

Abstract
Foraging choices in tufted capuchins monkeys are guided by perceptual, cognitive, and motivational factors, but only little is known about how these factors might interact. The present study investigates how different types of sensory information affect capuchins’ ability to locate hidden food. In two experiments, capuchins were presented with two cups, one baited and one empty. Monkeys were given visual, acoustic, or acoustic-visual information related to the baited cup, the empty cup, or both baited and empty cup. Results show that capuchins spontaneously used visual information to locate food, and that information indicating presence and absence of food led to higher success rates than information indicating only absence of food. In contrast, acoustic information did not lead to success rates above chance levels and failed to enhance performance in combination with visual information. Capuchins spontaneously avoided a visually empty cup, but they did not appear to associate sounds with either the presence or absence of food. Being able to locate food items with the aid of acoustic cues might be a learned process that requires interactive experiences with the task’s contingencies.
Keywords: Foraging choices, visual information, acoustic information, capuchin monkey, inferential reasoning
Similar to that of many other primates, the diet of capuchin monkeys (genus Cebus) is made up of mostly fruits and insects. What seems to set capuchin monkeys apart is the vast variety of plants and insects consumed, with over 90 families and over 600 species at a recent count (Fragaszy, Visalberghi, & Fedigan, 2004). While many of these food items are easily accessible, others are protected by hard shells, tree bark, spines, or thorns. Nonetheless, capuchins are able to gain access to these food items by way of their manipulative endeavor, often pounding, biting, rubbing, or otherwise breaking into defenses. These resourceful foraging techniques have deservedly earned them the description “extractive foragers”, if not “destructive foragers” (Fragaszy et al., 2004).
Doubtlessly, extractive foraging can be an energetic activity caused by an interplay of perceptual, cognitive, and motivational processes, and capuchins might engage in it only when they can expect a reasonable rate of return. Information from different sensory modalities might be utilized for this decision process. For example, capuchins in the wild have been reported to engage in ‘tap-scanning’ whereby an individual first scans the stem of a bamboo plant, taps it lightly with its fingertips, and then rips it open to retrieve an insect (Phillips, Grafton, & Haas, 2003). It seems likely that monkeys first use visual information to determine the potential presence of insects, and then generate acoustic information for confirmation through such tapping behavior.
Phillips, Shauver Goodchild, Haas, Ulyan, and Petro (2004) investigated what effect information from different sensory modalities has on foraging decisions under controlled laboratory conditions. Capuchins were presented with a foraging log that had worms hidden inside one of several cavities. Monkeys were able to obtain visual, acoustic, or olfactory information about the worms’ location, or combinations of information from two or all three sensory modalities. Overall, the frequency of monkeys’ manipulations of the log increased with increasing sensory information, suggesting that the amount of available sensory information directly affects monkeys’ motivation to engage in foraging behaviors.
In addition to motivational factors, one might also ask what cognitive processes are involved in successfully locating hidden food items. To evaluate their chances of successful food retrieval, capuchins might consider not only where food is available, but also where food is not available. That is, capuchins might infer the location of food items with the aid of positive cues (see, hear, or smell food item) or negative cues (failure to see, hear or smell food items). The ability to make decisions based on negative cues (or information of absence) has been called inferential or deductive reasoning, and is a prominent area of comparative cognitive investigations. For example, Call (2004) investigated inferential reasoning abilities in all 4 great ape species. Apes were presented with two cups, one empty and one baited, and were provided with visual information about one or both cups. Several individuals chose the correct cup significantly above chance in all conditions, meaning they selected the baited cup even when they had received information about only the empty cup. A recent study by Bräuer, Kaminski, Riedel, Call, and Tomasello (2006) with a similar set up confirmed these findings in chimpanzees and bonobos. Both species selected a baited location based on negative visual cues about a second location. The same study revealed that domestic dogs fail to infer the correct location based on the purely physical visual cues, but that their performance improves when the cues are provided by a human experimenter.
