

Abstract
The transcription of CAB genes, encoding the chlorophyll a/b-binding proteins, is rapidly induced in dark-grown Arabidopsis seedlings following a light pulse. The transient induction is followed by several cycles of a circadian rhythm. Seedlings transferred to continuous light are known to exhibit a robust circadian rhythm of CAB expression. The precise waveform of CAB expression in light–dark cycles, however, reflects a regulatory network that integrates information from photoreceptors, from the circadian clock and possibly from a developmental program. We have used the luciferase reporter system to investigate CAB expression with high time resolution. We demonstrate that CAB expression in light-grown plants exhibits a transient induction following light onset, similar to the response in dark-grown seedlings. The circadian rhythm modulates the magnitude and the kinetics of the response to light, such that the CAB promoter is not light responsive during the subjective night. A signaling pathway from the circadian oscillator must therefore antagonize the phototransduction pathways controlling the CAB promoter. We have further demonstrated that the phase of maximal CAB expression is delayed in light–dark cycles with long photoperiods, due to the entrainment of the circadian oscillator. Under short photoperiods, this pattern of entrainment ensures that dawn coincides with a phase of high light responsiveness, whereas under long photoperiods, the light response at dawn is reduced.
Keywords: firefly luciferase, luminescence imaging, circadian rhythm, gene expression
Plants combine endogenous timing information with light signals received from the environment to regulate development and metabolism (1). The genes encoding chlorophyll a/b-binding proteins (CAB genes) provide molecular markers for these regulatory pathways. CAB transcription is controlled by the circadian clock and also by light, transduced principally via the phytochrome family of red light photoreceptors (reviewed in ref. 2). CAB expression levels are very low in dark-grown seedlings. If dark-grown seedlings are briefly exposed to red light and returned to darkness, strong CAB expression is induced, and levels reach a peak within approximately 4 h of the light signal in bean or tobacco (3–6), or approximately 12 h in wheat (7). Expression levels then exhibit a circadian rhythm for several days (3, 7) before returning to the level of dark controls (2). This complex response comprises three types of regulation on different time scales: (i) the initial response within a few hours, (ii) the circadian rhythm with a period of approximately 24 h, and (iii) the increase in mean expression level, which persists for several days.
The mechanisms responsible for these patterns have been investigated at several levels. Differential involvement of the multiple phytochrome gene products has been demonstrated (5, 8), and the required CAB promoter sequences have been identified (9, 10). Although the mechanism of circadian regulation is unclear, the “gating hypothesis” (11) proposes that the circadian clock acts negatively on CAB expression, antagonizing the positive effects of phototransduction pathways during the subjective night.
The transient induction of CAB expression immediately after plants are exposed to light is referred to as the “acute” response to light (9, 11). Many circadian-regulated processes exhibit acute effects of light: the expression of clock-controlled genes (ccg) in Neurospora is induced by light (12), for example, and the production of melatonin in cultured avian pinealocytes is suppressed (13). Current evidence indicates that the circadian oscillator is not required for these acute responses: ccg remain inducible by light in a fungal clock mutant, frequency, that lacks a functional circadian oscillator (12). An intact oscillator can nevertheless modulate the acute responses. Stomatal aperture, for example, exhibits both an acute response to light and a circadian rhythm in many plant species. The rate and extent of the light-induced stomatal opening are modulated by a circadian rhythm: maximal light responsiveness coincides with the maximum of the rhythm in “resting” stomatal aperture (14). At the level of gene expression, the light induction of immediate-early genes (such as c-fos) in the mammalian suprachiasmatic nucleus is restricted to the subjective night by a circadian rhythm (for a review, see ref. 15).
The circadian oscillator is itself strongly modulated by light signals, which keep the oscillator to a 24-h period in natural conditions, and in phase with the local day-night cycle (1, 16). The period of CAB expression in Arabidopsis, for example, is approximately 24.5 h in constant light (LL) but lengthens to 30 h or more in constant darkness (DD) (17). The light–dark cycle (LD) must shift the phase of the oscillator by approximately 0.5 h each day to maintain a 24-h period, with the peak of CAB expression consistently in the early part of the day. This process is known as entrainment, and the phototransduction pathways involved are examples of the “input” pathways to the circadian clock (16).
To confer a selective advantage upon the organism, entrainment must be adaptable. Light signals, especially photoperiod, change with the seasons in temperate latitudes: the optimal phase for a rhythmic process may vary in parallel (18). The onset of activity in several nocturnal rodents, for example, occurs close to dusk under all photoperiods. Under short photoperiods, the phase of the activity onset is several hours after dusk; it occurs much closer to dusk under long photoperiods, however, and may even precede lights-off (18). This is in contrast to several rhythms in higher plants, in which the phase of entrainment is at a constant time after lights-on (19) or lights-off (ref. 20; for review, see ref. 21). Other plant rhythms exhibit two phases: one in long and the other in short photoperiods (22, 23). Entrainment thus ensures that circadian-regulated processes occur at appropriate times under all photoperiods, but not always at the same phase relative to dawn or dusk.
