

Abstract
The developmental trajectory of perceptual organization in humans is unclear. We investigated perceptual grouping abilities across a wide age range (8 to 30 years) using a classic compound letter global/local (GL) task and a more fine-grained microgenetic prime paradigm (MPP) with both few- and many-element hierarchical displays. In the GL task, contrary to adults, both children and adolescents exhibited a classic local bias. In the MPP, all three age groups evinced a bias to individuate the few-element displays; however, the ability to encode the global shape of the many-element displays at the short prime durations increased with age. These results indicate that the full process of garnering shape information from perceptual grouping, which is essential for the ability to do fast and efficient object recognition and identification, develops late into adolescence.
The visual world is rich, complex, and continuously changing. In principle, there are an infinite number of ways to organize visual input; however, despite the ambiguity and complexity of the input and the enormous disparity of experience, humans are remarkably consistent in their interpretation of the incoming sensory information. Studying developmental changes in the ability to perceptually organize this complex visual input and integrate individual visual elements into coherent shapes provides a foundation for understanding how children experience the world as containing bounded, continuous objects. The goal of these experiments was to investigate developmental changes in the ability to structure visual input into coherent global shapes.
There is considerable debate about the developmental trajectory of perceptual organization in humans. Much of the work suggests that infants are capable of grouping visual elements into unitary structures on the basis of lightness similarity (Farroni, Valenza, Simion, & Umiltà 2000; Quinn & Bhatt, 2006; Quinn, Burke, & Rush, 1993), form similarity (Quinn, Bhatt, Brush, Grimes, & Sharpnack, 2002), common fate (Johnson & Aslin, 1995), movement (Kellman, Spelke, & Short, 1986), and proximity (Farroni, et al., 2000). Some studies report that infants may even have greater sensitivity to the global than the local structure in visual stimuli (Freeseman, Colombo, & Coldren, 1993; Frick, Colombo, & Allen, 2000; Ghim & Eimas, 1988; Quinn et al., 1993; Quinn & Eimas, 1986), possibly because they exploit the figural outputs of these foundational Gestalt algorithms.
However, studies with older children suggest that there is a protracted developmental trajectory for the ability to integrate local visual features spatially across the visual field. For example, Kovács and colleagues reported that the ability to integrate collinearly aligned contours against a dense noise background is limited by the spatial range of the local interactions in children and shows significant improvement between the ages of 5 and 14 (Kovács, 2000; Kovács, Kozma, Fehér, & Benedek, 1999; Kozma, Kovács, & Benedek, 2001). Similarly, Hadad and Kimchi (2006) found that 10-year-olds and adults, but not 5-year-olds, can utilize collinearity to enhance closure for the perceptual grouping of shape when the closure-inducing fragments are spatially distant. Finally, several studies have reported that sensitivity to global structure in hierarchical visual stimuli, in which multiple levels of structure exist from the local elements to the global structure, continues to develop into late childhood and early adolescence (Burack, Enns, Iarocci, & Randolph, 2000; Enns, Burack, Iarocci, & Randolph, 2000; Kimchi, Hadad, Behrmann, & Palmer, 2005; Porporino, Iarocci, Shore, & Burack, 2004).
In one illustrative study, Kimchi and colleagues presented children, adolescents, and adults with hierarchical shapes (global diamond) composed of multiple local elements (local squares) in a visual search paradigm (Kimchi et al., 2005). Distracters were similar to targets in local elements, but differed in global shape, or, alternatively, distracters were similar to targets in global shape, but differed in local elements. The shapes included few large local elements or many small local elements. The results revealed that the ability to detect the targets that differed in global configuration from the distracters in the few-element display improved with age, as did the ability to identify targets that differed in local elements from the distracters in the many-element displays. These two conditions are flip sides of the same coin: the former requires the integration of the disparate local elements into a holistic representation and the latter requires the individuation of the elements when the global whole is dominant, by virtue of the texture surface accrued from the presence of the many small less-informative individual elements.
The disparate findings that pervade the literature on this form of visual development suggest that infants’ behavior reflects some basic perceptual organization skills, but that these skills are not fully mature. For example, previous studies with adults have contrasted element clustering, the ability to determine which local elements go together, and shape formation, the ability to identify the boundaries of element clusters, as two different kinds of perceptual grouping (Razpurker-Apfeld & Kimchi, 2007; Trick & Enns, 1997). Infants may show an early ability to determine what elements cluster together (as alluded to above), but are much less skilled at organizing such clusters into integrated, discrete shapes. It is also possible, perhaps counterintuitively, that global shape processing is not fully mature even in young adolescents.
In the present study, we systematically evaluated perceptual organization, and sensitivity to global and local information in complex visual displays, in individuals across a wide age range (8 to 30 years). To do so, we conducted two investigations. In the first, we employed a classic test of global and local processing using compound letter stimuli (Navon, 1977) to evaluate the developmental trajectory of precedence for global information in visual representations. Second, we investigated developmental differences in the content (global configuration and/or local elements) of entry-level units and longer term representations of hierarchical visual displays using a more fine-grained microgenetic approach (Kimchi, 1998).
An important issue to clarify is what we mean by global and local information and how it differs from other usages of “holistic” and “analytic” information in the perceptual development literature (see Kimchi, 1992). Here, by global information, we are referring to stimulus properties that derive from interrelations between component, or local, parts (e.g., Garner, 1978; Navon, 1977; Rock, 1986). We are particularly interested in the perception of the global property of shape, which is derived from the spatial relations among the individual elements in the visual display, because this information is especially relevant for determining object form and object identity. We are not evaluating the kinds of analytic and holistic processing that have been the focus of the holistic-to-analytic shift in the perceptual development literature (Shepp & Schwartz, 1976; Smith & Kemler, 1977), which described a shift in category learning that serves as the basis for the ability to identify commonalities across exemplars. We are studying processes involved in encoding specific exemplars as unique perceptual entities with specific properties. The developmental trajectories may be very different for the abilities to detect commonalities across objects versus to identify unique attributes of objects.
In our studies, children were required to be able to detect both the local and the global properties in visual displays before they could participate, which indicates that both levels of stimulus structure are psychologically real and available to our participants. The focus of these studies was to address when developmentally global properties, and global shape in particular, become primary during the temporal formation of a percept. This primacy of global shape processing is essential for the ability to do fast and efficient object recognition and identification (Behrmann & Kimchi, 2003), particularly when having to identify individual objects.
Method
General Methodology
Participants
The participants in this study were volunteers and included children, adolescents, and adults from the Greater Pittsburgh area. All participants were healthy individuals with normal or corrected vision and no history of neurological or psychiatric illness in themselves or in their first-degree relatives. The sample included 15 children (age range: 8 -13 years; M = 11 years, SD = 1.0 years; 10 males), 15 adolescents (age range: 14 - 17 years; M = 15 years, SD = 1 year; 12 males), and 12 adults (age range: 20 - 30 years; M = 26 years, SD = 4 years; 12 males). The participants were 94% Caucasian and 6% African American and were from families with an average socioeconomic status. Results from the adults were previously published as part of a larger study of perceptual processing in autism (Behrmann, Avidan, Leonard, Kimchi, Luna, Humphreys, et al., 2006). Participants and/or their legal guardians provided informed consent prior to participating in the study. All the experimental procedures complied with the standards of the Internal Review Boards at both the University of Pittsburgh and Carnegie Mellon University. Participants were paid $10.
General Procedure
The experiments were conducted on a Dell Inspiron 3200 (14-inch monitor) laptop computer and were executed with E-Prime software version 1.1 (Schneider, Eschman, & Zuccolotto, 2001). Participants made keyboard responses. Reaction time (RT), measured from the onset of the stimulus choice screen, and accuracy were recorded for all tasks. Participants executed the tasks in a dimly lit room at a viewing distance of approximately 60 cm from the screen. All tasks were completed in a single session, the order of which was counterbalanced across participants within each age group.