Capuchin’s visual reasoning abilities have also been investigated. In an early experiment on inference of food locations, Grether and Maslow (1937) presented four capuchins with two opaque cups, one baited and the other empty. An experimenter showed monkeys the empty cup and then made both cups available to the monkeys. Subjects easily chose the unseen, baited cup significantly above chance levels. It appears that one capuchin also successfully transferred to more complex 3- and 4-cup problems. More recently, Sabbatini and Visalberghi (2008) provided capuchin monkeys with two boxes, one baited and one empty, and gave the monkeys visual information about one or both boxes. Four out of four monkeys were able to select the baited box based on negative information, suggesting that inferential reasoning with the aid of visual cues might be widespread among capuchins.
The degree to which acoustic information can help individuals to find hidden food items has been investigated to a lesser degree. Naturally, animals that hunt might be very finely attuned to the sounds of their prey (e.g. marsh hawks to the sound of voles; Rice, 1982), and in return, animals that are preyed upon might be very finely attuned to the sounds of their predators (e.g. California ground squirrels to the sound of rattle snakes; Swaisgood, Rowe, & Owings, 1999). But can animals that are predominantly frugivores or folivores use acoustic information in inferential reasoning tasks? Call (2004) and Bräuer et al. (2006) found that great apes can successfully select a baited container based on positive or negative acoustic information. Dogs on the other hand appear to be attracted by food locations that emit sounds regardless of whether the sounds are causally or arbitrarily related to the presence of food (Bräuer et al., 2006). As for capuchins, Sabbatini and Visalberghi (2008) found that two out of four monkeys tested were able to select a baited box when a second box revealed a negative acoustic cue (noiselessly shaken).
To provide a fuller picture of what types of information capuchin monkeys use to locate food items, the following experiments were designed to evaluate whether capuchins utilize visual, acoustic, or both visual and acoustic information. In particular, we tested whether the combination of both visual and acoustic information might increase their success rate of selecting the correct cup when compared to information from only one sensory modality (visual or acoustic). In Experiment 1, monkeys were presented with two cups, one baited and the other empty. We varied the amount and type of information monkeys received by revealing where the food item was located (positive cue), where the food item was not located (negative cue), or both where the food item was and was not located (positive and negative cue). In the visual condition, monkeys were shown the contents of cups. In the acoustic condition, cups were shaken and for half the monkeys, the baited cup made a noise; for the other half, the empty cup made a noise. This arrangement allowed us to further investigate whether capuchins might have a natural propensity to associate positive acoustic cues with the presence rather than the absence of food items. Finally, in the acoustic-visual condition, monkeys were first given acoustic information (consistent with the information received in acoustic-only trials) and then visual information about the food’s location. We hypothesized that monkeys would select the correct food location in all three conditions.
Experiment 1
Subjects
Subjects were 19 tufted capuchin monkeys (Cebus apella), 6 adult males and 6 adult females (5+ years old) as well as 6 juvenile males and 1 juvenile female (2-4 years old; see Table 1). All animals were captive born and housed at the National Institute of Health Animal Center in Maryland, USA. Eleven animals were housed in small groups of 2-4 animals in cages (1.78m × 0.76m × 1.85m); the remaining 8 animals were part of a larger breeding group of capuchins that were housed in indoor runs during the winter (3 connected runs, each 2.3m × 4.1m × 2.1m) and in a large outdoor enclosure during the summer (ca. 250 m3). Monkeys were not food-deprived and received daily supplements of seeds and fresh fruits or nuts. Water and commercial monkey biscuits were available ad libitum.
Table 1.
