

Abstract
The location and density of biologically useful energy sources on Mars will limit the biomass, spatial distribution, and organism size of any biota. Subsurface Martian organisms could be supplied with a large energy flux from the oxidation of photochemically produced atmospheric H2 and CO diffusing into the regolith. However, surface abundance measurements of these gases demonstrate that no more than a few percent of this available flux is actually being consumed, suggesting that biological activity driven by atmospheric H2 and CO is limited in the top few hundred meters of the subsurface. This is significant because the available but unused energy is extremely large: for organisms at 30-m depth, it is 2,000 times previous estimates of hydrothermal and chemical weathering energy and far exceeds the energy derivable from other atmospheric gases. This also implies that the apparent scarcity of life on Mars is not attributable to lack of energy. Instead, the availability of liquid water may be a more important factor limiting biological activity because the photochemical energy flux can only penetrate to 100- to 1,000-m depth, where most H2O is probably frozen. Because both atmospheric and Viking lander soil data provide little evidence for biological activity, the detection of short-lived trace gases will probably be a better indicator of any extant Martian life.
The Martian environment is hostile to life. Typical summer surface temperatures at midlatitudes range between 230 K during the day and 180 K at night (1). Because the atmospheric surface pressure is only 7 mbar (1 bar = 100 kPa), water in liquid form is unstable on the surface (1). The two known water reservoirs, the atmosphere and polar caps, together hold the equivalent of a global layer of water only ≈30 m deep (1). The atmosphere contains 104 times less water than that of Earth, lacks a shielding ozone layer, and is also probably the source of the oxidizing component detected in the top few centimeters of the regolith by the Viking Landers (2, 3). The destructive chemical properties of this oxidant may explain why the Viking biology experiments found no evidence of organic compounds (2–4).
Nevertheless, a number of recent findings have generated excitement about the possibility that life could have existed or might continue to exist on Mars. There is strong evidence that, several billion years ago, the planet was significantly warmer and wetter. Geologists estimate that an initial global inventory of water at least ≈300 m deep is required to explain the ubiquitous erosion and sedimentation features observed by the Viking Orbiters (1). Although anomalous isotopic ratios in the atmosphere suggest that a significant fraction of this water has been lost to space (5), there is abundant geological evidence that ice was once deposited in the regolith, where it should still be present above midlatitudes (6). This ice, which probably extends to several kilometers in depth, could be held at pressures above the triple point and could be a source of liquid water near a magmatic intrusion (1).
It has been suggested that, on Earth, the biomass of subterranean organisms may equal or exceed that at the surface (7). These organisms can live in highly saline conditions at temperatures from 115°C to −20°C (8, 9). Such conditions might exist beneath the surface in an aquifer or hydrothermal system, where the oxidant is absent and the surrounding rock would also protect biota from solar ultraviolet radiation. This makes it conceivable that a subsurface biosphere might exist on Mars. How such systems might obtain energy and carbon is not completely clear, although it is believed that on Earth, the interaction of reduced basalt with groundwater produces H2 whereas carbon is derived from CO2 dissolved in the groundwater (10, 11). On Mars, another potential energy source is the Martian atmosphere, which could act as a photochemical conduit of solar insolation (12, 13).
