exposed to conventional dose chemotherapy as follows: for ovarian cancer patients, 180 mg/m2 VP16 (etoposide) as an intravenous bolus administration (IVB) once a day (qd) for 5 days and 600 mg/m2 cyclophosphamide IVB qd for 5 days; for breast cancer patients, a single dose of 4 g/m2 cyclophosphamide IVB. Engrafting doses of peripheral blood (breast cancer patients) or marrow (ovarian cancer patients) are collected, and CD34 cells were selected using CellPro (Bothell, WA) Ceprate SC Selector Monoclonal Antibody Column Chromatography and transduced with a MDR-1 safety-modified retrovirus. Then the patients receive intensive combination chemotherapy: for ovarian cancer patients, 1.7 g/m2 cyclophosphamide IVB qd for 3 days and 200 mg/m2 thiotepa IVB qd for 3 days; for breast cancer patients, 1.5 g/m2 cyclophosphamide IVB qd for 3 days, 200 mg/m2 thiotepa IVB qd for 3 days, and 150 mg/m2 N,N′-bis(2-chloroethyl)-N-nitrosourea (BCNU) IVB qd for 3 days. Seven days after the conclusion of the chemotherapy, mixtures of the MDR-1 modified and unmodified cells were then transplanted into the patients. Granulocyte colony-stimulating factor (5 μg/kg qd) was given subcutaneously starting 24 hr after the conclusion of the pre-stem cell chemotherapy or 24 hr following the transplant until full recovery was achieved.
In the case of the breast cancer patients, the peripheral blood cells were screened for the presence of neoplastic cells by an immunohistochemical assay, which is sensitive to one neoplastic cell in one million normal cells (10). If this test was positive, the collection would not be used for the transplant.
Collection and Transduction of Hematopoietic Cells.
Marrow cells (in ovarian cancer patients) and peripheral blood hematopoietic cells (in breast cancer patients) were collected when the white cell count reached 2000 cells per mm3 during the early phase of graunolcyte colony-stimulating factor-driven recovery from conventional dose chemotherapy-induced myelosuppression, and then processed through a CS3000 Cobe Laboratories (Lakewood, CO) Continuous Flow Cell Separator as described (11–13). The cells were then CD34 selected using the CellPro Ceprate SC Selector, and then subjected to one of the two following transduction protocols: (i) Suspension protocol: the CD34 selected cells were placed in 150-ml DuPont Cell Culture air-porous bags and suspended in the MDR-1 retroviral supernatant at a multiplicity of infection of at least 0.25 in the presence of protamine sulfate (4 μg/ml). The retroviral vector titer in the supernatant was 1 × 105 colony-forming units per cc. The incubation was carried out for 4–6 hr at 37°C on a rocking platform in a CO2 tissue culture incubator as described previously (11–12, 14, 15). (ii) Stromal growth factor protocol: The CD34 selected cells were inoculated on irradiated autologous stromal monolayers. The stromal cells were obtained from each patient by a diagnostic percutaneous bone marrow aspirate at least 2 weeks before the administration of the pre-stem cell collection chemotherapy. The bone marrow cells were then subjected to Ficoll/Hypaque (Pharmacia) density gradient centrifugation, cultured for 7–10 days until they were 70% confluent, trypsinized, replated in twice the number of T75 flasks, grown again to 70% confluency, and irradiated with 1500 cGy. The CD34 selected cells, obtained from a preparative bone marrow storage or from a leukapheresis, were then innoculated onto the autologous irradiated stromal monolayer for 24 hr in medium supplemented with 100 units/cc of both human IL-3 and IL-6 (National Cancer Institute, Bethesda). This system was then incubated at 37°C in a CO2 tissue culture incubator for 24 hr. Then one-half of the volume was removed, and the cells were spun down and then resuspended in an equal volume of fresh retroviral supernatant, which was supplemented with IL-3 and IL-6 (100 units/ml), and used to replace the volume removed. This was done every 12 hr for four feedings. The cumulative multiplicity of infection was at least 0.25. Twelve hours after the last feeding with retroviral supernatant, all of the medium was removed, and the cells were centrifuged and resuspended in tissue culture medium (MyeloCult H5100, Stem Cell Technologies, Vancouver) that was supplemented with IL-3 and IL-6 (100 units/ml). When this was completed, the supernatant cells were collected.