Age class, sex, housing, and experiment participation for each subject. 1 = acoustic congruent cue in Exp. 1, 2 = acoustic incongruent cue in Exp. 1, 3 = acoustic congruent cue in Exp. 2, 4 = acoustic incongruent cue in Exp. 2, 5 = participated in Exp. 3
| Name | Sex | Age class | Housing | Experiment participation and conditions |
|---|---|---|---|---|
| Garth | Male | Adult | Group | 1, 4, 5 |
| HotRod | Male | Juvenile | Group | 1, 4, 5 |
| Mr. Goodbar | Male | Juvenile | Group | 1, 4, 5 |
| Shane | Male | Juvenile | Group | 1, 4, 5 |
| Liv | Female | Adult | Group | 1, 4, 5 |
| Ivory | Female | Adult | Group | 1, 4, 5 |
| Lychee | Female | Adult | Group | 2, 3, 5 |
| Snickers | Female | Juvenile | Group | 2, 3, 5 |
| Alou | Male | Juvenile | Cage | 2, 3, 5 |
| DiMaggio | Male | Juvenile | Cage | 2, 3, 5 |
| Larry | Male | Adult | Cage | 2, 3 |
| Loco | Male | Adult | Cage | 2, 3 |
| Icabod | Male | Adult | Cage | 2, 3 |
| Lee | Female | Adult | Cage | 1 |
| Lorena | Female | Adult | Cage | 1 |
| Lyla | Female | Adult | Cage | 1 |
| Ian | Male | Adult | Cage | 2 |
| Sayla | Male | Adult | Cage | 2 |
| Hugo | Male | Juvenile | Cage | 2 |
Procedure
Caged-housed animals were separated from other animals during testing. For group-housed animals, the experimenter set up the apparatus in a quiet part of the enclosure and waited for the target animal to approach.
The experimental set up consisted of two white polystyrene foam cups (9 cm diameter and 11.5 cm in height) placed onto two white plastic plates (10 cm × 10 cm). Before each trial, the experimenter baited one of the cups behind a white screen (60 cm × 30 cm) by placing a grape underneath it. The empty cup was also handled so as to control for any visual or acoustic cues.
We tested the monkeys in 4 different conditions (see also Table 2):
Table 2.
Summary of conditions and cues used in Experiment 1.
| Visual condition | Acoustic condition | Acoustic-visual condition | Control | |
|---|---|---|---|---|
| Positive cue | The baited cup is lifted to reveal its contents, then returned to the starting position. The empty cup is touched. | The baited cup is shaken twice, then returned to the starting position. The empty cup is touched. | The baited cup is shaken twice, and returned to its starting position. Then the baited cup is lifted to reveal its contents, and returned to the starting position. The empty cup is touched. | --- |
| Negative cue | The empty cup is lifted to reveal its contents, then returned to the starting position. The baited cup is touched. | The empty cup is shaken twice, then returned to the starting position. The baited cup is touched. | The empty cup is shaken twice, and returned to its starting position. Then the empty cup is lifted to reveal its contents, and returned to the starting position. The baited cup is touched. | --- |
| Positive and negative cue* | The baited cup is lifted to reveal its contents, then returned to the starting position. The empty cup is lifted to reveal its contents, then returned to the starting position. | The baited cup is shaken twice, then returned to the starting position. The empty cup is shaken twice, then returned to the starting position. | The baited cup is shaken twice, and its contents are revealed. Then the empty cup shaken twice, and its contents are revealed. | --- |
| Control | --- | --- | --- | No cues are given |
In half the trials, the positive cue was given first; in the other half, the negative cue was given first.
visual: the experimenter lifted one cup so that its contents were visible on the plate, then returned the cup to the starting position.
acoustic: the experimenter shook one cup twice, then returned it to the starting position.
acoustic-visual: the experimenter first shook one cup twice and returned it to the starting position. The same cup was then lifted so that its contents were visible on the plate, and returned to its starting position.
control: cups were not manipulated.
In addition, we varied the amount of information the monkey received during each trial (see also Table 2):
positive: the cue revealed the location of the bait. The empty cup was briefly touched after the cue, but information about its contents were not revealed
negative: the cue revealed an empty cup. The baited cup was briefly touched after the cue, but information about its contents were not revealed
positive and negative: the cue revealed the location of the empty and the baited cup. For half the trials, the baited cup was shown first; for the other half, the empty cup was shown first
Ten monkeys were selected to receive congruent acoustic information in the acoustic and acoustic-visual condition. For these monkeys, a small metal disk (3 cm diameter) was always placed inside the baited cup in addition to the food reward, and the empty cup was filled with white cotton fabric. Shaking of the baited cup therefore produced an audible sound. The remaining 9 monkeys received incongruent acoustic information. For these monkeys, the metal disk was always placed into the empty cup, and the baited cup containing the food reward was filled with white cotton fabric. Shaking of the empty, but not the baited cup therefore produced an audible sound. Both cups were placed upside down onto the plastic plates, and the screen was removed. Each trial started when the subject was oriented towards the experimenter.