CAB expression in LD-grown plants is clearly regulated by an entrained circadian rhythm (2, 11). However, it is not clear which of the other regulatory components affect the expression of CAB in LD. The noninvasive luciferase (luc) reporter system has provided a powerful tool to dissect this regulatory network. Bioluminescence from transgenic plants containing a cab::luc fusion recapitulates the regulation of CAB transcription (17). The low-light imaging assay allows us to reveal temporal patterns of gene expression in unprecedented detail. We now use this marker to demonstrate that CAB transcription in LD-grown plants is indeed subject to the three regulatory components defined in dark-grown tissues: (i) the acute induction in response to light, (ii) the circadian rhythm, and (iii) the long-term control of mean expression level. In addition, the circadian clock dramatically modulates the acute response of CAB expression to light. We determine the phase of entrainment of the CAB rhythm under different photoperiods, showing that both lights-on and lights-off signals contribute to entrainment. After testing each component separately, we examine their combined effects on the waveform of CAB expression under various photoperiods. The observed changes in the pattern of CAB expression under various photoperiods are explained by the integration of endogenous information from the circadian clock with light signals from the environment.
MATERIALS AND METHODS
Plant Materials and Growth Conditions.
Transgenic Arabidopsis thaliana lines containing a cab::Ω::luc fusion in the C24 and Columbia gl1 backgrounds have been described (17, 24). Dark-grown seedlings were kept in a temperature-controlled dark room for 7 days in sterile culture on solid Murashige and Skoog (MS) medium containing sucrose (24). All other plants were germinated on the same medium in a 12-h light/12-h dark cycle (LD12:12) for 5 days before any other light treatments. Light sources for red light (25 μmol·m−2·s−1) and white light (50 μmol·m−2·s−1) treatments and other growth conditions were as described previously (17). The fluence-response of CAB induction was tested at 29 h in DD (see Fig. 2A); the standard light treatment was unlikely to be saturating, as 30 min of 270 μmol·m−2·s−1 light gave more than 2.5-fold higher acute inductions (unpublished data). Phase determinations (see Fig. 3) performed on replicate samples that were transferred to constant conditions after 5 or 7 days of entrainment gave similar results, indicating that the plants were stably entrained (unpublished data).
Figure 2.
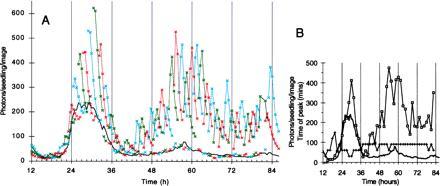
Circadian modulation of the acute response to light. Light-grown plants were transferred to DD at the normal lights-off time of the preceding LD12:12 (12 h). Replicate samples (a total of 47) received 30 min light every 1.5 h (marked by ticks above the time axis). Eight images were collected for each sample at 30-min intervals, two before and six after the light treatment. A dark control sample was imaged every 30 min, and is included as a reference in each graph (solid line). Data are representative of three experiments. Data points were lost at 80.5 and 81 h. (A) Data from successive samples are graphed in red, blue, and green for clarity. A gap in each trace marks the time of the light pulse. (B) The peak amplitude of each acute response after subtraction of the dark control level (squares), and the kinetics of induction measured as the time (min) between the light pulse and the image that gave the peak luminescence (crosses).
Figure 3.
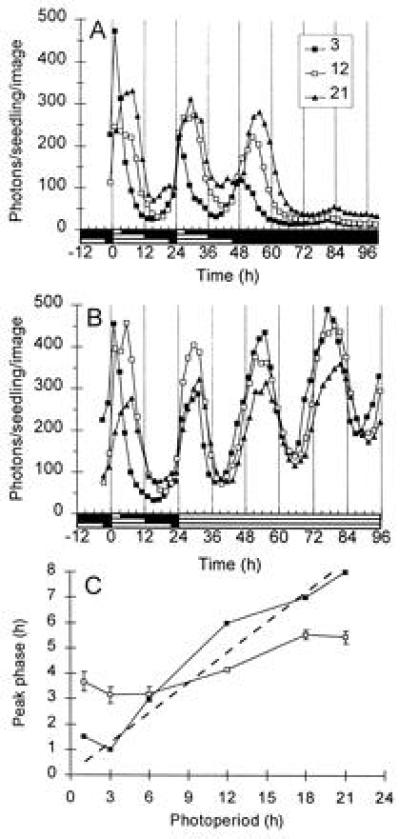
Entrainment of CAB under LD cycles. Luminescence of light-grown plants entrained to LD cycles with three photoperiods (indicated in the legend) and transferred into DD (A) and LL (B). (C) Estimated phases of peak luminescence in hours after the projected dawn of the preceding LD cycle, in plants entrained to various photoperiods and transferred to DD, as in A (solid symbols, phase of the first peak) and LL, as in B (open symbols, phase of a modified cosine fit). The dashed line marks the first 40% of the photoperiod. Data are representative of nine samples for each photoperiod, in two experiments; LL data in C are means ± SEM. Open box on time axis, light period; solid box, dark period.