The monolayer was lightly trypsinized (trypsin 0.25%, for 2.5 min at room temperature), the non-adherent cells were removed, and the total stromal monolayer was trypsinized to recover all of the cells. The cells were then cryopreserved. Both transduction methods are presented in Fig. 1.
Figure 1.
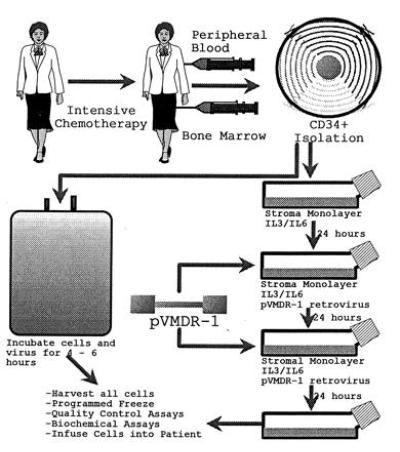
Transduction methods.
In Situ DNA PCR Assay.
The details of this assay have been reported (16). Hematopoietic cells were cytospun on silanized slides (Oncor) using the Shandon cytocentrifuge. The cells were then fixed overnight using buffered formalin (Sigma) at pH 7.0. The slides were treated with trypsinogen (Sigma) at 2 mg/ml for 10–15 min, dehydrated with ethanol, and then air dried. The sense and antisense primers for this assay were selective for the retroviral MDR-1 cDNA, on the basis of being in exons 20 and 22, respectively, which encompass very large introns, thus making the amplification of the MDR-1 endogenous gene sequences very unlikely. The substrate nucleotides were conjugated with digoxigenin (DIG-11-UTP, Boehringer Mannheim). The reaction was hot-started by incubating at 94°C for 4 min. The denaturing temperature was 94°C for 1 min, the elongation temperature was 72°C for 1 min, and the annealing temperature was 60°C for 1 min. Thirty cycles were used for the amplification. Following the 30 cycles, the slides were rinsed and then incubated with an antibody that recognized digoxigenin. The antibody was conjugated with an enzyme for the development of a nitroblue tetrazolium assay. Thus, the nuclei of the cells that are transduced turn blue, while the untransduced are counterstained pink. The slides are then analyzed using a light inverted microscope. The primers used for this DNA PCR assay are as follows: 5′ sense primer in the 5′ → 3′ direction: GATGCTGCTCAAGTTAAAGGG; 3′ antisense primer in the 3′ → 5′ direction: GGCTTGGCAACAAAGAAACTG.
GM-CFU Plating Efficiency Assay in Methylcellulose Supplemented with Taxol.
This assay was as described (15, 17, 18). The marrow cells from the patients were collected before each course of taxol and then fractionated on a Ficoll/Hypaque gradient. The mononuclear cells were then inoculated into 1.3 cc of methylcellulose (MyeloCult H4433, Stem Cell Technologies) at a dose of 100,000 cells per cc in the presence and absence of taxol (8–16 ng/ml) (Calbiochem) in small Petri dishes. The individual 1.3-cc culture dishes were placed in 100-mm Petri dishes along with 5–6 other dishes. One or two of these dishes just contained sterile water without a cover for humidification. After the lid was placed on the 100-mm Petri dish, it was introduced into a 37°C humidified water-jacketed tissue culture incubator and cultured for 2–3 weeks. Growing colonies in this semisolid medium were counted, analyzed, and picked at either 10, 15, or 20 days. PCR assays were further performed on the resistant colonies to confirm the presence of the pVMDR-1 transgene. The transduction frequency in the colony-forming cells (GM-CFU) in taxol was calculated by subtracting the background frequency of colony-forming cells untransduced in taxol (corrected for the plating efficiency in drug-free medium) from the frequency of colony-forming cells in drug (corrected for the plating efficiency in drug-free medium).