The most stable molecules containing carbon and hydrogen in the atmosphere of Mars are CO2 and H2O, respectively. Absorption of solar ultraviolet radiation by the atmosphere leads to the production of disequilibrium products such as CO, O2, and H2 (14, 15), as outlined in a comprehensive set of photochemical model by Nair et al. (16):
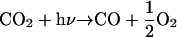 |
R1 |
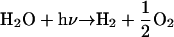 |
R2 |
In the current photochemical models of Mars, the surface is assumed to be biologically inactive and to have no sink or source for the gases in reactions 1 and 2 (R1 and R2). The ultimate fate of the disequilibrium products produced in R1 and R2 is conversion back to the thermodynamically most stable molecules, CO2 and H2O, in a timescale of 5 and 260 yr, respectively. However, if the subsurface of Mars contained an active biosphere, a small fraction of the photochemically produced CO and H2 could diffuse downward into the soil and be consumed as an energy source. These gases would be a likely first choice for many Martian organisms because they are far more abundant than other reduced species on Mars. They are also used as an energy source by many different species of terrestrial organisms (8, 17), as listed in Table 1. Although there are a number of reactions with H2 and CO that could release energy, the most energetic would likely use O2 as an oxidant:
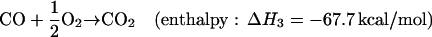 |
R3 |
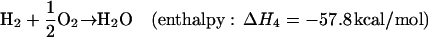 |
R4 |
Reactions of H2 and CO with most other atmospheric gases would not be able to provide nearly as much energy on a globally integrated scale because they would generally have less negative enthalpies and, more importantly, would involve gases that are much less abundant in the Martian atmosphere.§ Methanogenesis (CO2 + 4H2 → CH4 + 2H2O) and acetogenesis (2CO2 + 4H2 → CH3COOH + 2H2O) are probably the next most energetic reactions using atmospheric gases because, although they have enthalpies less negative than R3 and R4, they involve very abundant reactants. There is also a possibility that life could obtain energy from reactions of atmospheric CO2, O2, H2 with one or more of Fe (II), Fe (III), Mn (II), or with gases derived from the subsurface like H2, H2S, SO42−, S0, and CH4. As Table 1 demonstrates, such reactions are catalyzed by a diversity of terrestrial prokaryotes (17).
Table 1.
Representative microbes that use H2 or CO as an energy source
| Energy source | Oxic/anoxic | Organism type | Representative genera | Products of metabolism |
|---|---|---|---|---|
| H2 | Aerobic | Chemolithotrophs | Alcaligenes | H2O |
| Anaerobic | Sulfate reducers | Desulfovibrio | H2S | |
| Sulfur reducers | Desulfuromonas | H2S | ||
| Iron reducers | Shewanella | Fe(II) | ||
| Manganese reducers | Shewanella | Mn(II) | ||
| Nitrate reducers | Pseudomonas | N2, NH3, NOx | ||
| Methanogens | Methanococcus | CH4 | ||
| Acetogens | Acetobacterium | CH3COOH | ||
| CO | Aerobic | Carboxydotrophs | Pseudomonas | CO2 |
| Methylotrophs | Methylosinus | CO2 | ||
| Anaerobic | Methanogens | Methanobacterium | CH4 | |
| Acetogens | Clostridium | CH3COOH | ||
| Sulfate reducers | Desulfovibrio | H2S | ||
| Phototrophs | Rhodobacter | CO2 + H2 |
The list of bacteria here is not all-inclusive but contains representative types and genera. For a more complete treatment of this subject, see ref. 33. A wide variety of products are produced as a result of hydrogen and/or CO oxidation during metabolism. Many of these are atmospheric gases.
Any expectations that these pathways could support Martian life must be tempered by the fact that the Nair et al. model, which included no surface sink (biotic or abiotic), predicted atmospheric H2, CO, and O2 surface mixing ratios in good agreement with the measured (or, for H2, indirectly measured) values (18–20). This qualitatively suggests that any biological activity is not significant enough to affect the gross composition of the atmosphere. It is our intention in this paper to use the Nair et al. model (16) in combination with these measurements to put quantitative limits on the activity and size of a subsurface biosphere that consumes atmospheric H2 and CO for energy. In Section I, we modify the Nair et al. model by adding subsurface sinks for these gases. The purpose is to estimate the largest such sinks that are consistent with measurements of the CO and H2 surface abundances (18, 19). We find that any such surface sinks must be very small (i.e., any fluxes of these gases into the regolith are quite limited). In Section II, we compare these limits to what is potentially achievable by a putative layer of subsurface organisms that catalyze R3 and R4 at the maximum possible rates. We find that these “potential fluxes” are far larger than the limits set in Section I. In Section III, we demonstrate that this discrepancy is significant because the globally integrated energy flux derivable from R3 and R4 is much larger than that from geochemical sources. We conclude in Section IV by discussing other ways that subsurface biota might leave a recognizable signature in the atmosphere.