Each cue was presented ca. 60 cm in front of the monkey. After each cue, both cups were slid towards the monkey to a distance of ca. 30 cm. Monkeys could chose one cup by reaching towards it or touching/lifting it. If the baited cup was chosen, the experimenter picked up the grape, gave it to the monkey, and retrieved the cups to set up the next trial. If the empty cup was chosen, the monkey received no reward; the experimenter retrieved the cups and set up the next trial. If no cup was chosen after 30 sec, the experimenter replaced the screen and started the next trial. Monkeys received 48 trials in each condition, 12 trials per day. Trial order and location of the baited cup (left-right) was randomized for each monkey. If a monkey did not select a cup for 3 trials in a row, or failed to come forward in the group setting, we terminated the session and continued testing the next day.
Results
Visual Condition
In the visual condition, monkeys selected the baited cup significantly above chance levels (control condition, t (18) = 11.48, p<0.001; see Fig. 1). A repeated measure ANOVA indicated that the type of cue monkeys received affected their performance (F (2, 36) = 3.91, p=0.029). Further post-hoc comparisons with Bonferroni correction revealed that monkeys performed better when receiving both positive and negative cues (mean=13.42) than when receiving only negative cues (mean=11.63, p=0.050). No difference was found between positive cues (mean=12.89) and negative cues (p>0.05). At an individual level, 17 monkeys selected the correct cup significantly above chance levels (50% correct; binomial test, p<0.05).
Figure 1.
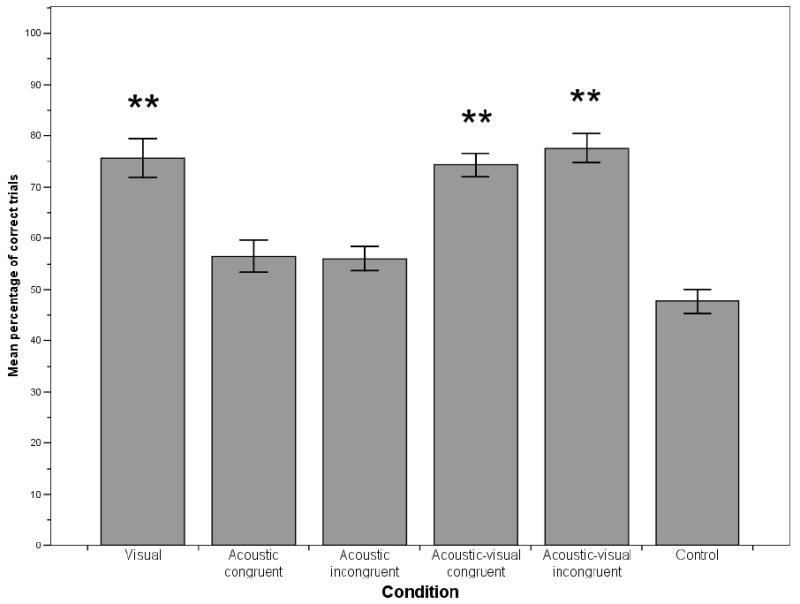
Experiment 1: Percentage of correct trials in each condition. Error bars represent standard errors, asterisk indicates significant difference to control condition with p<0.001.
Acoustic condition
Since monkeys received either acoustic congruent or acoustic incongruent information, we split monkeys into two groups and analyzed data separately for each group. Neither group selected the baited cup significantly above chance levels (control condition; t (8) = 2.06, p>0.05 for congruent and t (9) = 1.15. p>0.05 for incongruent; see Fig. 1). Furthermore, receiving congruent information did not significantly enhance performance compared to receiving incongruent information (t (17) = -0.25, p>0.05). At an individual level, 2 monkeys in the congruent group and 1 monkey in the incongruent group were significantly better than chance (50% correct; binomial test, p<0.05).