Data Analyses
Mean RTs for each participant were submitted to a log transformation to establish homogeneity of variance prior to being submitted to the analyses. Only correct trials were analyzed for RT differences across the experimental conditions. Error was measured as the proportion of incorrect items within each task block. Age group differences were evaluated using repeated-measures ANOVAs with the appropriate within-subject experimental factors and age group as a between-subjects factor. Interactions from the ANOVA analyses were investigated with Tukey HSD post-hoc comparisons. In both experiments, adults performed at ceiling in accuracy (see Behrmann et al., 2006), therefore, their error data were not included in the analyses of age-group differences in the sensitivity to local and global information. Effect sizes are reported as Partial Eta2.
Global/Local Processing with Compound Stimuli
In our first experiment, we employed the well-known hierarchical compound stimuli (Navon, 1977, 2003), which included global letters composed of smaller local letters that were either consistent or inconsistent with the global letter in identity (see Figure 1). This design allows for performance measures, such as the speed of identification and asymmetric interference during inconsistent trials, to be used to infer the advantage of one level over the other (Navon, 1977, 1983). TD adults exhibit a global advantage (i.e., faster identification of the global letter and asymmetric global-to-local-interference) when performing this task. We adopted this paradigm for use with children and adolescents, and compared their performance with that of adults to evaluate developmental changes in the precedence of global information. We used a focused attention version of the task in which participants identify the global or local letter in separate blocks since divided attention versions reduce the precedence of global information for TD adults (Hoffman, 1980, Kimchi, Gopher, Rubin, & Raij, 1993) and we wanted to maximize the potential for children and adolescents to demonstrate sensitivity to the global information.
Figure 1.
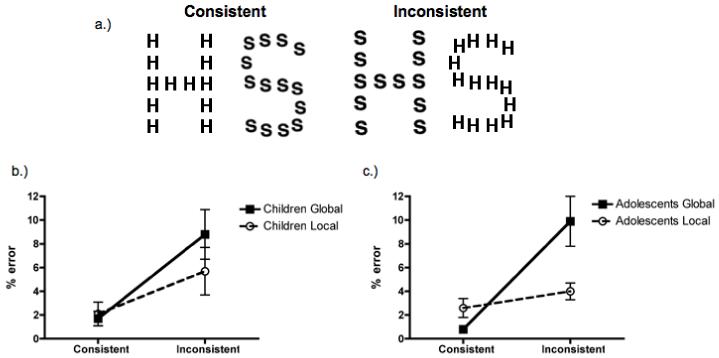
A.) Examples of hierarchical letter stimuli used to evaluate developmental differences in sensitivity to global (big letter) and local (small letters) information. Mean error ± 1 SEM for children (b) and adolescents (c) in the global/local task. Both developmental groups demonstrated local precedence in the pattern of their errors in this task.
Design and Procedure
Figure 1a shows that the stimuli included four hierarchical letters of two types: consistent letters, in which the global and local letters were matched on identity (i.e., a large H made of smaller Hs) or inconsistent letters, in which the letters had different identities at the two levels (i.e., a large H made of smaller Ss). The global letter subtended 3.2° in height and 2.3° in width, and the local letter subtended 0.44° in height and 0.53° in width.
The two tasks, global or local letter identification, were administered in separate blocks of 96 experimental trials each, preceded by 10 practice trials, for a total of 192 trials. There were an equal number of consistent and inconsistent stimuli, which were randomized within a block. In order to ensure that participants could identify both levels of information in the stimuli, before the experiment began, children and adolescent participants were required to manually trace with their finger the “big” letter and the “little” letter in a consistent stimulus and in an inconsistent stimulus. At the onset of each block, participants were verbally instructed whether to identify the ‘big’ or ‘little’ letter. Each trial started with a central fixation cross for 500 ms. One of the four possible stimuli immediately replaced the fixation and remained on the screen until a response was made. Participants were instructed to press the ‘s’ key with their left index finger or the ‘h’ key with their right index finger to indicate their response. The order of the blocks was randomized for each participant.
Results
Error rate
Figures 1b and 1c show the mean error rate plotted as a function of task and consistency for children and adolescents. A repeated-measures ANOVA with the factors of task (global, local), consistency (consistent, inconsistent), and age group (children, adolescents) failed to reveal a main effect of age group, F(1, 28) = 0.0, p = n.s. Children and adolescents were equally accurate when performing the global/local task and, in general, error was low (4.6% for children and 4.3% for adolescents), confirming that both groups could perform the task.
As predicted, there was a significant task × consistency interaction, F(1, 28) = 7.8, p < .01, ηp2 =.22. Both children and adolescents exhibited more than three times the amount of local-to-global interference (M = 8.1%, SD = 8.0%) than global-to-local (M = 2.5%, SD = 7.0%) interference, p < .01, meaning they overtly reported the identity of the ‘little’ letter when instructed to identify the ‘big’ letter. There were also main effects of task, F(1, 28) = 5.0, p < .05, ηp2 = .15, and consistency, F(1, 28) = 30.8, p < .001, ηp2 = .52. Children and adolescents made fewer errors in the local (M = 3.6%, SD = 3.1%) compared to the global (M = 5.3%, SD = 4.2%) task and in the consistent (M = 1.8%, SD = 1.9%) compared to inconsistent (M = 7.1%, SD = 5.3%) trials. There were no interactions between any of the factors and age.
The pattern of errors reveals that children and adolescents both exhibited local precedence during the global/local task, which included fewer errors at the local level and asymmetrical local-to-global interference. There were no age group differences in this sensitivity to the local information in accuracy.
Reaction Time
Figure 2 shows the mean reaction time plotted as a function of task and consistency separately for children (2a), adolescents (2b), and adults (2c). There were significant interaction of task × consistency × age group, F(2, 39) = 8.6, p < .001, ηp2 = .31, as well as of task × age group, F(2, 39) = 7.3 p < .005, ηp2 = .27, interactions. There were also main effects of age group, F(2, 39) = 16.8, p < .001, ηp2 = .46, and consistency, F(1, 39) = 142.7, p < .001, ηp2 = .79. Separate analyses within each age group revealed that the adults demonstrated the classic pattern of global precedence: they were faster to identify letters at the global compared to the local level, F(1, 11) = 25.1, p < .001, ηp2 = .70, and showed more global-to-local than local-to-global interference; there was a task × consistency interaction, F(1, 11) = 5.5, p < .05, ηp2 = .33. Adults exhibited roughly twice as much global-to-local (M = 77 ms, SD = 50 ms) as local-to-global interference (M = 31 ms, SD = 25 ms).
Figure 2.
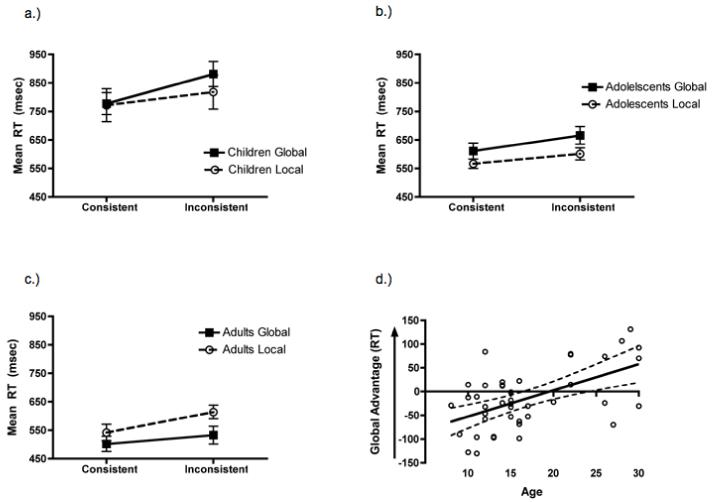
Reaction time results from the global/local task. Mean reaction time ± 1 SEM for children (a), adolescents (b), and adults (c) in the global/local task. Adults showed the classic pattern of global precedence. Both development groups demonstrated local precedence in this task. D.) Global advantage in reaction time [(local inconsistent - local consistent) - (global inconsistent - global consistent)] plotted as a function of age with 95% confidence intervals. The global advantage in RT significantly increases with age, p < .001.