Solution DNA PCR Assay for the pVMDR-1 Transgene.
The details of this assay were reported previously (17, 18). A guanidinium extraction solution was used to lyse cells from the bone marrow or peripheral blood of patients 24 hr after the 4- or 48-hr transduction, or from the bone marrow or peripheral blood at various time intervals posttransplantation. The primers for the vector MDR-1 were added in solution for the first round of amplification as follows: 5′ sense primer in the 5′ → 3′ direction: AACGTCGGATGGCCGCGAGAC; the 3′ antisense primer in the 3′ → 5′ direction: GCTCCTTGGAACGGCCACCAA. The 3′ primer for the endogenous MDR-1 was the same as that for the vector MDR-1. The 5′ primer for the endogenous MDR-1 was in the 5′ → 3′ direction: CCGGGAGCAGTCATCTGTGGT. The vector 5′ primer was homologous to gag sequences present in the 5′ untranslated region of the vector mRNA but not found corresponding the region of the endogenous MDR-1 gene; the 5′ endogenous primer was homologous to sequences from the 5′ untranslated region of the native MDR-1 endogenous mRNA, which are not present in the corresponding region of the vector 5′ untranslated region. These primers were reported previously (18). The annealing was conducted at 63°C for 30 sec; the denaturation was conducted at 94°C for 30 sec. There were 20 cycles. Then, 2 μl of the reaction mixture was then mixed with 48 μl of the second round of the amplification reaction mixture, which contained the primers as follows: 5′ sense primer for the vector MDR-1 in the 5′ → 3′ direction: CGGCACCTTTAACCGAGAC; 3′ antisense vector MDR-1 primer in the 3′ → 5′ direction: CACCAAGACGTGAAATCTTG. The 3′ primers for the endogenous MDR-1 was identical to that for the vector. The 5′ primers for the endogenous MDR-1 is AGGCTGATTGGCTGGGCAGGA. There were 40 rounds for the second amplification cycle, which was conducted under the identical conditions.
RESULTS
Percent of Vector MDR-1 Positive Hematopoietic Cells in the Immediate Posttransduction Period.
Direct DNA solution PCR was used to test for the presence of the expression of the retroviral MDR-1 transgene in marrow or peripheral blood mononuclear cells after the transduction of the hematopoietic cells. As shown in Table 1 and in Fig. 2, 10 of 10 samples from the stromal transduction were positive for the vector MDR-1 gene in the immediate posttransduction period with a single round of 40 cycles of solution PCR amplification, without nesting, whereas only two-sixths of the evaluable samples from the suspension transduction were positive with single round PCR. Seven-eighths of these samples were positive with the nested PCR assay (see Table 1).
Table 1.