I. Observational Constraints on the Sizes of Surface Sinks for H2 and CO.
The Nair et al. model, which included no surface sink or source for atmospheric gases, predicts a H2 and CO surface mixing ratio of 37 ppm and 490 ppm (16), in good agreement with the values of 40 ± 10 ppm [inferred by Krasnopolsky et al. (18) from measurements of atomic hydrogen] and 600 ± 150 ppm (ref. 19; measured by the Viking landers), respectively.¶ If the uncertainty on the H2 mixing ratio obtained by Krasnopolsky et al. (18) is taken as the maximum range of possible values, then any subsurface sink added to the Nair et al. model is restricted from perturbing the predicted H2 surface abundance below ≈30 ppm (25%). We sequentially added surface sinks of increasing magnitude to the Nair et al. model to determine the maximum sink permissible by the observations. All H2 that passed into these sinks was considered lost from the atmosphere and unable to return (except, perhaps, as part of water molecules). Because this is somewhat artificial, the effect on the H2 surface abundance should be considered approximate (but certainly correct within an order of magnitude). The results are presented in Fig. 1. The maximum permissible H2 flux into the subsurface is −2 × 107 cm−2⋅s−1. A similar experiment can be conducted with CO, but it is complicated by the fact that the original Nair et al. model predicted a CO mixing ratio that is ≈20% less than the observed value. One reasonable approach is to determine the maximum CO sink that perturbs the predicted surface abundance by <25% (the ratio of the measured value's uncertainty to the measured value). With this we find that the largest permissible CO sink is −6 × 107 cm−2⋅s−1.
Figure 1.

Effect of an H2 surface sink on the H2 surface abundance.
II. Estimation of the Potential Sizes of Biotic Sinks for H2 and CO.
We have estimated the maximum sizes of surface sinks for H2 and CO that are consistent with the Nair et al. model and the measured abundances of these gases. The question we now would like to answer is, How do these observationally derived limits compare with the potential sizes of such sinks that could be produced by subsurface oxidizing biota?
Consider a thin layer of biota living at a depth zb relying on the energy produced by R3 and R4, where the reactants are supplied solely by the atmosphere and diffuse down through the regolith. We will estimate the maximum size sinks that could result from the biological oxidation of H2 and CO. The sizes of the sinks will be determined by the rate of downward diffusion, F (molecules cm−2⋅s−1), specified by F = −D(dn/dz), where n is the number density of the diffusing component (molecules cm−3), z is the depth (cm) with the surface set to z = 0, D is the diffusion coefficient (cm2⋅s−1), and dn/dz is the density gradient (molecules cm−4). According to the Nair et al. model (16), at the surface, the least abundant species in R3 and R4 are CO and H2, respectively. Because the diffusion coefficients for all of the reactants are similar (see below), CO and H2 must then be the limiting reagents in R3 and R4, respectively. If we assume that the biota synthesize the maximum possible amount of H2O and CO2 (i.e., all of the CO and H2 that reaches the biotic layer is oxidized), then we see that, in the region zb < z < 0 in the steady state, dn/dz = Δn/Δz = n(z = 0)/zb.‖
Because the mean free paths of CO and H2 through the background atmosphere are greater than the typical pore size (≈1 μm), molecular collisions with the walls of the pores dominate the transport of these molecules. This is known as Knudsen diffusion, for which we can take D ≈ 2ɛr0/3τ 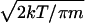 , where r0 is the pore size, ɛ is the porosity, τ is the tortuosity, and m is the molecular mass of the diffusing component (2, 3). Diffusion models of Mars typically use r0 ≈ 6 × 10−4 cm, ɛ ≈ 0.5, and τ ≈ 5 for depths less than ≈1 km, although these numbers are not well constrained (2, 3, 6, 22). At a globally averaged surface temperature of T = 220 K, we find that, above ≈1-km depth, DH2 ≈ 0.97 cm2⋅s−1 and DCO ≈ 0.26 cm2⋅s−1. Using surface densities from (16), we find that for zb < ≈1 km, FH2 = −1.1 × 1013 (zb)−1⋅cm−2⋅s−1 and FCO = −8.8 × 1012 (zb)−1⋅cm−2⋅s−1, where zb has units of cm.** Henceforth, we will often refer to these gas fluxes as “potential” gas fluxes to distinguish them from the limiting fluxes as derived from surface abundance measurements in Section I.