Bioluminescence Assays.
Bioluminescence imaging conditions were as described previously (17, 24), except that dark-grown material and the dark-adapted plants shown in Fig. 2A were handled under infrared safelight. Video images were analyzed as described previously (24), except that the raw data were corrected for variations in camera sensitivity over the field of view, before camera noise subtraction and normalization for seedling number. Phase estimates for rhythms in LL were derived from the least-squares fitting procedure described (17), except that the function used was a modified cosine rather than a sine. Samples released into LL had periods of approximately 24.5 h, and the period estimates from each experiment fell within a range of 0.7 h. The phases of peak expression in DD were estimated manually. The rapid damping of expression levels in these conditions makes the least-squares fitting unreliable and impedes manual scoring of the second peak.
RESULTS
Acute Responses in Both Dark- and LD-Grown Tissue.
A brief red light treatment induces multiple peaks of CAB expression in dark-grown seedlings of several species (3–6), allowing us to distinguish the initial, acute response to light clearly from circadian-regulated peaks of CAB expression. Fig. 1A confirms this result in cab::luc Arabidopsis. Dark-grown seedlings luminesced very weakly, at or below the detection limit of the imaging system, so no circadian components were clearly distinguishable. A 2-min red light treatment induced a peak of luminescence in the image 1.5 h after the light pulse, which fell to a trough by 4 h after the pulse. Two peaks were detected thereafter, at 22 and 56 h, suggesting that the light pulse initiated or revealed a rhythm with a long period (34 h), as in cab::luc tobacco (6).
Figure 1.
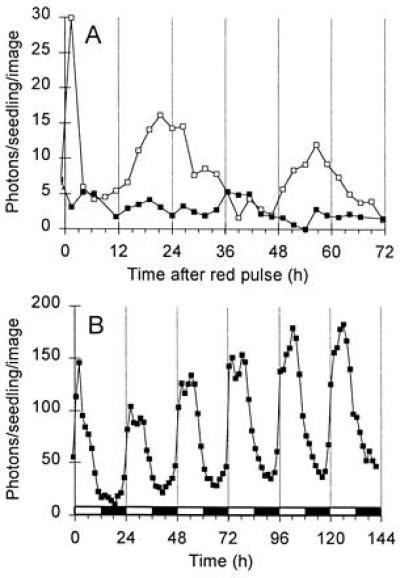
The acute response to light in dark-grown and light-grown seedlings. Video images integrating luminescence for 25 min were collected at the times indicated, and were analyzed to calculate luminescence levels, as detected photons per seedling per image. (A) Dark-grown cab::luc Arabidopsis exposed to 2 min red light at time 0 (open symbols) or maintained in dark (filled symbols). Data are representative of three experiments. (B) cab::luc Arabidopsis grown and imaged in LD12:12. Open box on time axis, light period; solid box, dark period.
Any acute response that occurs after dawn in light-grown plants may be largely obscured by circadian-regulated CAB expression. Rhythmic expression levels rise rapidly in anticipation of dawn, and peak 4–6 h after dawn in plants grown under our standard LD12:12 photoperiod. Fig. 1B shows that images taken at 1.5-h intervals can reveal bimodal peaks in LD12:12, suggesting that an acute induction by light preceded the circadian peak of CAB expression. This presumptive acute response was most prominent on the first day of the time course, but was reduced to a shoulder of the circadian peak by the fifth day, and was barely detectable on the sixth day, suggesting that the acute response is developmentally regulated (see Discussion).
Circadian Gating of the Acute Response.