Acoustic-visual condition
As in the acoustic condition, we split monkeys into two groups depending on whether they received acoustic congruent or acoustic incongruent information. Both groups selected the baited cup significantly above chance levels (control condition; t (8) = 9.31, p<0.001 for congruent and t (9) = 6.94, p<0.001 for incongruent; see Fig. 1). Monkeys that received congruent information did not perform significantly better than monkeys that received incongruent information (t (17) = -1.23, p>0.05). There was no difference in success rate depending on the type of cue received (positive, negative, or both positive and negative; congruent information: F (2, 16) = 1.54, p>0.05; incongruent information: F (2, 18) = 2.38, p>0.05). At an individual level, 9 monkeys in the congruent and 9 monkeys in the incongruent group performed significantly better than chance (50% correct; binomial test, p<0.05).
Discussion
Monkeys successfully selected the baited cup in both the visual and acoustic-visual condition. In the visual condition, monkeys were more successful when receiving both positive and negative cues than when receiving only negative cues, whereas monkeys were equally successful regardless of which cue they received in the acoustic-visual condition. In contrast, monkeys as a group performed at chance levels when they received only acoustic information. Only 3 monkeys performed above chance levels on an individual basis, two monkeys based on congruent and one monkey based on incongruent cues. These results suggest that monkeys do not spontaneously associate sound with the presence of food items, and that they locate food items with the aid of visual and acoustic-visual information, but not with acoustic information alone.
The fact that monkeys failed to select the baited cup above chance levels in the acoustic condition raises questions about whether they were able to utilize the acoustic information in the acoustic-visual condition. That is, did acoustic information contribute to the monkeys’ success rate, or did acoustic information fail to impact on performance in the acoustic-visual condition? In Experiment 2, we tested whether acoustic information contributed to the capuchins’ selection choice in the acoustic-visual condition. Experiment 2 was identical to Experiment 1 with the only difference that the type of acoustic information (congruent or incongruent information) was reversed in both the acoustic and acoustic-visual condition, i.e. if sound indicated the presence of food in Experiment 1, it now indicated the absence of food and vice versa. We hypothesized that if monkeys used acoustic information in the acoustic-visual condition to select the baited cup, then reversing the significance of this cue would send conflicting information to the monkeys about the bait’s location and diminish their performance.
Experiment 2
Subjects
Subjects were all group-housed animals from Experiment 1, namely 1 adult male, 5 juvenile males, 3 adult females and 1 juvenile female, and 3 caged-housed adult males (see also Table 1).
Procedure
The testing procedure was identical to Experiment 1 with the only difference that subjects who previously received acoustic congruent information now received acoustic incongruent information and vice versa. Subjects received 48 trials in each condition in random order.
Results
Visual condition
Monkeys selected the baited cup significantly above chance levels (control condition; t (12) = 14.55, p<0.001; see Fig. 2). The type of cue received had an effect on the monkeys’ chances of success (F (24, 2) = 3.67, p=0.041). Further post-hoc comparisons with Bonferroni correction showed that monkeys selected the baited cup at higher rates when given both positive and negative cues (mean=15.46) than when given only negative cues (mean=14.15, p=0.051). No differences between positive cues (mean=14.69) and negative cues were found (p>0.05). All 13 subjects performed significantly better than chance on an individual basis (50% correct; binomial test, p<0.05). Subjects also performed significantly better in Experiment 2 compared to Experiment 1 (t (12) = -3.19, p=0.008).
Figure 2.
