

Abstract
To determine the mechanism of action responsible for the in vivo antitumor activity of a phosphorothioate antisense inhibitor targeted against human C-raf kinase (ISIS 5132, also known as CGP69846A), a series of mismatched phosphorothioate analogs of ISIS 5132 or CGP69846A were synthesized and characterized with respect to hybridization affinity, inhibitory effects on C-raf gene expression in vitro, and antitumor activity in vivo. Incorporation of a single mismatch into the sequence of ISIS 5132 or CGP69846A resulted in reduced hybridization affinity toward C-raf RNA sequences and reduced inhibitory activity against C-raf expression in vitro and tumor growth in vivo. Moreover, incorporation of additional mismatches resulted in further loss of in vitro and in vivo activity in a manner that correlated well with a hybridization-based (i.e., antisense) mechanism of action. These results provide important experimental evidence supporting an antisense mechanism of action underlying the in vivo antitumor activity displayed by ISIS 5132 or CGP69846A.
Unequivocal proof of the in vivo mechanism of action for any drug has always been one of the greatest challenges facing the molecular pharmacologist. This challenge is mainly due to the difficulty in obtaining sound experimental evidence for drug mechanisms in complex animal models or in humans. Indeed, the mechanisms of action for many drugs that are used routinely in the clinic today with significant therapeutic benefit are poorly characterized. Unfortunately, without a clear understanding of a particular drug’s mechanism in vivo, improvement in the drug’s pharmacological properties and advances in the discovery of new drug classes based on novel mechanisms of action will be greatly impeded.
Antisense oligonucleotides represent a new paradigm for drug discovery that holds great promise for the delivery of potent and specific drugs with fewer undesired side effects (reviewed in refs. 1 and 2). This paradigm offers the opportunity to rapidly identify lead compounds based upon knowledge of the biology of a disease process. With this knowledge, the practitioner of antisense drug discovery can rapidly design, synthesize, and test a series of compounds in animal models and, because of the rational manner by which these compounds are designed to act, can experimentally address their mechanism of action in vivo with a relatively high degree of confidence. The most compelling evidence to support antisense mechanisms in living systems are (i) specific reduction in target gene expression and (ii) gradual reduction in drug potency as a result of incorporating increasing numbers of nonhybridizing bases (mismatches) into the oligonucleotide (i.e., rank order potency based on hybridization affinity). However, despite the fact that oligonucleotides have been shown to exert potent pharmacological activity in a variety of animal models, rigorous analyses providing strong experimental evidence to support an antisense mechanism of action underlying in vivo pharmacological activity have generally been lacking (2).
In this study, experimental evidence is provided supporting a mechanism of action based on Watson–Crick (3) base pair hybridization (i.e., antisense) underlying the in vivo antitumor activity of a phosphorothioate oligodeoxyribonucleotide (ODN) targeted against human C-raf kinase (C-raf). Previously we have demonstrated that this ODN, ISIS 5132 (also referred to as CGP69846A), is a potent inhibitor of tumor growth in vivo, and its administration to tumor-bearing mice results in reduced C-raf mRNA levels in tumors (4). We now address the sequence specificity of ISIS 5132 by comparing the effects of a series of mismatched analogs of this ODN on C-raf expression in vitro and tumor growth in vivo. Our results demonstrate a clear rank order potency that correlates with hybridization affinity for the pharmacological effects observed for ISIS 5132 and its mismatched analogs both in vitro and in vivo.
MATERIALS AND METHODS
Oligonucleotide Synthesis.
All oligonucleotides used in these studies were phosphorothioate ODNs. Synthesis and purification of phosphorothioate ODNs for tissue culture experiments and animal studies were performed as described (5–7). Briefly, ODNs were synthesized on an automated DNA synthesizer (MilliGen/Biosearch model 8800 DNA synthesizer) using modified phosphoramidate chemistry. For the stepwise thiation of phosphite linkages, the oxidation step was in 0.2 M [3H]1,2-benzodithiole-3-1,1,1-dioxide in acetonitrile. After cleavage from the controlled pore glass and deblocking in concentrated ammonium hydroxide at 55°C (18 h), the ODNs were purified by reverse-phase high-performance liquid chromatography in methanol/water/NaOAc. Only ODNs judged to be >90% full-length material by capillary gel electrophoresis were used. The 20-mer RNA complement to ISIS 5132, AAUGCAUGUCACAGGCGGGA, was synthesized as described previously (7).