Contrary to the pattern of adult performance, adolescents showed more of a local bias. They were faster to identify letters at the local compared to global level, F(1, 14) = 8.6, p < .01, ηp2 = .38, and to identify consistent compared to inconsistent items, F(1, 14) = 63.4, p < .001, ηp2 = .82. However, they did not exhibit asymmetric local-to-global interference; indeed, there was no task × condition interaction, F(1, 14) = 2.2, p = n.s. Similarly, children showed a local bias, but only in the asymmetric pattern of interference; there was a task × consistency interaction, F(1, 14) = 9.7, p < .001, ηp2 = .41, but no main effect of task, F(1, 14) = 1.4, p = n.s. Children were equally fast to identify the local and global letters, but they exhibited twice as much local-to-global (M = 104 ms, SD = 50 ms) as global-to-local interference (M = 46 ms, SD = 60 ms).
Finally, in order to evaluate the transition from a local bias to a global advantage in perceptual processing in a more continuous way, a measure of global advantage was computed for each participant {(local inconsistent - local consistent) - (global inconsistent - global consistent)} and submitted to a regression with age. Interestingly, there was a significant effect of age, F(1, 40) = 17.8, p < .001. As is evident from Figure 2d, with increasing age, there is a concomitant increase in global precedence, although we note that there is quite a bit of variance across the age range, too.
Speed/Accuracy Trade Off
Pearson product correlational analyses of mean reaction time and mean accuracy revealed that children did not trade speed for accuracy in either task (global, local) at either level of consistency. Adolescents only tended to trade speed for accuracy on global inconsistent trials, r = -0.51, p = .052, two-tailed, in which RT was fast but error rate was somewhat high, relative to the other conditions. This provides further evidence that performance on the global inconsistent condition is showing the most transformation with age.
Gender Differences
In order to evaluate the possibility that the developmental differences in perceptual processing biases were due to a disproportionate distribution of males and females in the age groups (no females in adult group), the analyses were re-run with only male participants. The pattern of results was identical, indicating that the age effects we have observed cannot be accounted for by differences in male/female ratios. In the RT data, there was a significant task × consistency × age group interaction, F(2, 31) = 8.0, p < .005, ηp2 = .34. Separate analyses within the age groups revealed that the adolescents were significantly faster to identify local than global letters, F(1, 11) = 6.7, p < .025, ηp2 = .38, and consistent than inconsistent items, F(1, 11) = 64.3, p < .001, ηp2 = .85. As in the analyses that included females, the adolescents did not show asymmetrical patterns of interference, F(1, 11) = 2.8, p = n.s. Male children also showed the local bias. They showed more local-to-global than global-to-local interference, F(1, 9) = 7.7, p < .025, ηp2 = .46, and they were faster to identify consistent than inconsistent items, F(1, 9) = 94.0, p < .001, ηp2 =.91. Together these results indicate that the developmental differences in perceptual processing biases in this task were not the result of gender differences in perceptual processing.
Discussion
The goal of this study was to evaluate the developmental trajectory of the emergence of global precedence in visuoperceptual processing. We found no evidence of global precedence in the visual representations of either children or adolescents. Both groups showed a classic local bias when processing the hierarchical forms. They were faster and more accurate when identifying letters at the local compared to at the global level. Also, both groups demonstrated more local-to-global interference in reaction time and accuracy than global-to-local interference. Finally, the tendency to exhibit global precedence in this task increased linearly with age.
These results are consistent with two other studies that failed to find evidence of global precedence using similar compound stimuli in typically developing individuals across a similar age range (Mottron, Burack, Stauder, & Robaey, 1999; Mottron, Geldart, Maurer, & de Schonen, 2003) and one study that reported developmental improvements in global processing of these same compound stimuli, particularly when they were composed of fewer, more sparse elements in 4- and 6-year-olds (Dukette & Stiles, 1996). However, our findings are inconsistent with several studies that have also evaluated infants’ and children’s visuoperceptual processing biases (Cassia, Simion, Milani, & Umiltà, 2002; Mondloch, Geldart, Maurer, & de Schonen, 2003; Ozonoff, Strayer, McMahon, & Filloux, 1994; Plaisted, Swettenham, & Rees, 1999; Rinehart, Bradshaw, Moss, Brerenton, & Tonge, 2000), which may be explained by differences in task parameters that differentially affected children’s perceptual biases.
In adults many parameters affect the global advantage (see Kimchi, 1992), including the length of the exposure duration and the spacing between and number of local elements, which are especially relevant to differences in the developmental findings. In some adult studies, asymmetrical global-to-local interference has only been observed under very short exposure durations (Paquet & Merikle, 1984) and when there are many local elements close together that generate good exemplars of the global figure (Kimchi, 1998; Martin, 1979; Navon 1983). In the current study, participants were given an unlimited stimulus exposure duration even though several of the previous developmental studies used a limited duration (Ozonoff et al., 1994; Plaisted et al., 1999). The unlimited exposure duration may have resulted in behavioral responses that are more reflective of the final percept and not a temporally early processing bias. Also, previous experiments reporting a global advantage effect in children often used a stimulus that contained many (56) small, densely packed local letters (Ozonoff et al., 1994; Plaisted et al., 1999), a scenario known to strongly bias any existing global representation (Kimchi, 1998). In contrast, our stimuli included a global letter made from either 12 (H) or 14 (S) more sparse local letters. One potential resolution, then, is that perceptual organization involves multiple processes that vary in developmental trajectories. Kimchi and colleagues (2005) found evidence for the early development of perceptual grouping for many, small element items. However, the ability to perceptually group few large elements continues to develop into late childhood.
In the next experiment, we evaluated whether children and adolescents exhibit an early bias to encode the global shape of a hierarchical stimulus, but find the local elements more salient and dominant in their final percept, and whether they show more of a global advantage in response to many-element stimuli.
Microgenetic Analysis of the Perceptual Organization of Hierarchical Stimuli
In order to investigate whether there are developmental differences in 1) the initial and subsequent encoding of global and local information and 2) perceptual grouping and individuation abilities when local items are few large or many small, we adopted a microgenetic approach using a primed matching paradigm developed by Kimchi (1998, Expt 1). A microgenetic approach is particularly powerful because it reveals how the internal representation of a visual stimulus develops over time by varying the duration of the prime to tap earlier and later internal representations (Sekuler & Palmer, 1992). In this particular task, participants view (but ignore) an ambiguous prime followed immediately by a pair of test figures (probes) and judge whether the two probes are same or different. The prime and probe stimuli include patterns (i.e., global diamonds composed of smaller circles) with few large elements or with many small elements. Each test stimulus includes two probes from one of two conditions, defined by their similarity to the prime stimulus (see Figure 3). In the element-similarity (ES) condition, probes are similar to the prime in their local elements (circles), but differ in their global configuration (global square instead of global diamond). In the configuration-similarity (CS) condition, probes are similar to the prime in their global configuration (diamond), but differ in their local elements (local squares instead of local circles). The prime is presented at several durations, providing multiple temporal windows over which the representation evolves prior to the onset of the probe, and behavioral responses are compared across the prime durations.
Figure 3.
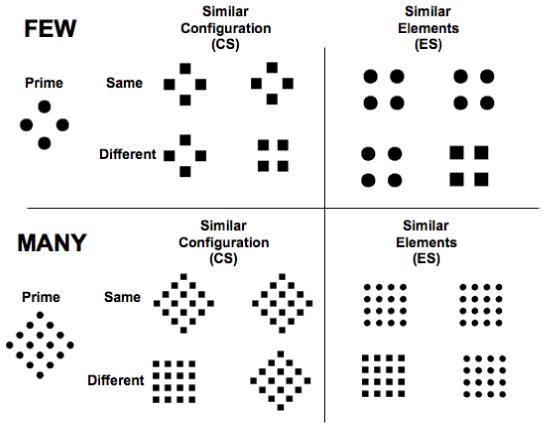
Examples of hierarchical shape stimuli used in microgenetic few/many task.
This paradigm reveals the entry-level units of a developing representation by comparing behavioral responses across the prime durations. In other words, at short prime durations only the most dominant characteristic of the percept of the priming stimulus is represented and can act as a prime. When test figures share this basic structure, or the entry-level units of representation, with the prime stimulus, responses will be facilitated. Such entry-level units may encode individual elements of hierarchical visual displays, clusters of some of these elements, or even the entire global shape of the visual forms. Other superordinate and subordinate units of the representation may develop over longer prime durations, revealing what is encoded at various stages in the microgenesis of the percept (see Kimchi, 1998).