, where r0 is the pore size, ɛ is the porosity, τ is the tortuosity, and m is the molecular mass of the diffusing component (2, 3). Diffusion models of Mars typically use r0 ≈ 6 × 10−4 cm, ɛ ≈ 0.5, and τ ≈ 5 for depths less than ≈1 km, although these numbers are not well constrained (2, 3, 6, 22). At a globally averaged surface temperature of T = 220 K, we find that, above ≈1-km depth, DH2 ≈ 0.97 cm2⋅s−1 and DCO ≈ 0.26 cm2⋅s−1. Using surface densities from (16), we find that for zb < ≈1 km, FH2 = −1.1 × 1013 (zb)−1⋅cm−2⋅s−1 and FCO = −8.8 × 1012 (zb)−1⋅cm−2⋅s−1, where zb has units of cm.** Henceforth, we will often refer to these gas fluxes as “potential” gas fluxes to distinguish them from the limiting fluxes as derived from surface abundance measurements in Section I.
Thus, a biotic layer situated at 10-m depth would produce H2 and CO sinks of −1.1 × 1010⋅cm−2⋅s−1 and −8.8 × 109⋅cm−2⋅s−1, respectively. The latter potential flux corresponds to a potential global carbon flux of 8 Tg − C⋅yr−1, as compared to the terrestrial primary productivity of 2 × 105 Tg − C⋅yr−1. Actually, this H2 potential flux to a biotic layer at 10-m depth is not reasonable because it exceeds the photochemical production rate of H2 in the atmosphere [8.6 × 108 cm−2⋅s−1 (16)]. This is because the potential H2 flux above was calculated using a surface abundance from a photochemical model with no H2 subsurface sink. Thus, the above flux is clearly not sustainable. The maximum potential flux into the surface can be no larger than the production rate (and is probably significantly less), and so we take FH2 = −8.6 × 108 cm−2⋅s−1 for zb = 10 m. This is significantly less than the CO flux. In fact, the CO flux will significantly exceed the H2 flux for zb < ∼1 km, and so CO will be the dominant energy source.
The potential gas fluxes to a biotic layer at 10-m depth just computed are significantly larger than what is permitted by surface abundance measurements. The maximum H2 sink as calculated in Section I is only 2% of the potential H2 flux to biotic layers at 10 m, and 2% of its photochemical production rate. Similarly, the maximum CO sink is 1% of the potential CO flux to biotic layers at 10 m, and 0.03% of its photochemical production rate. This means that the atmospheric gases and their associated energy are going almost completely unused on Mars and that both biological and nonbiological oxidation (e.g., by the regolith oxidant) of atmospheric H2 and CO is extremely limited. Because a biotic layer at even 300 m depth could potentially oxidize H2 at up to 40% of the production rate, this implies that biological activity that uses atmospheric hydrogen is extremely limited in the top few hundred meters of the Martian subsurface for reasons other than energy. Thus, extant biota of this kind is likely to be inaccessible to the current generation of surface landers and rovers planned for the 2005 Mars sample return mission.