Melting Curves.
Absorbance versus temperature curves were measured at 260 nm using a Gilford Response II spectrophometer. The buffer contained 100 mM Na+, 10 mM phosphate, and 0.1 mM EDTA (pH 7). ODN concentration was 4 μM for each strand, which was determined from the absorbance at 85°C based on extinction coefficients calculated according to Puglisi and Tinoco (8). Thermal melting temperature (Tm) values, free energies of duplex formation, and dissociation constants were obtained from fits of data to a two-state model with linear sloping base lines (9). Reported parameters are averages of three experiments.
Oligonucleotide Treatment in Cell Culture.
A549 lung carcinoma cells (10) growing on 10-cm plates at a density of 50–75% confluency were treated with ODNs in the presence of cationic lipid (Lipofectin Reagent, GIBCO/BRL) as described previously (4).
Northern Blot Analysis.
For determination of mRNA levels by Northern blot analysis, total RNA was prepared from cells by the guanidinium isothiocyanate procedure (11) 24 h after initiation of ODN treatment. Total RNA was isolated by centrifugation of the cell lysates over a cesium chloride cushion (4, 11). Northern blot analysis was as described previously (11). RNA was quantitated and normalized to G3PDH mRNA levels using a Molecular Dynamics PhosphorImager as described (12).
Oligonucleotide Treatment in Tumor Models.
Female BALB/c nude mice were obtained from Bomholtgard Breeding and Research Center (Ry, Denmark) and were used when 10–12 weeks old. A549 tumor fragments were implanted subcutaneously, and then serially passaged by a minimum of three consecutive transplantations before the start of treatment. Tumor fragments (≈25 mg) were implanted subcutaneously into the left flank of the animals with a 13-gauge trocar needle under Forene anesthesia (Abbott). ODN treatments were initiated when the tumors reached a mean tumor volume of 100 mm3. ODNs (formulated in saline solution) were administered intravenously daily by bolus infusion into the tail vein at the indicated doses. Tumor volumes were calculated as described (13). Each experimental condition included at least five animals per group. Average tumor volumes and standard deviations were calculated for each group and plotted.
RESULTS
We have previously identified a phosphorothioate ODN “antisense-designed” inhibitor of human C-raf kinase (C-raf) that specifically inhibits the expression of C-raf mRNA and protein in human cell lines in vitro (4). This ODN, ISIS 5132 (also referred to as CGP69846A), designed to hybridize with 3′-untranslated sequences within human C-raf mRNA, has also been shown to inhibit the growth of a variety of tumor types in vivo using nude mouse tumor xenografts and to inhibit C-raf mRNA expression in tumors in vivo (4). The sequence of ISIS 5132 is shown in Table 1.
Table 1.
Design, binding affinities, and activities of matched and mismatched phosphorothioate antisense ODNs targeted against human C-raf kinase mRNA
| ISIS # | Sequence | Tm, °C | Kd, nM at 37°C | Inhibition of C-raf mRNA levels in vitro (IC50, nM) | % inhibition of tumor growth in vivo |
|---|---|---|---|---|---|
| 5132* | TCCCGCCTGTGACATGCATT | 59.5 | 0.27 | 100 | 80.6 |
| 11691 | TCCCGCCTGCGACATGCATT | 52.5 | 4.7 | 225 | 68.9 |
| 11689 | TCCCGCCTGCTACATGCATT | 45.9 | 72 | 500 | 54.4 |
| 11688 | TCCCGCCTACTACATGCATT | 40.2 | 730 | 1000 | 21.4 |
| 11687 | TCCCGCCTACTTCATGCATT | 35.3 | 3000 | >1000 | 9.71 |
| 11686 | TCCCGCCCACTTCATGCATT | 30.5 | >5000 | >1000 | 0.00 |
| 11685 | TCCCGCCCACTTGATGCATT | 27.2 | >5000 | >1000 | ND |
| 10353 | TCCCGCGCACTTGATGCATT | >1000 | 0.00† |
Tm and Kd were determined for the indicated phosphorothioate ODNs targeted to complementary 20-mer RNA. Underlined bases indicate mismatches within the ISIS 5132 antisense ODN sequence, which targets RNA sequences within the 3′-untranslated region of human C-raf. Kd values for binding of oligonucleotides were calculated using G370 values derived from analyses of melting curves. Inhibition of C-raf mRNA levels in vitro was determined by Northern blot analysis 24 h after ODN treatment, and IC50 values calculated from dose-response curves (results representative of three independent experiments). Percent inhibition of tumor growth in vivo was determined at day 35 (see Fig. 3) by comparing the average tumor volume in animals receiving ODNs with tumor volumes in animals not receiving ODN (saline control). ND, not determined. Evaluation of the antitumor activity of ISIS 10353 was described previously (ref. 4). See text for methods.