Kimchi (1998) showed that, in adults, for patterns with few large elements, the entry-level units are, indeed, the elements and there is only a weaker representation of the global configuration. In other words, when looking at the few-element items, adults were faster to make similarity judgments about probes that shared local elements with the prime and took much longer to make such judgments about probes that shared a global configuration but differed in the local elements. On the other hand, for patterns with many, small elements, Kimchi found that the entry-level units represent the global configuration and that the individuation of the local elements requires more time and focused attention. In other words, when looking at the many-element items, adults were faster to make similarity judgments about probes that shared a global configuration, regardless of the local elements. Interestingly, this access to the global configuration was apparent even at the shortest prime duration (40 ms).
We employed this same paradigm with those participants who completed Experiment 1 to evaluate whether perceptual organization develops along the local - global integrative dimension, regardless of the number of local elements, or whether different perceptual grouping processes develop along different developmental trajectories.
Methods
Participants
The participants included the same 15 children, 15 adolescents, and 10 adults (age range: 20 - 30 years; M = 25 years, SD = 4 years; 10 males) from the Global/Local experiment. Two of the adults who participated in the global/local experiment did not participate in the microgenetic priming study.
Design and Procedure
This experiment used the primed matching paradigm described above. In a single block, only few- or many-element items were presented. The few-element prime consisted of a global diamond made of four relatively large circles, which subtended 0.36° in diameter. The local squares in the few-element probe stimuli subtended 0.38° in diameter. The many-element prime was a global diamond made of 16 relatively small circles, which subtended 0.18° in diameter. The local squares in the many-element probe stimuli subtended 0.19° in the many-element patterns. Both global size and the ratio of element size to the distance between elements were matched in the few- and many-element figures even though they differed in terms of the number and size of the local elements. The global diamond subtended 1.25° of visual angle, and the global square 96°. There was a distance of 7cm between the centers of the two probe stimuli in each test pair.
The few-element and many-element stimuli were administered in separate blocks of 160 trials, each of which was preceded by 16 practice trials, for a total of 320 experimental trials. Before each experimental block, participants were required to identify the “big” shape and the “little” shape in a few-element stimulus or in a many-element stimulus. Each trial began with a small fixation dot that appeared in the center of the screen for 250 m, followed by a prime. The presentation time for the prime stimulus could occur at one of five durations, 40, 90, 190, 390, or 690 ms. A probe test pair appeared immediately after the prime on either side of the location previously occupied by the prime. The test pair stayed on the screen until the participant responded, or until a maximum of 3000 ms. Participants had to decide whether the probes in the test pair were “exactly the same” or “different in any way”. They were instructed to respond “as quickly as possible without making mistakes” with their left index finger on the ‘s’ key if the items were identical or on the ‘d’ key with their right index finger if the items were different. All combinations of the three factors (prime duration, condition, and response) were randomized within a block, with each combination occurring on an equal number of trials.
RT responses were only evaluated for correct same trials since the relationship between the prime and the test pair is only fully controlled when the two items in the test pair are the same. Also, previous studies using similar paradigms in adults have only found priming effects for same responses and have failed to find priming effects for different responses (Beller, 1971; Kimchi, 1998). Responses from the two tasks were analyzed separately since the few-element and many-element displays elicit different kinds of grouping and individuation processes (Kimchi et al., 2005).
Results
Error rate
Figure 4a shows the mean error rate in the Few-element task for same responses plotted as a function of prime duration and condition for children and adolescents. In the Few-element task, children and adolescents were equally accurate, with error rates generally low (5.5% for children and 9% for adolescents). A repeated-measures ANOVA with the factors of condition (CS, ES), prime duration (40, 90, 190, 390, 690), and age group (children, adolescents) failed to reveal a main effect of age group, F(1, 28) = 2.0, p = n.s. Across all prime durations, participants made fewer errors during the ES (M = 4.5%, SD = 5.5%) compared to the CS (M = 10.0%, SD = 10.6%) trials; there was a main effect of condition, F(1, 28) = 9.3, p < .005, ηp2 = .25. There were no other main effects or interactions among any of the factors. These results reflect a strong bias to encode the local information within the few-element displays at all prime durations for both children and adolescents.
Figure 4.
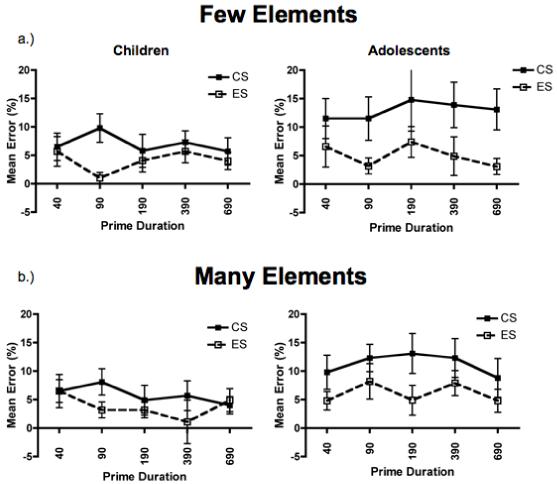
Accuracy results for Few (a.) and Many (b.) tasks, plotted as mean percent error ± 1 SEM separately for children and adolescents. Across both tasks, children and adolescents were less accurate at identifying the configural (CS) compared to the elemental (ES) information.
As in the Few-element task, children and adolescents were equally accurate in the Many-element task (5.8% errors for children and 8.7% errors for adolescents) (see Figure 4b). Although there was no main effect of age group, F(1, 28) = 2.6, p = n.s., there was a statistical trend for a main effect of condition, F(1, 28) = 3.4, p = .075, ηp2 = .11. As in the Few-element task, across all prime durations participants tended to make fewer errors in the ES (M = 5.9%, SD = 5.3%) compared to the CS (M = 8.5%, SD = 7.5%) condition; however, there was also a statistical trend for a condition × age group interaction, F(1, 28) = 3.4, p = .092, ηp2 = .10. Children tended to be less affected by the condition than were adolescents.
This pattern of results indicates that both children and adolescents are more accurate at encoding the local elements than the global configuration in the few-element displays and they tend to be more accurate at encoding the local elements in the many-element displays as well. This was true across all prime durations, suggesting that both the entry-level units and the most salient aspects of children’s and adolescents’ representations for these hierarchical displays include the local elements.
Reaction Time
Figure 5 shows the mean RT for the Few-element displays for same responses plotted as a function of prime duration and condition for children (5a), adolescents (5b), and adults (5c). There was a main effect of age group, F(2, 37) = 13.1, p < .001, ηp2 = .42. Bonferroni corrected post-hoc comparisons revealed that children were slower to discriminate the probe stimuli than were adolescents and adults, p < .001, who were not different from one another. Besides the generally slower RTs from children, the three age groups performed very similarly in the Few-element task. Participants were consistently faster to discriminate the probe stimuli in the ES condition than in the CS condition; there was a main effect of condition, F(1, 37) = 60.9, p < .001, ηp2 = .62. Across all three groups of participants and prime durations, there was an 89 ms advantage for the ES compared to CS condition. Although there was no condition × age group interaction, F(2, 37) = 2.1, p = n.s., children (106 ms) and adults (108 ms) showed twice the advantage for ES compared to CS, as did adolescents (56 ms). Finally, participants were consistently affected by the prime duration, F(4, 148) = 1.3, p < .024, ηp2 = .07. Bonferroni corrected post-hoc comparisons revealed that participants were faster to discriminate probe items at the 190 and 690 ms prime durations compared to the 40 ms prime duration, p < .05. There were no interactions between any of the variables. In summary, although children were slower to respond in the Few-element task, like adolescents and adults, they were more sensitive to the element information and slower to detect the configural information in the few-element displays.
Figure 5.