Pretransplant transduction efficiency
| Patient no. | Transduction sample no. | Cancer type | In situ PCR | Plating efficiency | Solution PCR
|
|
|---|---|---|---|---|---|---|
| Without nesting | With nesting | |||||
| Suspension | Suspension | Suspension | Suspension | |||
| O1 | 1 | Ovary | ND | 2.0 | + | + |
| O2 | 2 | Ovary | ND | 8.4 | NE | NE |
| O3 | 3 | Ovary | ND | 5.7 | − | + |
| O4 | 4 | Ovary | ND | 16.5 | ND | + |
| O5 | 5 | Ovary | 3.0 | 5.7 | ND | + |
| O6 | 6 | Ovary | 1.2 | 20.1 | − | + |
| O9 | 7 | Ovary | 1.9 | 6.0 | + | ND |
| B1 | 8 | Breast | ND | 2.5 | ND | + |
| B2 | 9 | Breast | ND | 18.1 | − | + |
| B3 | 10 | Breast | 5.3 | 2.3 | − | + |
| Mean | 2.8 | 8.7 | ||||
| Median | 1.9 | 5.7 | ||||
| Stromal | Stromal | Stromal | Stromal | |||
| O7 | 1 | Ovary | 3.6 | 8.1 | + | ND |
| O8 | 2 | Ovary | 1.8 | 9.4 | + | ND |
| O10 | 3 | Ovary | 8.1 | 2.1 | + | ND |
| B4 | 4 | Breast | 6.9 | −0.22 | + | ND |
| B5 | 5 | Breast | 9.0 | 14.7 | + | ND |
| B6 | 6 | Breast | 3.0 | 4.3 | + | ND |
| B7 | 7 | Breast | 2.5 | 2.2 | + | ND |
| B8 | 8 | Breast | 8.9 | 2.7 | + | ND |
| B9 | 9 | Breast | 3.7 | 1.0 | + | ND |
| B10 | 10 | Breast | 6.1 | 1.0 | + | ND |
| Mean | 5.4 | 4.5 | ||||
| Median | 6.1 | 2.7 | ||||
The method of transduction (suspension or stromal) is indicated above each column of data. ND, not done; NE, not evaluable.
Figure 2.
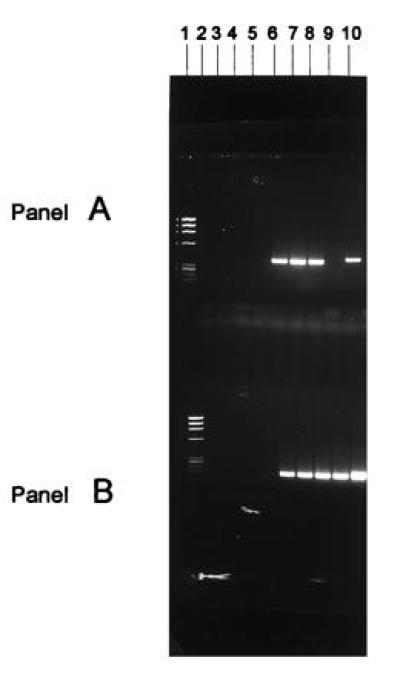
Solution DNA PCR analysis of vector MDR-1 positive cells. The cells from the suspension transduction protocol were incubated for an additional 24 hr in the presence of IL-3 and IL-6 growth factors in suspension culture after 6 hr of incubation with the vector before PCR assay. Cells that were subjected to the stromal or the suspension transduction procedures were cultured for an additional 24 hr after exposure to the retroviral vector in long-term culture on stromal monolayers before being collected for PCR assay. Cells were also obtained from the peripheral blood or bone marrow of patients at various times after transplant. DNA solution PCR analysis of vector MDR-1 sequences in the cells transduced with a MDR-1 retroviral vector. (A) DNA solution PCR analysis of vector MDR-1 sequences; (B) DNA solution PCR analysis of endogenous MDR-1 sequences. Lanes: 1, Phi-X174 molecular weight markers; 2, PCR buffer negative control; 3 and 4, marrow cells from ovarian cancer patient no. 9 unexposed and exposed to the MDR-1 vector suspension transduction method (both negative for the vector sequences); 5 and 6, nonadherent and adherent CD34+ cells from stromal cultures of breast cancer patient no. 5 not exposed to the MDR-1 vector (negative for MDR-1 vector sequences); 7–9, nonadherent hematopoietic cells, mixture of adherent hematopoietic cells and stromal cells, and stromal cells from cultures used for transduction with the MDR-1 vector stromal transduction method from breast cancer patient no. 5, respectively (all three samples positive for the vector MDR-1 sequences); 10, positive control consisting of cDNA from K562 cells known to contain the MDR-1 vector sequences.
This difference is statistically significantly different at the P < 0.0001 level by the Fisher exact test. Suspension transduction involves culturing of the target cells in the retroviral supernatant for 6 hr, whereas stromal transduction involves culturing the CD34 selected cells on stromal monolayers in pVMDR1 retroviral vector supplemented with IL-3 and IL-6.