III. Implications.
Using the potential gas fluxes calculated in Section II and assuming that the biological oxidation process is 10% efficient, we find that for biotic layers at 10-m depth, the maximum energy fluxes from R3 and R4 are Φ3 = FCO⋅ΔH3/NA⋅10% = 1.0 × 10−13 kcal⋅cm−2⋅s−1 and Φ4 = FH2⋅ΔH4/NA⋅10% = 8.3 × 10−15 kcal⋅cm−2⋅s−1, where NA is Avogadro's number. Note that the present-day ratio of the energy flux provided by these oxidation reactions at 10-m depth to the solar insolation is only 10−8. However, it is unlikely that organisms could use this radiative energy directly because of the extreme conditions on the surface.
How significant is it that most of the available photochemical energy in the atmosphere is not being used by any life? For one, it implies that the apparent scarcity of life near the surface of Mars is not attributable to lack of energy. This is because this photochemical energy is far larger than other biologically usable energy sources in the subsurface. Jakosky and Shock (11) estimated the amount of energy that could be available to Earth and Martian organisms from two geochemical sources: hydrothermal systems and chemical weathering. They found that, on Mars, a total of ≈20 g⋅cm−2 of biota over the last four billion years could be produced from these two inventories. Scaled for today's lower rate of volcanism, the present-day energy flux from these sources is about 60,000, 6,000, and 2,000 times lower than that which could be provided today by the atmosphere to 1-, 10-, and 30-m depth, respectively. The average of these geochemical fluxes (over 4 Ga) is about a factor of 2 closer to the present atmospheric flux. Even more interestingly, the total flux that they calculated for the early Earth—500–800 g⋅cm−2—is 1,500–2,400, 150–240, and 50–80 times lower.
Nevertheless, on Mars, the geochemical flux should dominate in the vicinity of hydrothermal systems in which that energy is highly concentrated [≈0.01 kcal/kg of vent fluid (11)], as well as at greater than 1-km depth, where the atmospheric flux is small. On the cold outskirts of hydrothermal systems, the atmospheric energy could be more important. It is possible that Martian organisms could use reactions that combine gases derived from the atmosphere and those from hydrothermal systems, as occurs near Earth's mid-oceanic ridges. If not, this may mean that the atmospheric energy is not very useful because any organisms would presumably require liquid water. This is because at the depths (0–1 km) at which the atmospheric energy is available, H2O is probably frozen except near hydrothermal systems (1). The availability of water is probably the more important factor that is limiting biological activity on Mars.
IV. Signatures of Subsurface Life.
By estimating the biomass that could be supported by these potential gas fluxes, we now demonstrate that the above results are consistent with limits on the concentration of biological compounds in the Martian soil established by the Viking landers. The maximum biomass that could be supported by these energy fluxes would depend on the minimum amount of power required for survival (the “maintenance energy”). Unfortunately, the maintenance energies of most terrestrial organisms are not well established. Although it has been shown in several instances that the maintenance energy can be virtually 0 over decades at a time (24), in most environments, a finite amount of energy is required. Measurements of the minimum energy requirements of the hydrogen-oxidizing bacteria Alcaligenes eutrophus in typical laboratory conditions give values in the range 0.6–3 × 10−5 kcal⋅(g dry weight)−1⋅s−1 (24, 25). Taking this as a typical value, for a biotic layer at 10-m depth relying on R3 and R4 as described in Section II, the energy flux could maintain a potential biomass of 2 × 10−8 (g dry weight)⋅cm−2. Similarly, for zb = 6 cm, the greatest depth probed by the Viking biology experiments, the density could be as large as 3 × 10−6 (g dry weight)⋅cm−2.‡‡ This is about at the quoted limits of detectability of the Viking Gas Chromatograph-Mass Spectrometer (4, 29). However, we know from Section I that this much energy is not being consumed; if we instead calculate the maximum biomass that could be supported by the smaller gas fluxes permitted by the abundance measurements, then we see that the actual density must be less than 3 × 10−8 (g dry weight)⋅cm−2, which is a factor of 100 below the limits of the Viking Gas Chromatograph-Mass Spectrometer.†† Thus, the nondetection of organic molecules by Viking is consistent with the stringent biomass limits imposed by the H2 and CO surface abundance measurements.