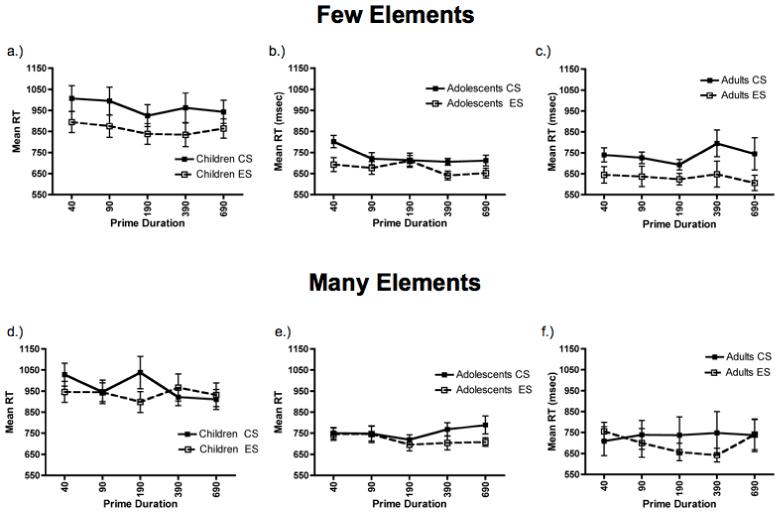
Reaction time results for Few (a-c) and Many (d-f) task, plotted as mean percent error ± 1 SEM separately for children (a, d), adolescents (b, e), and adults (c, f). In the Few task, all three age groups were faster to identify the local elements (ES) compared to the global configuration (CS) in the few-element displays. In the Many task, adults showed an early advantage for the global configuration (CS) compared to the elemental information (ES) at the 40 ms prime duration. Neither children nor adolescents demonstrated this advantage. In fact, both developmental groups showed an element advantage, which occurred at the early prime durations for the children and at the later prime durations for the adolescents.
Of great interest, there were large developmental differences in the Many-element task. Figures 5d - 5f show the mean RT for the many-element displays for same responses plotted as a function of prime duration and condition for each age group. There was a significant condition × prime × age group interaction, F(8, 148) = 3.6, p < .001, ηp2 = .16. There were also main effects of age, F(2, 37) = 9.9, p < .001, ηp2 = .35, and condition, F(1, 37) = 5.6, p < .025, ηp2 = .13. Separate analyses within each age group revealed that adults’ sensitivity to configural and elemental information was affected by the prime duration; there was a significant condition × prime interaction, F(4, 36) = 2.9, p < .05, ηp2 = .25. A priori planned comparisons demonstrated that adults were faster to discriminate probe stimuli in the CS compared to the ES condition at the shortest prime duration of 40 ms, t(9) = 2.5, p = .036, two-tailed. They exhibited a 46 ms advantage for the CS compared to the ES condition. This replicates the original finding with adults using this paradigm (Kimchi, 1998). Adults were equally fast to discriminate probe items in the CS and ES conditions in the many-element displays at all other prime durations.
Adolescents tended to be consistently faster to discriminate probe stimuli in the ES the compared to the CS condition in the Many-element displays; there was a trend for a main effect of condition, F(1, 14) = 4.4, p = .053, ηp2 = .24. They exhibited a 36 ms advantage for the ES compared to the CS condition across all prime durations. There was also a statistical trend for a condition × prime interaction, F(1, 14) = 2.0, p = .11, ηp2 = .12. A priori planned comparisons revealed that adolescents were equally fast to discriminate probe stimuli on the basis of elemental and configural information at the short prime durations and showed a 64 ms advantage for the ES condition at the 390 ms prime duration, t(14) = 2.2, p < .05, two-tailed, and an 81 ms advantage at the 690 ms prime duration, t(14) = 2.4, p < .05, two-tailed.
Children demonstrated a slightly different pattern of heightened sensitivity to the elemental information in the many-element displays. There was no main effect of condition, F(1, 14) = 1.7, p < .n.s., but there was a significant condition × prime interaction, F(4, 56) = 4.1, p < .01, ηp2 = .23. A priori planned comparisons revealed that children were faster to discriminate probe stimuli on the basis of elemental compared to configural information at the 40 ms, t(14) = 2.2, p < .05, two-tailed, and 190 ms, t(14) = 3.2, p < .005, two-tailed, prime durations. They demonstrated an 82 ms advantage in the ES condition at the 40 ms prime duration and a 140 ms advantage at the 190 ms prime duration.
In order to represent the developmental differences in performance on the Many-elements task more clearly, mean RT differences scores (CS-ES) were computed for each age group for each prime duration (see Figure 6). A repeated-measures ANOVA on this difference score with the factors of age group and prime duration, revealed a significant prime duration × age group interaction, F(8, 148) = 3.8, p < .001, ηp2 = .17. Separate one-way ANOVAs performed at each prime duration showed significant age group differences at the 40 ms, F(2, 37) = 3.8, p < .05, 190 ms, F(2, 37) = 6.3, p < .005, and 690 ms, F(2, 37) = 3.7, p < .05, durations. Bonferroni corrected post-hoc comparisons revealed that children showed a stronger element bias (i.e., shorter RTs in the ES condition) than adults at the 40 ms, p < .05, and 190 ms, p < .005, prime durations. Children tended to show more of an element bias than adolescents at the 190 ms prime duration, p = .066. Finally, adolescents tended to show more of an element bias than adults at the 690 ms prime duration, p = .056.
Figure 6.

Summary of developmental differences in Many-element reaction time results, plotted as a function of element bias (configural similarity - element similarity) for each age group.
In summary, neither children nor adolescents exhibited the adult configural advantage at the short 40 ms prime duration for the Many-element displays. In fact, both children and adolescents were faster at encoding the element information, at the earlier prime durations for the children, and at longer prime durations for the adolescents.
Speed/Accuracy Trade Off
Pearson product correlational analyses of mean RT and mean error revealed that neither children nor adolescents traded speed for accuracy during the Few-element task for either condition at any prime duration. However, in the Many-element task, children traded speed for accuracy during element-similarity trials at the shortest 40 ms prime duration, r = -0.71, p < .001, two-tailed. Ten of the 15 children made no errors in this condition and this speed-accuracy trade off was largely due to the performance of two children who scored less than 75% correct in this particular condition and exhibited very slow RTs. When these two individuals were taken out of the analyses, the results revealed that the group as a whole, was not trading speed for accuracy in this condition, r = -.47, p = n.s. Ultimately this means that these two individuals contributed fewer trials with longer RTs to the analyses of RT differences in the ES condition at 40 ms. If anything, data from these two individuals may have reduced the magnitude of the element bias (i.e., faster RTs in the ES condition) in the children, instead of amplified it. Finally, adolescents traded speed for accuracy during the configuration-similarity trials at the longest 690 ms prime duration, r = -0.55, p < .05, two-tailed.
Gender Differences
Again, to evaluate the possibility that the developmental differences in perceptual processing biases in the Many-elements task were due to a disproportionate number of males and females in each age group, the analyses were re-run with only male participants. The pattern of results was the same. Among the male participants’ RT data, there was a significant condition × prime duration × age group interaction, F(8, 116) = 2.6, p < .025, ηp2 = .15. Separate one-way ANOVAs the CS-ES difference score with the factor of age group, revealed a significant effect of age group at the 40 ms, F(2, 29) = 3.6, p < .05, and at the 190 ms, F(2, 29) = 4.1, p < .05, prime durations. Children tended to show a stronger element bias than adolescents and adults at the 40 ms prime duration, p = .06, and a significantly stronger element bias than adults at the 190 ms prime duration, p < .05. These results indicate that the developmental differences in perceptual processing biases in the Many-elements task are not attributable to gender differences in perceptual processing.
Discussion
The goals of this experiment were to investigate whether there are developmental differences in 1) the initial and subsequent encoding of global and local information and 2) perceptual grouping and individuation abilities. First, we replicated the original findings with adults (Kimchi, 1998), providing more support for the notion that adults individuate few large element displays and perceptually group many small element displays. Second, we did not observe developmental differences in response to the few-element displays. All three age groups were biased to individuate these displays into the local elements and demonstrated a relatively weak representation of the global configuration. This was true across all prime durations suggesting that the local elements dominate the entry-level units and the final percept of the representations of few-element displays across the age range tested.