We also used in situ PCR to test for the presence of the vector MDR-1 DNA in the hematopoietic cells immediately after transduction. The mean transduction frequency for cells transduced with the suspension transduction procedure was 2.8%, whereas the mean for the stromal transduction procedure was 5.6% (Table 1). An example of the in situ PCR is shown in Fig. 3.
Figure 3.
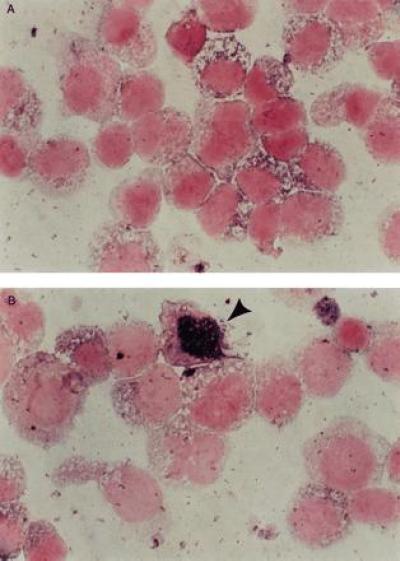
In situ DNA PCR analysis of marrow or peripheral blood cells 24 hr posttransduction or from patients at various time intervals posttransplant. A Boehringer Mannheim kit was used for an in situ DNA PCR assay as described previously (16). The nuclei of the cells that are transduced turn blue, whereas those that are not transduced stay white. Controls that were run with every patient sample included cells known to be negative for the vector pVMDR-1 and the patient’s own pretransduction cells. The percent of cells positive in these negative controls (which is probably due to a primer independent repair incorporation reaction) was subtracted from the percent of cells positive for the blue intranuclear color in known negative cells (16). In situ DNA PCR analysis of negative control cells (A) and cells transduced by the stromal transduction method (B). The positive cells contain intensely staining blue nuclei.
We also analyzed the transduction frequency by the difference in the plating efficiency of hematopoietic cells before and after transduction in methylcellulose supplemented with taxol for transduced and untransduced hematopoietic cells. The calculation of this percentage is presented in the methods section. As can be seen from Table 1, all but one of the GM-CFU samples analyzed showed positivity for increased resistance to taxol in the immediate posttransduction period. There was no difference between the suspension and the stromal transduction methods when assayed in the methylcellulose plating system for resistance to taxol.
As shown in Table 2, the transduction frequencies, as measured by plating efficiencies in the GM-CFU fraction, were totally overlapping.
Table 2.
Pretransplant transduction efficiency as judged by plating efficiency in methylcellulose supplemented by taxol
| Suspension | Stromal |
|---|---|
| 20.1 | 14.7 |
| 18.1 | 9.4 |
| 16.5 | 8.1 |
| 8.4 | 4.3 |
| 6.0 | 2.7 |
| 5.7 | 2.2 |
| 5.7 | 2.1 |
| 2.5 | 1.0 |
| 2.3 | 1.0 |
| 2.0 | −0.22 |
| 8.7 | 4.5 |
A comparison of the transduction frequencies by assaying the plating efficiency of GM-CFU in methylcellulose supplemented by taxol. It is to be noted that this assay also documents expression of the vector GM-CFU in the myeloid progenitor cells, because expression of the vector MDR-1 transgene is necessary for increased plating efficiency. The method of the plating assay is summarized in text.
Positivity of the Hematopoietic Cells for the PVMDR-1 Transgene in the Cells from Patients Transplanted with Cells Transduced by the Suspension Versus the Stromal Transduction Procedure.