Although, the Viking results as well as the gross composition of the atmosphere suggest there is little life near the surface of Mars, there are still some small differences between the predictions of atmospheric models and spacecraft observations that might be attributable to biological activity. Nair et al. (16) found that their mechanism for oxygen loss, escape from the exosphere, is 40 times larger than the measured value. They noted that diffusion of O2 into the surface at a rate of 6 × 107 cm−2⋅s−1 followed by oxidation of a subsurface reduced gas like H2, CH4, or CO could make up the difference. Although much of this discrepancy appears to have been recently explained by Fox (30), there still is need for an additional sink because her model gives the needed loss only at times of high solar activity whereas the photochemical model was computed at average solar activity. A biosphere that oxidizes upwelling subsurface gases—resembling such a system possibly found on Earth (10)—could provide such a sink.
Although it appears that, on a globally averaged scale, biological consumption of the gases derived from the atmosphere is extremely limited, very active consumption of H2 and CO could still be occurring locally. The presence of such pockets of subsurface life might be detected by observing how the mixing ratios of atmospheric CO, H2, CO2, and H2O deviate on a local scale from the equilibrium values predicted by the photochemical model (16). However, given that the surface abundances of these gases suggest that most of them are not being biologically processed, a better approach might be to search for short-lived trace gases like formaldehyde, methane, or the other products listed in Table 1, many of which have yet to be detected on Mars. Formaldehyde, which has no known geological source on Mars and has a lifetime in the Martian atmosphere of only 13 hours, may have already been detected by the Phobos spacecraft (31), although this is uncertain (32). One might even speculate that spatially resolved measurements of these or other species at the Martian surface by landers, penetrators, or orbiting spacecraft might some day be precise enough so that spatial variations could be used to locate biologically active regions in the Martian interior.
Acknowledgments
We thank A. Haldemann, M. Allen, H. Holland, B. Jakosky, and E. Gaidos for helpful discussions. We also thank R. Goody for reviewing and communicating this work to PNAS. This research is supported in part by NASA Grant NAG5-4022 and the NASA Astrobiology Institute grant to the California Institute of Technology.
Abbreviation
- R1
reaction 1.
Footnotes
Less abundant gases would provide less energy partly because of the lower number of molecules that could be consumed. There would also be a larger energy cost associated with extracting and concentrating them out of the surrounding atmosphere, a process that reduces the entropy of the gas.
Article published online before print: Proc. Natl. Acad. Sci. USA, 10.1073/pnas.030538097.
Article and publication date are at www.pnas.org/cgi/doi/10.1073/pnas.030538097
Note, however, that Antarctic soils typically contain 104 times more organic carbon in nonliving form than in living bacteria (4).
This would overestimate the flux if the diffusing gases experienced adsorption on regolith particles possibly followed by loss due to heterogeneous reactions with the regolith. However, these processes are unlikely to be important because, as demonstrated in the last paragraph of Section II, the Nair et al. model strongly suggests that little of the available CO and H2 is being consumed by the surface (abiotically or otherwise), which would not be the case if the regolith were reactive with these gases. Furthermore, laboratory measurements on analogs of the Martian regolith (21) suggest that CO adsorption would not be significant.
The soil porosity and temperature probably do not vary substantially in the top 1 km (22). The behavior of pore size and tortuosity are less well understood, although they are obviously correlated with porosity. If pore size and tortuosity were fairly constant over the top kilometer, then over that range the flux that would be delivered to a biotic layer would be approximately proportional to depth of the layer. In reality, however, even if the soil parameters were constant, this linear scaling should only be appropriate for depths substantially less than 1 km because the diffusing CO and H2 have finite lifetimes due to photochemical reconversion back to CO2 and H2O. Because the timescale for diffusion to a depth l is approximately l2/D, CO and H2 should only be able to penetrate 64 and 890 m, respectively, before they become depleted by reconversion. Below these depths, the energy sources postulated here are still available, but at a small fraction (≈10%) of what would be estimated from the above linear scaling. This is because the HOx radicals that catalyze the reconversions in the atmosphere should strongly adsorb to the Martian surface, causing them to diffuse into the regolith very slowly (23). Because HOx are short-lived, this means that they will not penetrate very deeply and so will not be encountered during the random-walking of deeper CO and H2 molecules.