Third, we observed dramatic developmental differences in the way individuals process the many-element displays. Adults were the only group to show the early advantage for encoding the global configuration. At the other end of the spectrum, children continued to be biased to encode the local elements of these many-element displays, particularly at the short prime durations, indicating that the entry-level units for their representations of these hierarchical forms appear to be the local elements. However, given enough time, children do encode a weak representation of the global form, suggesting that both kinds of information are available in their final percept. Adolescents began to demonstrate the beginnings of the early global advantage. At the short prime durations, adolescents were equally fast to encode the global form and local elements; however, they were more accurate when encoding the local elements. At later prime durations, adolescents were faster and more accurate to encode the local elements. This pattern of results suggests that the representation of the global configuration is becoming stronger in the initial encoding of the many-element displays, but the local information still dominates the final percept. In summary, our results suggest that the entry-level units of both few- and many-element hierarchical displays are dominated by the local elements through childhood and only begin to incorporate the global configuration in adolescence.
It is important to note that we did not include a neutral prime (as the experiment was already long for the young children), as in the original study (Kimchi, 1998), which prevented us from directly investigating the relative costs and benefits of processing the local elements versus the global configuration, relative to a neutral baseline. Follow-up studies in which a performance baseline is initially established with ES and CS displays following a neutral prime are clearly indicated. Comparing the evolution of element-based versus more holistic representations in relation to this will add to our growing understanding of the developmental trajectory of perceptual grouping processes.
General Discussion
The goal of these experiments was to investigate developmental changes in the ability to structure disparate elements of visual input into global units. We investigated sensitivity to global and local information in hierarchical visual displays in individuals across a wide age range (8 to 30 years) using compound letter stimuli (Navon, 1977) and using a microgenetic approach to map the evolution of both the local and global aspects of the visual representation (Kimchi, 1998). Our results suggest that visual perception is biased to encode and represent local elements in visual scenes even in adolescence.
This is not to say that children and adolescents are not sensitive to global information. In fact, they were highly accurate when identifying the global letter in the compound letter stimuli and when making similarity judgments about probe stimuli that shared a global configuration with the prime in the microgenetic experiment. However, the slower reaction times in response to the global information in both experiments for both children and adolescents indicates that the global properties are considerably more weakly represented than is the local information. In other words, local precedence appears to dominate perceptual organization until adolescence, when a gradual transition begins for visual perception to become organized around more global or configural aspects of the input. This finding was evident across both paradigms, regardless of whether the stimuli included letters or shapes, were presented for brief or long durations, or contained few or many local elements that comprised the global shapes. These findings are consistent with several other studies reporting the late development of global processing (Burack et al., 2000; Dukette & Stiles, 1996; Enns et al., 2000; Kimchi et al., 2005; Mottron et al., 1999, 2003; Poporino et al., 2004).
It is important to note that these findings are not attributable to gender differences in perceptual organization or age differences in the use of attentional strategies. Our analyses with males resulted in the same pattern of developmental differences in both tasks and two recent studies of perceptual organization in adults using similar tasks failed to find gender differences in perceptual processing biases (Kimchi, Sulitzeanu-Kenan, & Amishav, in preparation; Müller-Oehring, Schulte, Raassi, Pfefferbaum, & Sullivan, 2007). Also, that we get the same pattern of results, namely that children and adolescents are much more inclined to initially encode individual local elements despite being quite accurate to identify local and global structure in a visual stimulus, across two different tasks that vary in attentional demands, suggests that the developmental differences are not a function of the different strategies for the task at hand.
In many ways, our findings characterize the strong biases in the way the visuoperceptual system becomes organized for processing stimuli with multiple levels of structure. If we assume that an important goal of perceptual organization is to facilitate object recognition and identification, it makes sense that these aspects of perceptual organization will encode the most reliable cues for object recognition and identification, which include global shape. Our results suggest that the automation of these processes takes a very long time to develop ontogenetically. This is not to say that adults could not “choose” to attend to a local detail of these visual stimuli, but that their default strategy (which is optimal in terms of real world recognition) is to derive a coherent representation of the whole stimulus. This is not the case for the other two age groups.
These results appear to be somewhat inconsistent with those of Kimchi and colleagues (Kimchi et al., 2005), who found an early maturation of perceptual grouping for many, small element displays using a visual search paradigm. Contrary to these findings, we found evidence for a more protracted developmental trajectory for the encoding of global configuration of hierarchical displays with many, small elements using the microgenetic priming paradigm. At the shortest prime duration, children were much faster to identify the probes that shared local elements with the prime, adolescents were equally fast at identifying the probes that shared a global configuration or local elements with the prime, and adults were faster at identifying probes that shared a global configuration with the prime. These findings indicate that the entry-level units of the visual representation of many-element stimuli are dominated by information about local elements in children, by the global configuration in adults, and by both kinds of information in adolescents.
There are, however, important methodological differences between the two studies. Kimchi and colleagues used a visual search paradigm in which the target and distracters were presented simultaneously and remained present until participants responded or until 7s elapsed for the children or 3s for the adults. Spatial proximity among the local elements within an item and spatial distance between the target and distracter items may have facilitated perceptual grouping for both the few- and many-element items, especially with the long exposure duration, leading to apparently different conclusions about the developmental trajectory of perceptual grouping abilities. Other studies support this interpretation and suggest that proximity facilitates perceptual grouping in visual search tasks for children (Burack et al., 2000; Enns et al., 2000; Hadad & Kimchi, 2006). However, this interpretation is insufficient for explaining the consistency across the two studies in responses to the few-element stimuli and the incongruent responses to the many-element stimuli. Children were biased to group the many-element stimuli in the visual search study, but not in the microgenetic study.
The procedural difference between the microgenetic and visual search paradigms in the temporal presentation of the stimuli may have important implications for the kinds of perceptual organization processes that were accessed in these paradigms. Kimchi and Navon (2000) argued that different perceptual processes are involved under conditions when stimuli are presented simultaneously versus when they are compared with a memory representation in that simultaneous comparison involves a relative judgment or a perception of contrasts, whereas sequential presentation invokes an absolute judgment or identification. This distinction is crucially relevant to the differences between the visual search and microgenetic paradigms.
The simultaneous presentation of the targets and distracters in the visual search paradigm may have minimized the need for absolute identification of the global shape of each item, instead a coarse representation of the global configuration may have been sufficient to support performance. However, in the microgenetic paradigm, a more precise representation of the local relations must be derived to facilitate responses to the probe stimuli that share the global shape with the prime and/or to interfere with responses to the probe stimuli that differ in global shape from the prime. Thus, a more rudimentary representation of the global configuration might have supported the visual search performance, whereas a more integrated shape representation is needed for the primed matching paradigm. It is this integrated shape that is not available to children and only partially available for adolescents. Together, the results from these two experiments suggest that children and adolescents are capable of grouping many small elements to a certain degree, which may support some global information and figural perception, but the full process of garnering shape information from perceptual grouping to the extent of facilitating shape identification appears to develop late into adolescence.
These results are consistent with the notion that there are multiple processes involved in perceptual organization that develop along different trajectories (Behrmann & Kimchi, 2003; Kimchi et al., 2005). Previous studies with adults have contrasted element clustering and shape formation as two different kinds of perceptual grouping (Razpurker-Apfeld & Kimchi, 2007; Trick & Enns, 1997; see also Koffka, 1935). Element clustering involves determining which elements belong together whereas shape formation involves determining cluster boundaries. The process of element clustering is facilitated when the number of elements increases and their relative size decreases, both for adults (Bacon & Egeth, 1991; Banks & Prinzmetal, 1976; Kimchi, 1998) and children (Dukette & Stiles, 1996, 2001; Kimchi et al., 2005; Plaisted, Dobler, Bell, & Davis, 2006). Taken together, the visual search results (Kimchi et al., 2005) and the present microgenetic results suggest that element clustering develops fairly early, and shape formation develops into late adolescence.