To examine acute responses to light at times other than the lights-on point of an LD cycle, we applied 30-min white light pulses to seedlings that were grown in LD12:12 and transferred to DD. Fig. 2A shows the patterns of luminescence following a light treatment at various times in DD. Dark control plants showed the reduced expression levels and long-period rhythm typical of DD (17). Luminescence levels before the light treatments were very similar to dark controls. Light induced a transient increase in luminescence, similar to the acute response in dark-grown tissue, but the response varied greatly over the time course. Fig. 2B shows the peak amplitude and the kinetics of induction. The strongest and most rapid (Fig. 2B) inductions coincided with the luminescence peaks in the dark controls (approximately 30, 58, and 82 h). At 30 h, induction reaches more than 400 photons per seedling per image above the dark control in the first image, 30 min after the light pulse began. Responses were small to negligible, and peaked in or after the image 90 min after light, at 13.5–20 h and 36–40 h. Induction amplitude rose after 40 h and remained high thereafter (Fig. 2B). Samples in each of three replicate experiments reached peak luminescence before the 90-min image during the second peak of luminescence in the dark control, though Fig. 2B indicates that this was not always consistent throughout the second peak. The mean time to peak luminescence for light pulses in the interval from 55.5 to 61.5 h was 79 min (SEM = 3, n = 14). This is significantly shorter than the induction times for equivalent intervals clearly before and after the second peak (92 min ± 3, n = 13 for each of 47–53 h and 67–73 h). These data indicate that plants grown in LD retain an acute response to light, which is modulated by the circadian clock. The waveform of CAB expression in LD must therefore reflect both a circadian rhythm and an acute response to dawn.
Entrainment of the CAB Rhythm by LD Cycles.
The dark-to-light and light-to-dark transitions at dawn and dusk are important cues in entraining the oscillator to natural day-night cycles. We monitored luminescence in cab::luc plants entrained under various LD cycles (from LD1:23 to LD21:3) to determine how the photoperiod affected the phase of CAB expression. Fig. 3 A and B show some of the patterns of luminescence, which were measured after transfer to DD (Fig. 3A) or LL (Fig. 3B), and Fig. 3C summarizes the phases of peak luminescence. The peak of expression occurred in the early or middle part of the light period in all but the shortest photoperiods (for comparison, the dashed line in Fig. 3C indicates when 40% of each photoperiod has elapsed). Long photoperiods lead to later phases in both DD and LL. The phase of the first peak in DD was strongly dependent on photoperiod, ranging from 1–2 h to 7–8 h after projected dawn (Fig. 3C). Peak phases in LL fell in a narrower range, (3.3 to 5.3 h after projected dawn) (Fig. 3C).
The phases in DD were scored at the first peak, whereas the phases in LL were estimated, more accurately, by cosinor analysis of several days of data (17). It is possible that the DD data reflect a transient phase shift, which is restricted to the first cycle after entrainment (1). Fig. 3A, however, shows that the phase of the second peak in DD is also more delayed following longer photoperiods, indicating that at least a portion of the DD phase shift is maintained beyond the first cycle. The severe damping of expression level and the lengthening of circadian period makes the estimation of peak position much less reliable for the second peak than for the first (see Materials and Methods). Nevertheless, data comprising nine samples for each photoperiod from two experiments (Fig. 3 A and C and unpublished data) indicated that the photoperiod-dependent delay of the second peak was significant (P = 0.02 in Student’s t test). The mean phases exhibited a range of 2.6 h (least to most delayed), though the associated standard errors are relatively large (0.4 to 0.8 h). These data indicate that CAB expression is regulated by an entrained, circadian oscillator in both LL and DD.
The Waveform of CAB Expression in LD Cycles.
We assayed CAB expression in a range of photoperiods to determine whether the interactions of light and circadian rhythms described above could account for the pattern of luminescence. Fig. 4A shows the patterns in short photoperiods; Fig. 4B shows the patterns in long photoperiods. The comparison of data from all the photoperiods reveals a strong trend in the mean luminescence level per seedling, which increased with photoperiod (only LD9:15 was higher than expected). This may in part be due to an effect of total fluence on plant growth (see Discussion). Peak luminescence levels progressively decreased over the time course in LD1:23 and LD3:21, whereas LD6:18 and the longer photoperiods maintained stable luminescence levels. This indicates that light-dependent processes independent of growth also contributed to maintaining CAB expression levels under photoperiods of 6 h or more. Such regulation is suggestive of a long-term control of expression level, similar to that described in etiolated seedlings (see Discussion).
Figure 4.