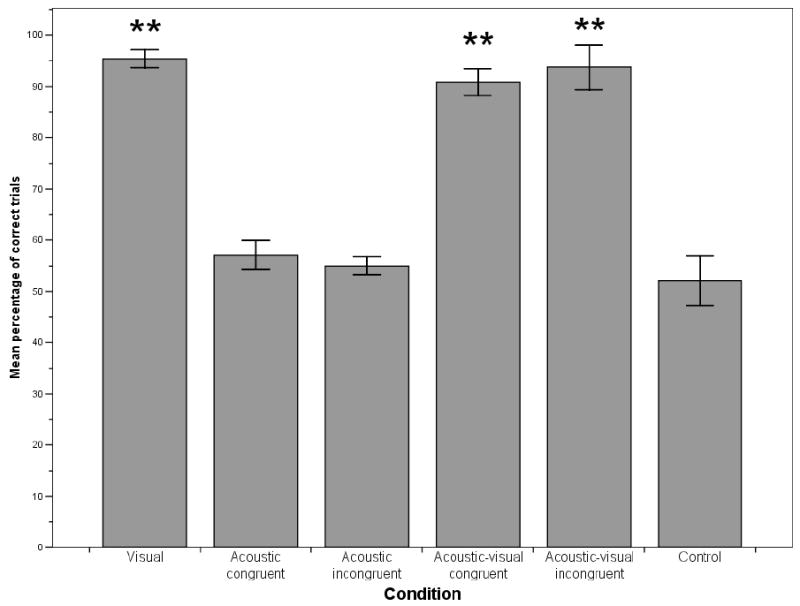
Experiment 2: Percentage of correct trials in each condition. Error bars represent standard errors, asterisk indicates significant difference to control condition with p<0.001.
Acoustic condition
We again split monkeys in two groups depending on whether they received acoustic congruent or incongruent information. Neither group selected the baited cup above chance levels (congruent information: t (6) = 0.95, p>0.05; incongruent information: t (5) = 1.08, p>0.05; see Fig. 2). Monkeys receiving the acoustic congruent cue did not perform significantly better than monkeys receiving the acoustic incongruent cue (t (11) = -0.51, p>0.05). No monkey performed significantly better than chance on an individual basis (50% correct; binomial test, p>0.05). Monkeys did not perform significantly better in Experiment 2 than in Experiment 1 (congruent information: t (6) = 0.07, p>0.05 and incongruent information: t (5) = 1.00, p>0.05).
Acoustic-visual condition
Monkeys who received acoustic congruent and monkeys who received acoustic incongruent information both performed significantly above chance levels (congruent: t (6) = 9.08, p<0.001 and incongruent: t (5) = 9.20, p<0.001; see Fig. 2). Acoustic congruent information did not significantly enhance monkeys’ success rates (t (11) = 0.63, p>0.05). The type of cue given also failed to impact on success rates (congruent: F (2, 12) = 2.96, p>0.05; incongruent: F (2, 10) = 2.73, p>0.05). Individually, all 13 subjects performed significantly better than chance (50% correct; binomial test, p<0.05). Both groups performed significantly better in Experiment 2 than in Experiment 1 (congruent: t (6) = -2.45, p=0.050; incongruent: t (5) = -4.25, p=0.008).
Discussion
As in Experiment 1, monkeys in Experiment 2 selected the baited cup significantly above chance levels in the visual and acoustic-visual condition. Moreover, monkeys performed significantly better in Experiment 2 than in Experiment 1 in both the visual and acoustic-visual condition, but performed at chance levels in the acoustic condition, which suggests that for the largest part, they relied on visual information in the acoustic-visual condition. It also suggests that monkeys improved their performances with increasing number of trials. In the visual condition, monkeys were again more successful when given both positive and negative cues rather than just negative cues. In the acoustic-visual condition, monkeys were equally successful regardless of the type of cue received.
Results from Experiment 1 and Experiment 2 indicate that capuchin monkeys spontaneously use visual, but not acoustic information to find hidden food items. Both experiments found higher success rates for positive and negative cues compared to only negative cues in the visual condition, which suggests that negative information might be more difficult for them to use than positive information. In a previous study, capuchins were presented with opaque tubes, one baited and two others empty, and monkeys could obtain information about the bait’s location by looking into tubes (Paukner, Anderson, & Fujita, 2006). In this situation, capuchins routinely searched tubes until they saw the bait before making a selection, indicating that capuchins may prefer positive visual cues to find hidden food. In the present study, capuchins performed above chance levels when given only negative visual information. Did they select the baited cup because they knew that it was the only possible place to contain a food item, or did they simply avoid an empty cup?