This interpretation may also help reconcile our results with existing studies that have reported an early sensitivity to global information in infants and children (Freeseman et al., 1993; Frick et al., 2000; Ghim & Eimas, 1988; Quinn et al., 1993). We suggest that infants’ sensitivity to global information may reflect perceptual organization related to element clustering and that the precedence for global shape in the visuoperceptual representation develops late ontogenetically. However, there are other possible explanations that may help reconcile results from the infant literature with the current findings of late development of global shape processing. In several of the infant studies, infants less than six months of age were not able to discriminate Navon-like stimuli in which the global configuration was the same but the local elements differed, even though they could discriminate these stimuli when the local elements and the global configuration differed (e.g., Freeseman et al., 1993; Ghim & Eimas, 1988). One possible contribution to this bias for global information in infancy is the limitation in visual acuity and high spatial frequency information, both of which are essential for perceiving individual local elements. Even at 6-months of age, infants’ acuity is approximately 20/200 compared to an average adult at 20/20 and they are only able to process low spatial frequencies at approximately 3 cycles/degree compared to an adult who can processing visual information at 30 cycles/degree (for review see Atkinson, 2000). It is not until about age 3 that children exhibit mature visual acuity and spatial frequency processing.
This developmental limitation in the visual system may help explain what appears to be a U-shaped pattern of development for processing global information. The poor acuity and inability to processes high spatial frequency information in early infancy limits the ability to perceive local elements, but the perceivable low spatial frequency information is sufficient to encode a crude representation of the global form. As the visual system matures, the local information begins to dominate the perception of visual stimuli; however, the complicated process of integrating disparate local elements in the service of perceiving a specific global shape requires the integration of many parts of the visual brain, resulting in a protracted developmental trajectory. Additional studies evaluating the specificity of global representations in infants and the relation between acuity/spatial frequency measures and global form perception will be essential to evaluate this hypothesis.
In conclusion, our findings are consistent with the notion that the development of the ability to configure elements in the service of perceptual organization of shape follows along a local - global integrative dimension (Kovács, 2000). This appears to be especially true for the entry-level units of the visual representation of a visual scene. In other words, the ability to integrate the local details of a visual scene into coherent shapes follows a long developmental trajectory through adolescence. This developmental timeline for adult-like perceptual organization processes is coincident with what is known about the structural and functional development of the ventral visual pathway (Bachevalier, Hagger, & Mishkin, 1991; Gogtay, Giedd, Lusk, Hayashi, Greenstein, Vaituzis, et al., 2004; Scherf, Behrmann, Humphreys, & Luna, 2007), which is implicated in these kinds of visuoperceptual processes (Sasaki, 2007).
Acknowledgments
The research reported in this paper was supported by NIH grants NICHD/NIDCD PO1/U19 to Marlene Behrmann and Bea Luna (PI: Nancy Minshew), which is part of the NICHD/NIDCD Collaborative Programs for Excellence in Autism, and T32 HD049354 to Ron Dahl and Robert Noll, as well as a post-doctoral fellowship from the National Alliance for Autism Research to Suzy Scherf and Beatriz Luna. We are grateful to the work of the staff in the CPEA for their help recruiting participants for this project and to our study families for making this research possible.
Contributor Information
K. Suzanne Scherf, Department of Psychology, Carnegie Mellon University.
Marlene Behrmann, Department of Psychology, Carnegie Mellon University.
Ruth Kimchi, Department of Psychology, University of Haifa.
Beatriz Luna, Departments of Psychiatry and Psychology, University of Pittsburgh.
References
- Atkinson J. The developing visual brain. Oxford University Press; New York: 2000. [Google Scholar]
- Bachevalier J, Hagger C, Mishkin M. In: Lassen NA, Ingvar DH, Raichle ME, Friberg L, editors. Functional maturation of the occipitotemporal pathway in infant rhesus monkeys; Alfred Benzon Symposium 31, Brain work and mental activity, quantitative studies with radioactive tracers; Copenhagen: Munksgaard. 1991.pp. 231–240. [Google Scholar]
- Bacon WF, Egeth HE. Local processes in preattentive feature detection. Journal of Experimental Psychology: Human Perception & Performance. 1991;17:77–90. doi: 10.1037//0096-1523.17.1.77. [DOI] [PubMed] [Google Scholar]
- Banks WP, Prinzmetal W. Configurational effects in visual information processing. Perception & Psychophysics. 1976;19:361–367. [Google Scholar]
- Behrmann M, Kimchi R. What does visual agnosia tell us about perceptual organization and its relationship to object perception? Journal of Experimental Psychology: Human Perception & Performance. 2003;29:19–42. doi: 10.1037//0096-1523.29.1.19. [DOI] [PubMed] [Google Scholar]
- Behrmann M, Avidan G, Leonard GL, Kimchi R, Luna B, Humphreys K, et al. Configural processing in autism and its relationship to face processing. Neuropsychologia. 2006;44(1):110–129. doi: 10.1016/j.neuropsychologia.2005.04.002. [DOI] [PubMed] [Google Scholar]
- Beller HK. Priming: Effects of advance information on matching. Journal of Experimental Psychology. 1971;87:176–182. doi: 10.1037/h0030553. [DOI] [PubMed] [Google Scholar]
- Burack JA, Enns JT, Iarocci G, Randolph B. Age differences in visual search for compound patterns: long-versus short-range grouping. Developmental Psychology. 2000;36(6):731–740. [PubMed] [Google Scholar]
- Cassia VM, Simion F, Milani I, Umilta C. Dominance of global visual properties at birth. Journal of Experimental Psychology: General. 2002;131(3):398–411. [PubMed] [Google Scholar]
- Dukette D, Stiles J. Children’s analysis of hierarchical patterns: evidence from a similarity judgment task. Journal of Experimental Child Psychology. 1996;63(1):103–140. doi: 10.1006/jecp.1996.0044. [DOI] [PubMed] [Google Scholar]
- Dukette D, Stiles J. The effects of stimulus density on children’s analysis of hierarchical patterns. Developmental Science. 2001;4(2):233–251. [Google Scholar]
- Enns JT, Burack JA, Iarocci G, Randolph B. The orthogenetic principle in the perception of “forests” and “trees”? Journal of Adult Development. 2004;7(1):41–48. [Google Scholar]
- Farroni T, Valenza E, Simion F, Umilta C. Configural processing at birth: evidence for perceptual organisation. Perception. 2000;29(3):355–372. doi: 10.1068/p2858. [DOI] [PubMed] [Google Scholar]
- Freeseman LJ, Colombo J, Coldren JT. Individual differences in infant visual attention: Four-month-olds’ discrimination and generalization of global and local stimulus properties. Child Development. 1993;64(4):1191–1203. [PubMed] [Google Scholar]
- Frick JE, Colombo J, Allen JR. Temporal sequence of processing at birth: Evidence for pereptual organization. Infancy. 2000;1:375–386. doi: 10.1207/S15327078IN0103_6. [DOI] [PubMed] [Google Scholar]
- Garner WR. Aspects of a stimulus: Features, dimentsions, and configurations. In: Rosch E, Lloyd BB, editors. Cognition and categorization. Erlbaum; Hillsdale, NJ: 1978. pp. 99–133. [Google Scholar]
- Ghim HR, Eimas PD. Global and local processing by 3- and 4-month-old infants. Perception & Psychophysics. 1988;43(2):165–171. doi: 10.3758/bf03214194. [DOI] [PubMed] [Google Scholar]