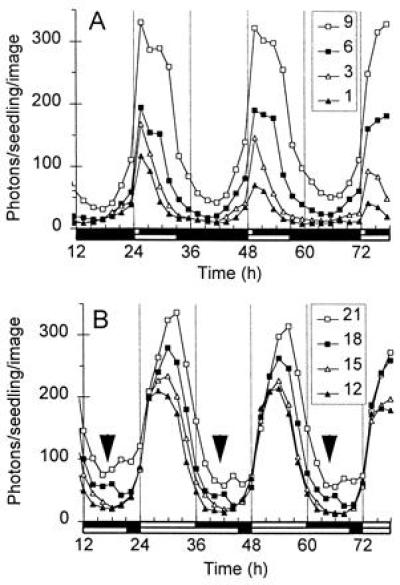
Waveform of CAB expression under LD cycles. Luminescence of light-grown plants entrained to LD cycles with various photoperiods (indicated in the legends). (A) LD1:23 to LD9:15; (B) LD12:12 to LD21:3. Data are representative of four experiments. Open box on time axis, light period; solid box, dark period. (A) LD1:23 and LD9:15; (B) LD12:12 and LD21:3. (B) Arrowheads indicate times of rising luminescence in LD18:6 but not LD12:12 or LD15:9 (see Discussion).
Under LD, circadian rhythms cannot always be distinguished from diurnal regulation in response to the light-to-dark transitions. Two features of the CAB waveform were indicative of circadian regulation. First, luminescence levels increased in anticipation of dawn and decreased before dusk under most photoperiods, indicating regulation by a timing mechanism. Second, the phase of peak luminescence depended on photoperiod, suggesting the photoperiod-dependent phase of entrainment (as in Fig. 3) that is unique to a circadian oscillator. The peak of luminescence moved to later phases under the long photoperiods, consistent with the pattern of entrainment observed above (Fig. 3).
The acute induction of luminescence at lights-on was also evident. The apparent amplitude of the induction varied strongly under the various photoperiods, however. This can be predicted because the amplitude of the acute response depends on the phase at which dawn occurs (Fig. 2), and the phase of entrainment depends on photoperiod (Fig. 3). The integration of these controls leads to the photoperiod-dependence of the acute response at dawn, although photoperiod-dependence of development (see below) could also contribute. LD1:23 and LD3:21 did not show distinct peaks from the acute response and the circadian rhythm (Fig. 4A), as expected if the early phase of the circadian peak leads to a complete overlap with the acute response at lights-on (Fig. 3C). The rate of change of luminescence increased rapidly at dawn, however, which is suggestive of an acute induction. The LD6:18 and LD9:15 traces alone suggested a distinct acute response immediately after dawn in up to 2 of the 3 days assayed, though the imaging interval used (2 h) did not reveal a bimodal peak. Luminescence in LD18:6 and LD21:3 did not show a clear acute response, though there was some enhancement in the rate of luminescence increase at lights-on. Dawn fell at early phases under these conditions, because the peak of CAB expression occurred about 1h later than in LD12:12 (Fig. 3C). The amplitude of the acute response was thus expected to be about 50% less than in LD12:12 (Fig. 2B). The amplitude of the circadian rhythm, in contrast, increased by about 60% concomitant with the rise in mean expression level. These factors combine to minimize the contribution of the acute response to the waveform of CAB expression in long photoperiods.
Fig. 4 A and B also suggest a developmental regulation of the acute response, in addition to its circadian modulation. The acute response is not clear in LD12:12, for example, in contrast to Fig. 1B. This is partly due to the lower time resolution of the data (2 h in Fig. 4B, 1.5 h in Fig. 1B). Samples in Fig. 4B had been entrained to the various photoperiods, however, and were therefore 3 days older than those of Fig. 1B, such that 12 h in Fig. 4B is equivalent to 84 h in Fig. 1B. The acute response at dawn was barely detectable in plants of that age in Fig. 1B, indicating that data from the two experiments are consistent. These data therefore suggest that CAB expression in LD is not only regulated by the three components revealed by a light pulse in etiolated seedlings. The pattern of CAB expression also reflects circadian and possibly developmental modulation of the acute response, photoperiod-dependent entrainment, and an effect of integrating these factors, namely the photoperiod-dependence of the acute response to lights-on.
DISCUSSION
CAB expression patterns in light-grown plants reflect endogenous timing information, light signals from the environment, and several interactions between these regulators. Our data indicate that three principal components control CAB expression in LD-grown plants: (i) the transient induction following the onset of light (Figs. 1 and 2), (ii) the circadian rhythm (Figs. 2 and 3), and (iii) a long-term control of mean expression level (Figs. 3 and 4). Two interactions between the regulators provide fine control: light signals entrain the circadian clock (Fig. 3), and the clock modulates the acute response to light (Fig. 2). The acute response may also exhibit a developmental regulation over several days.