Following the transduction and cryopreservation of the cells and completion of the quality control tests on the transduced cells, the patients were treated with intensive systemic therapy designed to reduce the level of the tumor cells in the patients. Seven days after completion of the chemotherapy, an equal mixture of cells that had been exposed to the transduction conditions and those that had not been exposed to the transduction conditions were infused into the patients. Three to four weeks following transplantation, at a time when the white cell count was 4000 cells per mm3 or more and at the time of the administration of the first course of posttransplantation taxol, cells were collected for the direct solution DNA PCR assay and the in situ DNA PCR assays.
As shown in Table 3 and Fig. 4, the cells collected from none of the patients (0/10) transplanted with cells transduced by the suspension transduction protocol were positive. In contrast, 5 of 8 of evaluable patients who were transplanted with the cells transduced by the stromal transduction protocol were positive by the solution DNA PCR assay (see Fig. 4), and 4 of 4 of the samples tested from this group by the in situ DNA PCR assay were positive. This difference in the solution DNA PCR assays was statistically significantly different at the P = 0.0065 level by the Fisher exact test.
Table 3.
Posttransplant DNA PCR analysis of cells from bone marrow of patients transplanted with cells exposed to the suspension or stromal transduction protocol
| Sample no. | Transduction procedure | Patient no. | Nested PCR analysis (solution) |
|---|---|---|---|
| 1 | Suspension protocol | O1 | − |
| 2 | Suspension protocol | O2 | − |
| 3 | Suspension protocol | O3 | − |
| 4 | Suspension protocol | O4 | − |
| 5 | Suspension protocol | O5 | − |
| 6 | Suspension protocol | O6 | − |
| 7 | Suspension protocol | O9 | − |
| 8 | Suspension protocol | B1 | − |
| 9 | Suspension protocol | B2 | − |
| 10 | Suspension protocol | B3 | − |
| 1 | Stromal protocol/GF | O7 | + |
| 2 | Stromal protocol/GF | O8 | ND |
| 3 | Stromal protocol/GF | O10 | + |
| 4 | Stromal protocol/GF | B4 | + |
| 5 | Stromal protocol/GF | B5 | + |
| 6 | Stromal protocol/GF | B6 | + |
| 7 | Stromal protocol/GF | B7 | − |
| 8 | Stromal protocol/GF | B8 | − |
| 9 | Stromal protocol/GF | B9 | − |
| 10 | Stromal protocol/GF | B10 | ND |
GF, growth factors; ND, not done; O, ovarian; B, breast.
Figure 4.
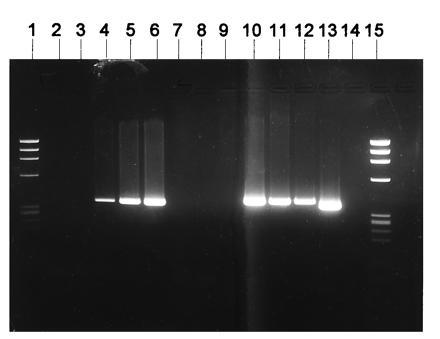
DNA solution PCR analysis of marrow cells from patients transplanted with cells exposed to the stromal transduction method. Lanes: 1, Phi-X174 molecular weight markers; 2, negative buffer PCR control; 3–6, 100,000 K562 cells untransduced mixed with 0,5,20 and 100 K562 cells known to be positive for the vector MDR-1 sequences; 7–9, negative cells; 10 and 11, bone marrow cells from two breast cancer patients collected after transplantation with CD34+ cells exposed to the stromal MDR-1 vector transduction method (positive for MDR-1 vector sequences); 12, peripheral blood cells from one ovarian cancer patient collected after transplantation with CD34+ cells exposed to the stromal MDR-1 vector transduction method (positive for MDR-1 vector sequences); 13, positive control; 14, buffer PCR negative control; 15, Phi-X174 molecular weight markers.
DISCUSSION
Transduction frequencies estimated pretransplant, using the plating efficiency of the GM-CFU in methylcellulose supplemented with taxol, were overlapping in the suspension and the stromal transduction methods. As shown in Table 1, however, the frequency of positivity for vector MDR-1 after the first round of the nested DNA PCR was lower in the case of the suspension transduction method than it was in the stromal transduction method, although all of the samples from the suspension as well as from the stromal transduction protocol were positive by nested solution DNA PCR assay posttransduction but before transplant.