- Gogtay N, Giedd JN, Lusk L, Hayashi KM, Greenstein D, Vaituzis AC, et al. Dynamic mapping of human cortical development during childhood through early adulthood. Proceedings of the National Academy of Sciences U S A. 2004;101(21):8174–8179. doi: 10.1073/pnas.0402680101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hadad BS, Kimchi R. Developmental trends in utilizing perceptual closure for grouping of shape: effects of spatial proximity and collinearity. Perception & Psychophysics. 2006;68(8):1264–1273. doi: 10.3758/bf03193726. [DOI] [PubMed] [Google Scholar]
- Hoffman JE. Interaction between global and local levels of a form. Journal of Experimental Psychology: Human Perception and Performance. 1980;6:222–234. doi: 10.1037//0096-1523.6.2.222. [DOI] [PubMed] [Google Scholar]
- Johnson SP, Aslin RN. Perception of object unity in two-month-old infants. Developmental Psychology. 1995;31:739–745. [Google Scholar]
- Kellman PJ, Spelke ES, Short KR. Infant perception of object unity from translatory motion in depth and vertical translation. Child Development. 1986;57(1):72–86. [PubMed] [Google Scholar]
- Kimchi R. Primacy of wholistic processing and global/local paradigm: a critical review. Psychological Bulletin. 1992;112(1):24–38. doi: 10.1037/0033-2909.112.1.24. [DOI] [PubMed] [Google Scholar]
- Kimchi R. Uniform connectedness and grouping in the perceptual organization of hierarchical patterns. Journal of Experimental Psychology: Human Perception and Performance. 1998;24(4):1105–1118. doi: 10.1037//0096-1523.24.4.1105. [DOI] [PubMed] [Google Scholar]
- Kimchi R, Gopher D, Rubin Y, Raij D. Performance under dichoptic versus binocular viewing conditions: effects of attention and task requirements. Human Factors. 1993;35(1):35–55. doi: 10.1177/001872089303500103. [DOI] [PubMed] [Google Scholar]
- Kimchi R, Hadad B, Behrmann M, Palmer SE. Microgenesis and ontogenesis of perceptual organization: evidence from global and local processing of hierarchical patterns. Psychological Science. 2005;16(4):282–290. doi: 10.1111/j.0956-7976.2005.01529.x. [DOI] [PubMed] [Google Scholar]
- Kimchi R, Sulitzeanu-Kenan A, Amishav R. Gender differences in discrimination of local orientation. in preparation. [DOI] [PubMed] [Google Scholar]
- Koffka K. Principles of Gestalt psychology. Harcourt, Brace & World; New York: 1935. [Google Scholar]
- Kovacs I. Human development of perceptual organization. Vision Research. 2000;40(10-12):1301–1310. doi: 10.1016/s0042-6989(00)00055-9. [DOI] [PubMed] [Google Scholar]
- Kovacs I, Kozma P, Feher A, Benedek G. Late maturation of visual spatial integration in humans. Proceedings of the National Academy of Sciences U S A. 1999;96(21):12204–12209. doi: 10.1073/pnas.96.21.12204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kozma P, Kovacs I, Benedek G. Normal and abnormal development of visual functions in children. Acta Biologia Szegediensis. 2001;45(1):23–42. [Google Scholar]
- Martin M. Local and global processing: The role of sparsity. Memory & Cognition. 1979;7:476–484. [Google Scholar]
- Mondloch CJ, Geldart S, Maurer D, de Schonen S. Developmental changes in the processing of hierarchical shapes continue into adolescence. Journal of Experimental Child Psychology. 2003;84(1):20–40. doi: 10.1016/s0022-0965(02)00161-3. [DOI] [PubMed] [Google Scholar]
- Mottron L, Burack JA, Stauder JE, Robaey P. Perceptual processing among high-functioning persons with autism. Journal of Child Psychology and Psychiatry. 1999;40(2):203–211. [PubMed] [Google Scholar]
- Müller-Oehring EM, Schule T, Raassi C, Pfefferbaum A, Sullivan EV. Local-global interference is modulated by age, sex, and anterior corpus callosum size. Brain Research. 2007;1142:189–205. doi: 10.1016/j.brainres.2007.01.062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Navon D. Forest before trees: The precedence of global features in visual perception. Cognitive Psychology. 1977;9:353–383. [Google Scholar]
- Navon D. How many trees does it take to make a forest? Perception. 1983;12(3):239–254. doi: 10.1068/p120239. [DOI] [PubMed] [Google Scholar]
- Navon D. What does a compound letter tell the psychologist’s mind? Acta Psychologica. 2003;114(3):273–309. doi: 10.1016/j.actpsy.2003.06.002. [DOI] [PubMed] [Google Scholar]
- Ozonoff S, Strayer DL, McMahon WM, Filloux F. Executive function abilities in autism and Tourette syndrome: an information processing approach. Journal of Child Psychology and Psychiatry. 1994;35(6):1015–1032. doi: 10.1111/j.1469-7610.1994.tb01807.x. [DOI] [PubMed] [Google Scholar]
- Paquet L, Merikle PM. Global precedence: The effect of exposure duration. Canadian Journal of Psychology. 1984;38:45–53. [Google Scholar]
- Plaisted K, Swettenham J, Rees L. Children with autism show local precedence in a divided attention task and global precedence in a selective attention task. Journal of Child Psychology and Psychiatry. 1999;40(5):733–742. [PubMed] [Google Scholar]
- Plaisted K, Dobler V, Bell S, Davis G. The microgenesis of global perception in autism. Journal of Autism and Developmental Disorders. 2006;36(1):107–116. doi: 10.1007/s10803-005-0047-0. [DOI] [PubMed] [Google Scholar]
- Porporino M, Shore DI, Iarocci G, Burack JA. A developmental change in selective attention and global form perception. International Journal of Behavioral Development. 2004;28(4):358–364. [Google Scholar]
- Quinn PC, Burke S, Rush A. Part-whole perception in early infancy: Evidence for perceptual grouping produced by lightness similarity. Infant Behavior & Development. 1993;16:9–42. [Google Scholar]
- Quinn PC, Bhatt RS. Are some gestalt principles deployed more readily than others during early development? The case of lightness versus form similarity. Journal of Experimental Psychology: Human Perception and Performance. 2006;32(5):1221–1230. doi: 10.1037/0096-1523.32.5.1221. [DOI] [PubMed] [Google Scholar]
- Quinn PC, Bhatt RS, Brush D, Grimes A, Sharpnack H. Development of form similarity as a Gestalt grouping principle in infancy. Psychological Science. 2002;13(4):320–328. doi: 10.1111/1467-9280.00459. [DOI] [PubMed] [Google Scholar]
- Quinn PC, Eimas PD. Pattern-line effects and units of visual processing in infants. Infant Behavior & Development. 1986;9(11):57–70. [Google Scholar]
- Razpurker-Apfeld I, Kimchi R. The time course of perceptual grouping: The role of segregation and shape formation. Perception & Psychophysics. 2007;69(5):732–743. doi: 10.3758/bf03193775. [DOI] [PubMed] [Google Scholar]
- Rinehart NJ, Bradshaw JL, Moss SA, Brereton AV, Tonge BJ. Atypical interference of local detail on global processing in high-functioning autism and Asperger’s disorder. Journal of Child Psychology and Psychiatry. 2000;41(6):769–778. [PubMed] [Google Scholar]
- Rock I. The description and analysis of object and event perception. In: Boff KR, Kaufman L, Thomas JP, editors. Handbook of perception and human performance. Vol. 2. Wiley; New York: 1986. pp. 33-1–33-71. [Google Scholar]
- Sasaki Y. Processing local signals into global patterns. Current Opinion in Neurobiology. 2007;17(2):132–139. doi: 10.1016/j.conb.2007.03.003. [DOI] [PubMed] [Google Scholar]
- Scherf KS, Behrmann M, Humphreys K, Luna B. Visual category-selectivity for faces, places and objects emerges along different developmental trajectories. Developmental Science. 2007;10(4):F15–30. doi: 10.1111/j.1467-7687.2007.00595.x. [DOI] [PubMed] [Google Scholar]
- Schneider W, Eschman A, Zuccolotto A. E-Prime User’s Guide. Psychology Tools, Inc; Pittsburgh: 2001. [Google Scholar]
- Sekuler AB, Palmer SE. Perception of partly occluded objects: A microgenetic analysis. Journal of Experimental Psychology: General. 1992;11:95–111. [Google Scholar]
- Shepp BE, Schwartz KB. Selective attention and the processing of integral and nonintegral dimensions: a developmental study. Journal of Experimental Child Psychology. 1976;22:73–85. doi: 10.1016/0022-0965(76)90091-6. [DOI] [PubMed] [Google Scholar]
- Smith JD, Kemler DG. Developmental trends in free classification: Evidence for a new conceptualization of perceptual development. Journal of Experimental Child Psychology. 1977;22:279–298. [Google Scholar]
- Trick LM, Enns JT. Clusters precede shapes in perceptual organization. Psychological Science. 1997;8(2):124–129. [Google Scholar]