LD-grown Arabidopsis exhibits an acute induction of CAB by light (Figs. 1B and 2) that bears many similarities to the response typically studied in dark-grown seedlings (Fig. 1A). The bimodal peaks of expression observed under LD cycles (Fig. 1B) are unlikely to represent two phase signals from the circadian clock, as bimodality is not observed under constant conditions (for example, see dark control of Fig. 2) and as induction follows light onset whenever light occurs (Fig. 2). The acute response of CAB to light in green tissue is gated by the circadian clock (Fig. 2). A similar effect was suggested by CAB RNA analysis in light-grown tomato, following 4-h light pulses given from 72 to 93 h (time relative to Fig. 2). Only the pulse at 72–76 h showed a significant induction of CAB RNA; this pulse overlaps with what appears to be a peak in CAB RNA accumulation in the unirradiated plants (25). These results contrast with the phase-independent inducibility of Cab1 RNA in dark-grown wheat (7). The luciferase system allows repeated measures from the same samples, removing variability, and provides far greater temporal resolution than was available in previous studies. Thus, our data quantify the precise circadian control of both the amplitude and the kinetics of the acute response.
The effect of the acute response on luminescence level is clearly not additive with the circadian rhythm (Fig. 2B), nor is it simply multiplicative (the ratio of induced to control luminescence is not constant, even on the first day in DD; data from Fig. 2B). The kinetics of the induction are also phase-dependent, as peak luminescence occurs within 30 min at the first peak of the dark control luminescence (30 h in Fig. 2A) but not until 90 min in the trough of the dark rhythm. The circadian clock must therefore interact with the phototransduction pathways regulating the CAB promoter. The interaction could occur at the CAB promoter, for example, by interactions of circadian-regulated transcription factors (10) with the CGF complex, which is required for the acute response in dark-grown tobacco (9). The pathways may alternatively interact in the cytoplasm: the concentration of cytoplasmic Ca2+, for example, is a candidate phototransduction intermediate that also exhibits circadian regulation (26). The reciprocal control model for phytochrome signaling (27) suggests a possible mechanism for such regulation. CAB gene expression is activated by a Ca2+/calmodulin-dependent branch of the phytochrome signal transduction pathway in this model. This activation is antagonized by a cGMP-dependent pathway, albeit at a step downstream of calmodulin. Circadian control of flux through the cGMP-dependent pathway might therefore result in circadian modulation of the terminal portion of the Ca2+/calmodulin-dependent pathway, and hence, in circadian control of the acute response of CAB expression to light.
Our results suggest that the acute response pathway mediates the short-term responsiveness of the CAB promoter to light, which results in rapid responses to lights-on and lights-off. This pathway may be continuously active in LL (perhaps modified by photoreceptor adaptation). The circadian rhythm in LL is therefore likely to reflect the rhythmic antagonism of the acute response by signals from the circadian clock, as suggested by the gating hypothesis (11).
The rhythm of acute responsiveness appears to be tightly coupled to the rhythm in “basal” CAB expression (Fig. 2A), suggesting that a single output signal transduction pathway from the oscillator may mediate both rhythms. Only one instance in our data suggests an uncoupling of basal expression from light responsiveness. A small increase in luminescence was consistently observed at the end of the light interval under 18 and 21 h photoperiods (indicated by arrows for LD18:6 in Fig. 4B; see Fig. 3 A and B for LD21:3), followed by a decrease immediately after the transition to darkness. This suggests that CAB expression was light responsive at this time. Plants under LD12:12 or LD15:9 were already in the dark interval at this time, and showed no increase in luminescence levels. Rather, luminescence was still falling to its trough, reinforcing the conclusion that the rising luminescence in LD18:6 samples was light-dependent and not due to the circadian rhythm alone. This suggests that the antagonism of the acute response by the circadian clock may be relieved before (and independently of) the circadian-regulated rise of CAB expression in darkness, in anticipation of lights-on.
Development may also modulate the acute response to light. The amplitude of the acute response at dawn in LD diminished as the plants aged over the several days of an experiment (Fig. 1B), which is suggestive of developmental regulation. Similar developmental control of photoreceptor pathways has been well documented, not only affecting the responsiveness of specific promoters to light induction (28, 29) but also determining which photoreceptor(s) regulate a specific promoter (30). It remains possible that acute induction could be reduced after a certain number of lights-on signals, independent of developmental stage. The high amplitude of the acute response after 40 h in DD (Fig. 2) likely reflects the accumulation of light-labile phytochrome (PhyA) (ref. 31 and references therein), which is known to contribute to the acute induction of CAB in dark-grown tissue (5).
Our results suggest that light-grown plants exhibit a long-term control of CAB expression levels. Our data relate to whole seedlings: their growth over the course of these experiments is probably responsible in part for the upward trend of luminescence in LL (Fig. 3B; ref. 17) and for increased luminescence in long photoperiods relative to shorter ones (Fig. 4). However, the peak expression level progressively decreases over the course of the experiment, in plants under short photoperiods (Fig. 4A). This suggests that a long-term control of expression level by phototransduction pathways must also contribute to maintaining expression levels under long photoperiods, independently of growth. Such long-term regulation likely depends on photoreceptor pools with stable activity, such as the stable forms of phytochrome (32). Measurements of plant development during the imaging series and the use of mutants deficient in specific photoreceptors should resolve these issues.