The results of methylcellulose plating assays for the GM-CFU precursor cells pretransplant showed increased resistance of the cells to taxol immediately posttransduction, and that the frequency of transduction using the stromal and suspension transduction procedures were indistinguishable. In contrast, the results of the PCR assays for the vector MDR-1 posttransplantation suggested that only the cells of patients transplanted with cells transduced by the stromal transduction protocol were positive by the solution DNA PCR assay in posttransplantation cells. None of the cells transplanted with cells transduced by the suspension transduction protocol were positive for the vector MDR-1 transgene posttransplant.
These results indicate that although the pretransplant GM-CFU cells were modified as frequently by the suspension method of transduction as by the stromal transduction method, the majority of the cells modified by the suspension method did not repopulate the intensively treated patients sufficiently to show up as positives in the posttransplantation PCR assays. In contrast, the stromal transduction procedure generated cells in the majority of cases that were capable of promoting the reconstitution of the intensively treated patients with genetically modified cells. These data indicate that the genetically modified GM-CFU from the suspension transduction procedure do not repopulate the hematopoietic tissue sufficiently to be detectable by PCR assays in the immediate posttransplantation period.
The question may be asked as to why the stromal transduction protocol generates modified cells that can repopulate the hematopoietic tissues of the recipients, whereas the solution transduction protocol cannot. It is possible that the conditions of the stromal monolayers may elicit a replication event in the target hematopoietic cells that serves to incorporate the MDR-1 retroviral vector, whereas the suspension transduction method does not support the survival of the reconstituting cells and that the replication-dependent integration of the MDR-1 retroviral transcription unit into cells, which can reconstitute the hematopoietic tissues of the recipient after intensive therapy, does not occur at sufficiently high levels to be detected. It is still possible that a very small subset of GM-CFU could be contributing to short term reconstitution, but that this very small subset could only be detected at much higher transduction frequencies than were achievable in these experiments.
It is to be noted that in the suspension transduction method, although the exposure time of the cells to the viruses is only 4–6 hr, the cells are incubated for an additional 24 hr in the absence of stromal cells but in the presence of the IL-3 and the IL-6 before freezing or analysis. It is possible that the absence of the stromal cells during this period adversely affects the survival or the proliferation of reconstituting cells necessary for engraftment of the genetically modified cells.
These data indicate that the majority of GM-CFU do not contribute measurably to posttransplant reconstitution. In addition, it also suggests that programs that require integration of retroviral transgenes into cells that can repopulate the hematopoietic tissue of intensively treated patients will require incubation of the target cells under conditions that will both preserve their reconstitutive capabilities as well as elicit replicative events for the integration of the transgenes into chromosomal DNA. These data may be generally useful for other workers attempting to use bone marrow or peripheral blood transplantation as a method for introducing gene products into patients for the therapy of human disease.
Acknowledgments
We thank Joyce Palmer and Rosemarie Lauzon for editorial assistance; Drs. David Parkinson, Langdon Miller, Mario Sznol, and Al Fallavolitta of the Chemotherapy Evaluation Branch of the National Cancer Institute of the National Institutes of Health; and the staff of the Sandoz Pharmaceutical Corporation for the gift of IL-3 and IL-6. We acknowledge support to A.B.D. from the Anderson Chair for Cancer Research, from the Bush Leukemia Research Fund, National Cancer Institute Grant PO1 CA49639 (The Therapy of chronic myelogenous leukemia), from Ingenex Inc., from The Hull Fund, and from The Ensign Professorship at Yale.
Footnotes
Abbreviations: GM-CFU, granulocyte/macrophage colony-forming unit; IL, interleukin; IVB, intravenous bolus administration; qd, once a day.