This assumes that the energy flux is large enough to be biologically useful. Bacteria are usually restricted from becoming larger than a few micrometers in diameter, in part because their surface area to volume ratio declines with radius r, limiting their energy intake (8). The maintenance energy M is probably proportional to mass (24) and hence to r3, and so the surface area to maintenance energy ratio is A/M∝1/r. For survival, the energy flux must fulfill ΦA ≥ M. Thus, r ≤ kΦ, for some constant k. This means that low-density energy fluxes are capable of supporting only small organisms. There may also be a lower limit (about r = 50 nm) on the size of organisms set by the sizes of the molecules of which they are built (8). If this lower size limit is real, a viable energy source must not be so diffuse that it would require organisms to violate it. For organisms like A. eutrophus (26, 27), which has a fairly high maintenance energy compared to many chemolithotrophs, we find that k = 5 × 107 cm3⋅s⋅kcal−1. Thus, for the energy flux when zb = 10 m, we see that r ≤ 45 nm, which is near the above lower limit. The maximum permissible radius is actually somewhat larger than suggested by this analysis because the radii of cylindrical bacteria are governed by the same flux-radius equation but can be arbitrarily long and thus have significantly more internal space. For the reasons stated above, one might expect that, in energy-starved environments, organisms would be smaller than those living in more energy-rich regions. Given that the absolute lower limit for self-sustaining life is still a matter of some debate, it remains conceivable that some of the diminutive (50 nm in radius) structures interpreted as nanobacteria in the Martian meteorite ALH84001 are such energy-starved resting stages (28).
This CO surface abundance is that from the photochemical model with modified chemical rate constants [case f of Nair et al. (16)].
References
- 1.Carr M H. Water on Mars. New York: Oxford Univ. Press; 1996. [Google Scholar]
- 2.Bullock M, Stoker C, McKay C, Zent A. Icarus. 1994;107:142–154. doi: 10.1006/icar.1994.1012. [DOI] [PubMed] [Google Scholar]
- 3.Hartman H, McKay C. Planet Space Sci. 1995;43:123–128. doi: 10.1016/0032-0633(94)00223-e. [DOI] [PubMed] [Google Scholar]
- 4.Biemann K, Oro J, Toulmin P, Orgel L E, Nier A O, Anderson D M, Simmonds P G, Flory D, Diaz A V, Rushneck D R, et al. J Geophys Res. 1977;82:4641–4680. [Google Scholar]
- 5.Owen T, Maillard J, Debergh C, Lutz B. Science. 1988;240:1767–1770. doi: 10.1126/science.240.4860.1767. [DOI] [PubMed] [Google Scholar]
- 6.Mellon M, Jakosky B. J Geophys Res. 1993;98:3345–3364. [Google Scholar]
- 7.Whitman W, Coleman D, Wiebe W. Proc Natl Acad Sci USA. 1998;95:6578–6583. doi: 10.1073/pnas.95.12.6578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Nealson K. J Geophys Res. 1997;102:23675–23686. [PubMed] [Google Scholar]