The phase of peak CAB expression in DD was later in plants exposed to long photoperiods in the preceding LD, relative to those from short photoperiods (Fig. 3 A and C). A narrower range of phases resulted after release into LL (Fig. 3 B and C). The linear correlation of phase in DD to photoperiod (except in LD1:23), with no constant phase relationship to either lights-on or lights-off, indicates that both transitions contribute to entrainment. This is in contrast to many other rhythms examined in higher plants (see Introduction) but is consistent with an earlier report in wheat seedlings, which was based on a single entraining light-to-dark transition (33). The phase differences in our data were not due solely to period lengthening in the plants entrained to short photoperiods (as in DD, see Fig. 2 and ref. 17), for this would give later phases following short photoperiods, rather than the earlier phases observed. A photoperiod-dependent phase difference was also observed for the second peak of expression in DD, though the rapid damping and period lengthening make phase estimation more difficult for this peak. Most circadian rhythms do not show such dramatic amplitude and period modifications in DD. The entrainment of CAB expression, however, formally supports our previous conclusion that even the unusual rhythm of CAB in DD is regulated by a circadian oscillator (17), not by a simple “hourglass.” Light input to the Arabidopsis circadian oscillator thus regulates phase as well as period (17), such that the peak of CAB expression occurs in the early part of the light period, under a wide range of photoperiods (Fig. 3C). Entrainment of this type may be one of the selective advantages conferred by a circadian clock (18), as opposed to an “hourglass” mechanism that times a fixed interval from dusk. Such an hourglass would give appropriate phases of CAB expression under only a limited range of photoperiods.
The waveform of CAB expression in LD cycles reflects regulation by light (in the acute response and the long-term maintenance of expression level), by the circadian clock, and by their interactions described above. The integration of entrainment and the circadian modulation of the acute response to light has an interesting consequence in short photoperiods. Dawn falls shortly before the peak of CAB expression in these conditions (Fig. 3C), due to the early phase of entrainment. The light induction of CAB therefore exhibits both high amplitude and rapid kinetics during the light interval of the LD cycle (Fig. 2). This combination of the circadian-regulated peak of CAB expression and the maximal responsiveness of CAB to ambient light may be selectively advantageous, concentrating resources on photosynthesis during the brief photoperiod. A mathematical model could now be derived to describe the waveform of CAB expression, and used to characterize mutations that affect the various components (either in the CAB promoter sequence or in upstream regulators). Multiple photoreceptors will be activated by our white light treatments, however, and models related to a single photoreceptor will be simpler and may be more useful in initially dissecting signaling mechanisms. The quantitative analysis permitted by the luminescent reporter system will be a critical tool in ultimately dissecting the network of regulatory interactions that operates under natural conditions.
Acknowledgments
We thank Dr. Gene Block and members of the Kay laboratory for useful discussions; Dr. M. Straume for biomathematical tools; and K. W. Smith, D. Gerber, K. Fisher, P. Tait, and A. Moran for help in imaging shifts. This work was supported by National Science Foundation Grant MCB-9316756 to S.A.K. A.J.M. was a Department of Energy Fellow of the Life Sciences Research Foundation. S.A.K. is supported by an award from the W. M. Keck Foundation.
Footnotes
Abbreviations: CAB, chlorophyll a/b-binding protein gene; LL, constant white light; DD, constant dark; LDx:y, a cycle of x-h white light and y-h dark.