We conducted Experiment 3 to test directly whether capuchins have a tendency to avoid an empty cup. In particular, we tested if monkeys would select a cup they know to be empty below chance levels. In order to avoid any potential learning effects, we rewarded monkeys regardless of whether they selected the empty cup or any other cup.
Experiment 3
Subjects
Ten monkeys were available for Experiment 3, 6 males and 4 females; see Table 1 for details.
Procedure
All subjects were individually tested in a tunnel of cages (0.55 m × 1.55 m × 0.65 m). Using the same screen to cover the baiting process as in Experiments 1 and 2, an experimenter placed three cups (9 cm diameter and 11.5 cm in height) in front of the subject. Two of these cups were baited with a small piece of grape; the third cup was empty. Always starting from the left, the experimenter started each trial by individually lifting each cup. If the cup was baited, the experimenter lifted the cup and replaced it, without revealing its contents, on a platform. If the cup was empty, the experimenter lifted the cup, tilted it to show to the monkey that it was empty, and replaced it on the platform. After all three cups had been lifted and replaced, the experimenter slid the platform towards the monkey, and the monkey could select a cup by reaching towards it or touching it. If the monkey selected a baited cup, it was allowed to retrieve it and eat its contents. If the monkey selected the empty cup, the experimenter immediately retrieved the cup and gave the monkey a piece of grape. All monkeys received one session of 12 trials. Location of the empty cup was counterbalanced and order of trials was randomized for each monkey.
Results
Out of 12 trials, monkeys selected the empty cup on average 1.7 times (st. dev. 1.418). Assuming that chance performance to pick any one cup is 33.33% (or 4 trials out of 12 with 3 cups presented), capuchins selected the empty cup significantly below chance levels (Chi-Square = 19.838, df = 1, p<0.001). Looking at individual performance, 1 monkey never selected the empty cup (binomial test against 0.33, p<0.05), and 5 monkeys selected the empty cup once (binomial test against 0.33, p=0.057).
Discussion
Not surprisingly, there was indeed a strong tendency in our capuchins to avoid a cup that they had seen to be empty. It therefore seems likely that when given negative visual cues, capuchins in Experiments 1 and 2 selected the baited cup because of a tendency to avoid the empty cup.
Inferences of object locations are typically assessed in object permanence tasks, first described by Piaget (1954) as developing over six stages for children. The final stage 6 of object permanence, typically reached by age 15-18 months (Haake & Somerville, 1985), involves the tracking of an invisibly displaced item and inferring its only possible location. Subjects are presented with a desirable item covered by a small container A, which is passed underneath a larger container B. Container A is then retrieved from underneath container B, and shown to be empty. The subject must therefore trace the item’s passage and infer its presence in container B rather than an additional third container C. The difference between object permanence paradigms and the present paradigm is that in the present paradigm, subjects did not witness the hiding process. Nonetheless, there are parallels between both paradigms in that subjects must infer the location of the object/food with the aid of visual cues presented at various potential object locations. With appropriate controls, capuchins fail the stage 6 object permanence task; they do not infer the only possible location of the bait (Natale & Antinucci, 1989). It therefore seems likely that the selection of the correct cup in the present experiment was based on avoidance of the empty cup, and the experiment’s simple set up with only two cups might have made it seem as if they were correctly inferring the food’s location.