Also known as CGP69846A.
From ref. 4.
To further address the mechanism of action of ISIS 5132, a series of phosphorothioate analogs were synthesized containing between one and seven mismatches to the ISIS 5132 binding site (Table 1). Melting temperatures (Tm) along with corresponding dissociation constants (Kd) were determined for the duplexes of each of these ODNs with a 20-mer oligoribonucleotide (RNA) complementary to ISIS 5132 (Fig. 1). As expected for Watson–Crick-based nucleic-acid hybridization (3), affinity decreased as the number of mismatches contained within the ISIS 5132 sequence increased (Table 1). No cooperative binding was observed for ODNs containing more than six mismatches.
Figure 1.
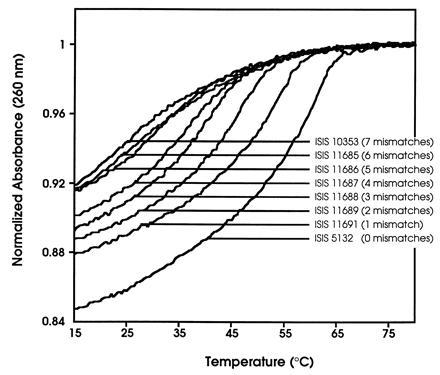
Hybridization of phosphorothioate ODNs to a synthetic 20-mer oligoribonucleotide (RNA) corresponding to the ISIS 5132 target sequence within the 3′-untranslated region of human C-raf kinase mRNA. Indicated ODNs are described in Table 1. Absorbance normalized at 95°C versus temperature for an equimolar mixture of antisense ODN and RNA complement. Tm values calculated from these curves are shown in Table 1.
To examine the specificity of ISIS 5132-mediated effects on C-raf gene expression in vitro, A549 cells were treated with either ISIS 5132 or mismatched ODNs over a range of concentrations and C-raf mRNA levels were examined (Fig. 2). ISIS 5132 treatment resulted in dose-dependent reduction in C-raf mRNA levels, displaying an IC50 for this effect of ≈100 nM (Table 1). None of the mismatched ODNs were as potent as ISIS 5132 in inhibiting C-raf mRNA expression. Furthermore, inhibition of C-raf mRNA levels gradually diminished as the number of mismatches within the ISIS 5132 was increased. No activity was observed for ODNs containing more than four mismatches. These findings are the predicted results if the effects of ISIS 5132 on C-raf mRNA expression were occurring through a mechanism based on Watson–Crick hybridization to cellular C-raf RNA.
Figure 2.
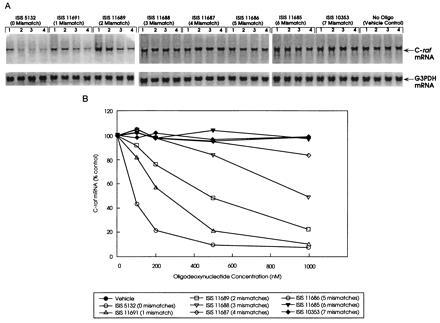
Reduction in C-raf kinase mRNA levels in A549 lung carcinoma cells after treatment with ISIS 5132 and mismatched phosphorothioate antisense ODNs. A549 cells grown in culture were treated with the indicated antisense ODNs (see Table 1) at increasing concentrations (100–1000 nM), and total RNA was prepared 24 h later and analyzed for C-raf and G3PDH mRNA levels by Northern blot analysis. (A) Northern blot analysis of C-raf mRNA levels. ODN treatments are indicated. Concentrations used were as follows. Lanes: 1, 100 nM; 2, 200 nM; 3, 500 nM; 4, 1000 nM. See Table 1 for ODN description. (B) Quantitation of C-raf mRNA levels shown in A after normalization to G3PDH mRNA levels. Quantitation was performed by PhosphorImager analysis as described. Results are representative of three independent experiments.