References
- 1.Sweeney B M. Rhythmic Phenomena in Plants. San Diego: Academic; 1987. [Google Scholar]
- 2.Batschauer A, Gilmartin P M, Nagy F, Schafer E. In: Photomorphogenesis in Plants. Kendrick R E, Kronenberg G H M, editors. Vol. 2. Dordrecht, The Netherlands: Kluwer; 1994. pp. 558–600. [Google Scholar]
- 3.Tavladoraki P, Kloppstech K, Argyroudi-Akoyunoglou J. Plant Physiol. 1989;90:665–672. doi: 10.1104/pp.90.2.665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Wehmeyer B, Cashmore A R, Schafer E. Plant Physiol. 1990;93:990–997. doi: 10.1104/pp.93.3.990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Reed J W, Nagatani A, Elich T D, Fagan M, Chory J. Plant Physiol. 1994;104:1139–1149. doi: 10.1104/pp.104.4.1139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Millar A J, Short S R, Chua N-H, Kay S A. Plant Cell. 1992;4:1075–1087. doi: 10.1105/tpc.4.9.1075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Nagy F, Fejes E, Wehmeyer B, Dallman G, Schafer E. Proc Natl Acad Sci USA. 1993;90:6290–6294. doi: 10.1073/pnas.90.13.6290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.White M J, Kaufman L S, Horwitz B A, Briggs W R, Thompson W F. Plant Physiol. 1995;107:161–165. doi: 10.1104/pp.107.1.161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Anderson S L, Kay S A. Proc Natl Acad Sci USA. 1995;92:1500–1504. doi: 10.1073/pnas.92.5.1500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Carré I A, Kay S A. Plant Cell. 1995;7:2039–2051. doi: 10.1105/tpc.7.12.2039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kay S A, Millar A J. In: The Molecular Genetics of Biological Rhythms. Young M W, editor. Vol. 4. New York: Dekker; 1993. pp. 73–90. [Google Scholar]
- 12.Arpaia G, Loros J J, Dunlap J C, Morelli G, Macino G. Mol Gen Genet. 1995;247:157–163. doi: 10.1007/BF00705645. [DOI] [PubMed] [Google Scholar]
- 13.Takahashi J S, Noboru M, Nikaido S S, Pratt B L, Robertson L M. Rec Prog Hormone Res. 1989;45:279–352. doi: 10.1016/b978-0-12-571145-6.50010-8. [DOI] [PubMed] [Google Scholar]
- 14.Gorton H L, Williams W E, Assmann S M. Plant Physiol. 1993;103:399–406. doi: 10.1104/pp.103.2.399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Takahashi J S. Annu Rev Neurosci. 1995;18:531–553. doi: 10.1146/annurev.ne.18.030195.002531. [DOI] [PubMed] [Google Scholar]
- 16.Johnson C H. Semin Cell Biol. 1994;5:355–362. doi: 10.1006/scel.1994.1042. [DOI] [PubMed] [Google Scholar]
- 17.Millar A J, Straume M, Chory J, Chua N-H, Kay S A. Science. 1995;267:1163–1166. doi: 10.1126/science.7855596. [DOI] [PubMed] [Google Scholar]
- 18.Pittendrigh C S, Daan S. J Comp Physiol. 1976;106:291–331. [Google Scholar]
- 19.Abler M L, Scandalios J G. Maydica. 1994;39:83–88. [Google Scholar]
- 20.Salisbury F B, Denney A. In: Biochronometry. Menaker M, editor. Washington, DC: Natl. Acad. Press; 1971. pp. 292–311. [Google Scholar]
- 21.Lumsden P J. Annu Rev Plant Physiol Plant Mol Biol. 1991;42:351–371. [Google Scholar]
- 22.Halaban R. Plant Physiol. 1968;43:1894–1898. doi: 10.1104/pp.43.12.1894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Sundararajan K S. J Interdiscipl Cycle Res. 1984;15:155–162. [Google Scholar]
- 24.Millar A J, Short S R, Hiratsuka K, Chua N-H, Kay S A. Plant Mol Biol Rep. 1992;10:324–337. [Google Scholar]
- 25.Meyer H, Thienel U, Piechulla B. Planta. 1989;180:5–15. doi: 10.1007/BF02411404. [DOI] [PubMed] [Google Scholar]
- 26.Johnson C H, Knight M R, Kondo T, Masson P, Sedbrook J, Haley A, Trewavas A. Science. 1995;269:1863–1865. doi: 10.1126/science.7569925. [DOI] [PubMed] [Google Scholar]
- 27.Bowler C, Yamagata H, Neuhaus G, Chua N-H. Genes Dev. 1994;8:2188–2202. doi: 10.1101/gad.8.18.2188. [DOI] [PubMed] [Google Scholar]
- 28.Kretsch T, Emmler K, Schäfer E. Plant J. 1995;7:715–729. [Google Scholar]
- 29.Brusslan J A, Tobin E M. Proc Natl Acad Sci USA. 1992;89:7791–7795. doi: 10.1073/pnas.89.16.7791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Frohnmeyer H, Ehmann B, Kretsch T, Rocholl M, Harter K, Nagatani A, Furuya M, Batschauer A, Hahlbrock K, Schäfer E. Plant J. 1992;2:899–906. [Google Scholar]
- 31.Somers D E, Quail P H. Plant Physiol. 1995;107:523–534. doi: 10.1104/pp.107.2.523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Furuya M. Annu Rev Plant Physiol Plant Mol Biol. 1993;44:617–645. [Google Scholar]
- 33.Lam E, Chua N-H. J Biol Chem. 1989;264:20175–20176. [PubMed] [Google Scholar]