References
- 1.To L B, Dyson P G, Juttner C A. Lancet. 1986;ii:404–405. doi: 10.1016/s0140-6736(86)90096-6. [DOI] [PubMed] [Google Scholar]
- 2.Siena S, Gregni M, Brando B, Ravagnani F, Bonadonna G, Gianni A M. Blood. 1989;74:1905–1914. [PubMed] [Google Scholar]
- 3.Juttner C A, To L B. In: Fifth International Symposium on Autologous Bone Marrow Transplantation. Dicke K A, Armitage J O, Dicke-Evinger M J, editors. Arlington, TX: The Cancer Research and Educational Institute; 1990. pp. 483–486. [Google Scholar]
- 4.Berenson R J, Andrews R G, Bensinger W I, Kalamasz D, Knitter G, Spitzer G, Buckner C D, Berstein I D. Blood. 1991;77:1717–1772. [PubMed] [Google Scholar]
- 5.Murray L, Chen B, Galy A, Chen S, Rushinski R, Uchida N, Negrin R, Tricot G, Jagannath S, Vesole D, Barlogie B, Hoffman R, Tsukamoto A. Blood. 1995;85:368–378. [PubMed] [Google Scholar]
- 6.Uchida N, Aguila J L, Fleming W H, Jerabek L, Weissman I L. Blood. 1994;83:3758–3779. [PubMed] [Google Scholar]
- 7.Lansdorp P M. Exp Hematol. 1995;23:187–191. [PubMed] [Google Scholar]
- 8.Deisseroth A B, Kavanagh J, Champlin R. Hum Gene Ther. 1994;5:1507–1522. doi: 10.1089/hum.1994.5.12-1507. [DOI] [PubMed] [Google Scholar]
- 9.Deisseroth A B, Holmes F, Hortobagyi G, Champlin R. Hum Gene Ther. 1996;7:401–416. doi: 10.1089/hum.1996.7.3-401. [DOI] [PubMed] [Google Scholar]
- 10.Osborne M P, Asina S, Wong G Y, Old L J, Cote R. J Cancer Res. 1989;49:2510–2513. [PubMed] [Google Scholar]
- 11.Kantarjian H M, Talpaz M, Hester J, Feldman E, Korbling M, Liang J, Rios M B, Calvert L, Deisseroth A B. J Clin Oncol. 1995;13:553–559. doi: 10.1200/JCO.1995.13.3.553. [DOI] [PubMed] [Google Scholar]
- 12.Talpaz M, Kantarjian H, Liang J, Calvert L, Hamer J, et al. Blood. 1995;85:3257–3263. [PubMed] [Google Scholar]
- 13.Carella A M, Pollicardo N, Pungolino E. Leuk Lymphoma. 1993;9:477–483. doi: 10.3109/10428199309145754. [DOI] [PubMed] [Google Scholar]
- 14.Brenner M, Rill D, Moen R, Krance R A, Mino J, Jr, Anderson W F, Zhle J N. Lancet. 1993;342:1134–1137. doi: 10.1016/0140-6736(93)92122-a. [DOI] [PubMed] [Google Scholar]
- 15.Deisseroth A B, Zu Z, Claxton D, Hanania E G, Fu S, et al. Blood. 1995;83:3068–3076. [PubMed] [Google Scholar]
- 16.Long A A, Komminoth P, Lee E, Wolfe H J. Histochemistry. 1993;99:151–162. doi: 10.1007/BF00571876. [DOI] [PubMed] [Google Scholar]
- 17.Hanania E G, Deisseroth A B. Cancer Gene Ther. 1994;1:21–25. [PubMed] [Google Scholar]
- 18.Hanania E G, Fu S, Zu Z, Hegewisch-Becker S, Korbling M, Hester J, Durett A, Andreeff M, Mechetner E, Holzmayer T, Roninson I, Giles R E, Berenson R, Heimfeld S, Deisseroth A B. Gene Ther. 1995;2:285–294. [PubMed] [Google Scholar]