ISIS 5132 and it’s mismatched ODN analogs were also examined for antitumor activity in vivo against human A549 tumor xenografts. Fig. 3 shows the effects of ISIS 5132 and mismatched ODNs containing between one and five mismatches on the growth of A549 tumors in mice over time. ODNs were administered to tumor-bearing animals daily at a dose of 0.60 mg/kg. As reported previously (4), ISIS 5132 is a potent inhibitor of tumor growth in this model. Daily administration of this ODN reduced tumor volume by ≈80% at day 35 after tumor implantation (Table 1). Similar to the effects observed for the mismatched ODNs on C-raf mRNA expression in vitro, antitumor activity of ISIS 5132 was gradually diminished as increasing numbers of mismatches were incorporated into the ODN (Fig. 3 and Table 1). Even the incorporation of a single mismatch resulted in a significant loss of antitumor activity (80.6% versus 68.9% at day 35). No antitumor activity was observed for ODNs containing more than two mismatches.
Figure 3.
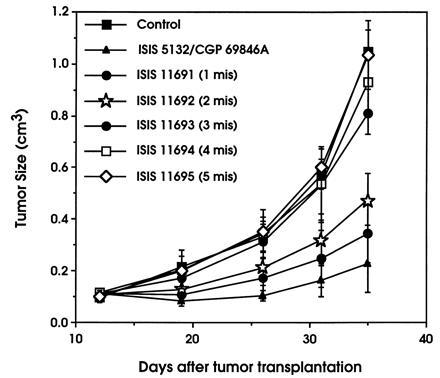
Effects of ISIS 5132 and mismatched phosphorothioate ODNs on the growth of A549 tumors in nude mice. A549 tumors were established subcutaneously in nude mice over a 12-day period as described. Following establishment of tumors, indicated ODNs (see Table 1) prepared in saline solution were administered once daily by bolus intravenous injection at a dose of 0.60 mg/kg, and tumor size was determined and tumor volume was calculated over a 3-week period following initiation of ODN treatment. The number of mismatches for each ODN is indicated using the abbreviation “mis” for mismatch. Each point represents the mean tumor volume in experimental groups containing eight animals per group. Standard deviations per group at each time point are indicated.
DISCUSSION
Phosphorothioate ODNs have been reported to exert biological effects through non-antisense mechanisms under certain experimental conditions (reviewed in refs. 2 and 14–16). Such studies have included both in vitro assays and, to a lesser extent, in vivo animal models. Furthermore, it has been suggested that the antitumor activity exerted by some phosphorothioate ODNs may be due to either a non-sequence-related effect of ODNs on binding to specific growth factors or a sequence-specific effect of ODNs on stimulation of natural killer cell activity in host animals (16–20). These reports, as well as others (2, 14, 15), demonstrate the need to rigorously examine the mechanism of action of a particular ODN before concluding that the pharmacological effects exerted by that ODN are derived from an antisense mechanism of action.
Previously, we reported the identification of a phosphorothioate antisense ODN (ISIS 5132 or CGP69846A) targeted against human C-raf kinase that inhibits C-raf mRNA and protein expression in vitro in a highly specific manner (4). We also demonstrated potent in vivo antitumor activity for ISIS 5132 that correlated with reduced C-raf mRNA levels in tumors. Furthermore, ISIS 5132 was reported to be completely devoid of natural killer stimulatory activity (4). Although these studies provide considerable experimental evidence for an antisense mechanism of action underlying the in vivo antitumor properties of ISIS 5132, additional studies addressing this issue in greater detail were clearly warranted.
Possibly the most convincing experimental evidence to support an antisense mechanism of action in vitro or in vivo is rank order potency based on hybridization affinity. Although demonstrating reduced levels of target mRNA or protein after ODN administration is important evidence supporting antisense mechanisms of action, it is clear that a variety of factors including altered growth conditions can have marked effects on gene expression. Thus, it is generally difficult to conclude whether reduced tumor growth is the consequence or the cause of reduced target gene expression.
In this study, we report on the rank order potency of mismatched derivatives of ISIS 5132 using both in vitro and in vivo assays. We demonstrate that the presence of a single mismatch in the ISIS 5132 sequence results in a significantly less potent inhibitor of C-raf mRNA expression in vitro and of tumor growth in vivo. Moreover, incorporation of additional mismatches results in further loss of in vitro and in vivo activity in a manner that correlates well with a Watson–Crick (3) hybridization-based (i.e., antisense) mechanism of action. These results, along with the results described in our previous studies (4), provide strong experimental evidence for an antisense mechanism underlying the in vivo antitumor activity displayed by ISIS 5132. More generally, these results demonstrate the utility of antisense technology for the rational design of pharmacological agents for therapeutic intent.