- 9.Priscu J, Fritsen C, Adams E, Giovannoni S, Paerl H, McKay C, Doran P, Gordon D, Lanoil B, Pinckney J. Science. 1998;280:2095–2098. doi: 10.1126/science.280.5372.2095. [DOI] [PubMed] [Google Scholar]
- 10.Stevens T, McKinley J. Science. 1995;270:450–454. [Google Scholar]
- 11.Jakosky B, Shock E. J Geophys Res. 1998;103:19359–19364. doi: 10.1029/98je01892. [DOI] [PubMed] [Google Scholar]
- 12.Wolfgang R. Nature (London) 1970;225:876. doi: 10.1038/225876a0. [DOI] [PubMed] [Google Scholar]
- 13.Clark B C. Origins Life. 1978;9:241–248. doi: 10.1007/BF00932498. [DOI] [PubMed] [Google Scholar]
- 14.McElroy M B, Donahue T M. Science. 1972;177:986–988. doi: 10.1126/science.177.4053.986. [DOI] [PubMed] [Google Scholar]
- 15.Hunten D M, Parkinson T M. J Atmos Sci. 1972;29:1380–1390. [Google Scholar]
- 16.Nair H, Allen M, Anbar A, Yung Y, Clancy R. Icarus. 1994;111:124–150. doi: 10.1006/icar.1994.1137. [DOI] [PubMed] [Google Scholar]
- 17.McCollom T, Shock E. Geochim Cosmochim Acta. 1997;61:4375–4391. doi: 10.1016/s0016-7037(97)00241-x. [DOI] [PubMed] [Google Scholar]
- 18.Krasnopolsky V, Mumma M, Gladstone G. Science. 1998;280:1576–1580. doi: 10.1126/science.280.5369.1576. [DOI] [PubMed] [Google Scholar]
- 19.Clancy R, Muhleman D, Berge G. J Geophys Res. 1990;95:14543–14554. [Google Scholar]
- 20.Trauger J, Lunine J. Icarus. 1983;55:272–281. [Google Scholar]
- 21.Leu M, Blamont J, Anbar A, Keyser L, Sander S. J Geophys Res. 1992;97:2621–2627. [Google Scholar]
- 22.Squyres S W, Clifford S M, Kuzmin R O, Zimbelman J R, Costard F M. In: Mars. Kieffer H, Jakosky B, Snyder C, Matthews M, editors. Tucson, AZ: Univ. of Arizona Press; 1992. pp. 523–554. [Google Scholar]
- 23.Anbar A, Leu M, Nair H, Yung Y. J Geophys Res. 1993;98:10933–10940. [Google Scholar]
- 24.Battley E H. Energetics of Microbial Growth. New York: Wiley; 1987. [Google Scholar]
- 25.Muller R, Babel W. Appl Environ Microbiol. 1996;62:147–151. doi: 10.1128/aem.62.1.147-151.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Aragno M, Schlegel H. In: The Prokaryotes. Starr M, Stolp H, Truper H, Balows A, Schlegel H, editors. New York: Springer; 1981. pp. 867–893. [Google Scholar]
- 27.Ingraham J L, Maaloe O, Neidhardt F C. Growth of the Bacterial Cell. Sunderland, MA: Sinauer; 1983. [Google Scholar]
- 28.McKay D, Gibson E, Thomas-Keprta K, Vali H, Romanek C, Clemett S, Chillier X, Maechling C, Zare R. Science. 1996;273:924–930. doi: 10.1126/science.273.5277.924. [DOI] [PubMed] [Google Scholar]
- 29.Burgess E. To the Red Planet. New York: Columbia Univ. Press; 1978. [Google Scholar]
- 30.Fox J. Geophys Res Lett. 1997;24:2901–2904. [Google Scholar]
- 31.Korablev O, Ackerman M, Krasnopolsky V, Moroz V, Muller C, Rodin A, Atreya S. Planet Space Sci. 1993;41:441–451. [Google Scholar]
- 32.Krasnopolsky V, Bjoraker G, Mumma M, Jennings D. J Geophys Res. 1997;102:6525–6534. [Google Scholar]
- 33.Aragno M, Schlegel H G. In: The Prokaryotes. Balows A, Trueper H G, Dworkin M, Harder W, Schliefer K-H, editors. New York: Springer; 1992. pp. 344–384. [Google Scholar]