Footnotes
Abbreviations: ODN, oligodeoxynucleotide; Tm, thermal melting temperature.
References
- 1.Milligan J F, Matteucci M D, Martin J C. J Med Chem. 1993;36:1923–1937. doi: 10.1021/jm00066a001. [DOI] [PubMed] [Google Scholar]
- 2.Crooke S T, Bennett C F. Annu Rev Pharmacol Toxicol. 1996;36:107–129. doi: 10.1146/annurev.pa.36.040196.000543. [DOI] [PubMed] [Google Scholar]
- 3.Watson J D, Crick F H C. Nature (London) 1953;171:737–739. doi: 10.1038/171737a0. [DOI] [PubMed] [Google Scholar]
- 4.Monia B P, Johnston J F, Geiger T, Muller M, Fabbro D. Nat Med. 1996;2:668–675. doi: 10.1038/nm0696-668. [DOI] [PubMed] [Google Scholar]
- 5.Monia B P, Lesnik E A, Gonzalez C, Lima W F, McGee D, Guinosso C J, Kawasaki A M, Cook P D, Freier S M. J Biol Chem. 1993;268:14514–13522. [PubMed] [Google Scholar]
- 6.Chiang M-Y, Chan H, Zounes M A, Freier S M, Lima W F, Bennett C F. J Biol Chem. 1991;266:18162–18177. [PubMed] [Google Scholar]
- 7.Bennett, C. F., Kornbrust, D., Henry, S., Dutson, S., Hall, W. & Jacoby, H. I. J. Pharmacol. Exp. Ther., in press. [PubMed]
- 8.Puglisi J D, Tinoco I., Jr Methods Enzymol. 1989;180:304–325. doi: 10.1016/0076-6879(89)80108-9. [DOI] [PubMed] [Google Scholar]
- 9.Petersheim M, Turner D H. Biochemistry. 1983;31:12055–12061. [Google Scholar]
- 10.Valenzuela D M, Groffen J. Nucleic Acids Res. 1985;14:844–852. doi: 10.1093/nar/14.2.843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ausubel F M, Brent R, Kingston R E, Moore D D, Seidman J G, Smith J A, Struhl K, editors. Current Protocols in Molecular Biology. New York: Greene & Wiley; 1987. [Google Scholar]
- 12.Dean N M, McKay R. Proc Natl Acad Sci USA. 1994;91:11762–11766. doi: 10.1073/pnas.91.24.11762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Meyer T, Regenass U, Fabbro D, Alteri E, Rosel J, Muller M, Caravatti G. Int J Cancer. 1989;43:851–856. doi: 10.1002/ijc.2910430519. [DOI] [PubMed] [Google Scholar]
- 14.Stein C A, Cheng Y-C. Science. 1993;261:1004. doi: 10.1126/science.8351515. [DOI] [PubMed] [Google Scholar]
- 15.Krieg A M, Stein C A. Antisense Res Dev. 1995;5:241. doi: 10.1089/ard.1995.5.241. [DOI] [PubMed] [Google Scholar]
- 16.Stein C A. Nat Med. 1996;1:1119–1121. doi: 10.1038/nm1195-1119. [DOI] [PubMed] [Google Scholar]
- 17.Yamamoto S, Yamamoto T, Shimada S, Kuramoto E, Yano O, Kataoka T, Tokunaga T. Microbiol Immunol. 1994;36:983–997. doi: 10.1111/j.1348-0421.1992.tb02102.x. [DOI] [PubMed] [Google Scholar]
- 18.Kuramoto E, Yano O, Kimura Y, Baba M, Makino T, Yamamoto S, Yamamoto T, Kataoka T, Tokunaga T. Jpn J Cancer Res. 1992;83:1128–1131. doi: 10.1111/j.1349-7006.1992.tb02734.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Yamamoto T, Yamamoto S, Kataoka T, Tokunaga T. Microbiol Immunol. 1994;38:831–836. doi: 10.1111/j.1348-0421.1994.tb01867.x. [DOI] [PubMed] [Google Scholar]
- 20.Guvakova M A, Yakubov L A, Vlodavsky I, Tonkinson J L, Stein C A. J Biol Chem. 1994;270:2620–2627. doi: 10.1074/jbc.270.6.2620. [DOI] [PubMed] [Google Scholar]