GENERAL DISCUSSION
In two experiments, capuchin monkeys selected a baited cup above chance levels with the use of visual information. Furthermore, even though there was no difference in performance when given positive or negative cues, monkeys performed significantly better when given both positive and negative cues rather than just negative cues. In Experiment 3, monkeys were presented with three cups instead of two, and one was shown to be empty. Here, monkeys avoided the empty cup and preferentially selected a cup on which they had no further information. This pattern of behavior suggests that rather than the cognitively complex process of inferring the presence of an item within the baited cup, monkeys could have succeeded in Experiments 1 and 2 simply by avoiding the cup for which they received negative visual information. Since Experiments 1 and 2 presented only two cups to the monkeys, the baited cup was the only alternative to the empty cup that monkeys avoided, which might have led to their high rate of success.
Capuchins were unable to select the correct cup when information was presented acoustically, regardless of whether the cue indicated the presence or absence of the bait. By contrast, apes can select the correct food location with the aid of acoustic information (Call, 2004; Bräuer et al., 2006), but whether they do so based on inferential reasoning or association is debated (Penn & Povinelli, 2007). Capuchins even failed to learn the significance of the acoustic information during the course of the experiment, and they continued to select the baited cup at chance levels. A discrepancy between visual and acoustic information is also apparent in capuchins’ ability to remember visual and acoustic information (Colombo & D’Amato, 1986) and may reflect a generalized impairment in the processing of acoustic information in comparison to visual information.
An accurate understanding of the acoustic cue also requires an understanding of the causal factors between shaking movements and sound. That is, to be able to correctly connect presence of food with a sound, one would need to grasp that objects in a container bounce of the walls of the container when shaken, thereby causing the emanating noise. Capuchins’ ability to engage in causal reasoning has been intensively explored by Visalberghi and colleagues in the context of tool-using skills. When presented with a clear plastic tube containing a food item, capuchins readily attempt to manipulate the tube by inserting various items into it. By careful analysis of what and how they insert items into the tube, Visalberghi and colleagues were able to show that capuchins appear to have little understanding of the effects of the various tools on the food item (e.g. Visalberghi & Trinca, 1989; Visalberghi & Limongelli, 1994; Fragaszy et al., 2004). Fujita, Kuroshima, and Asai (2003) reported that capuchins can identify at least some relevant aspects of tools in a food retrieval task, and that capuchins can understand spatial relationships between a tool and a food item, but not between tool, food item, and other environmental constraints. The present study shows that capuchins do not spontaneously associate a noise emanating from a shaken cup with a food item, suggesting that they do not causally attribute object presence with shaking movements and noise.
That is not to say, however, that capuchins might not have come to associate sounds and presence of food given sufficient experience with such contingencies. For example, capuchins tested by Sabbatini and Visalberghi (2008) became more successful at using auditory cues after they had been given the opportunity to interact with baited objects that produced sounds when being manipulated. Visalberghi and Neel (2003) evaluated whether captive capuchins use sound or weight cues to determine whether a nut is full or empty. When full and empty nuts could only be discriminated by sound, one capuchin finger-tapped nuts before choosing one of two nuts, and she selected the full nut significantly above chance levels. Even in wild capuchins, adults are reported to be twice as successful compared to juveniles at retrieving insects using the ‘tap-scanning’ method (Phillips et al., 2003), which suggests that a certain amount of experience is required to accurately use acoustic information. As for visual information, monkeys also improved their performance between experiments, which is in line with the idea that the underlying cognitive process was based on a chain of association rather than reasoning. It is also worth noting that wild capuchin monkeys are undoubtedly under stronger selective pressures than our captive capuchins. Gaining an edge in foraging decisions by attunement to acoustic information might therefore be crucial to wild capuchins, but perhaps not for our well-provisioned captive capuchins. Given more intensive experience with the task and perhaps increased importance of the task’s consequences, capuchins might have come to associate the acoustic information with food locations.
A second factor that should be considered is that monkeys in the present study were passive observers of the experimenter’s action. That is, monkeys never interacted with the cups themselves, and therefore never had the opportunity to produce the sounds associated with the baited cup. Within their normal housing, our monkeys readily pick up environmental enrichment objects (pieces of wood, plastic containers, metal mirrors etc.) and bang them against the floor, perches or walls of the enclosure, which produces loud noises. However, we do not customarily provide them with objects that make sounds if just shaken, so we assume that their experience of this type is limited. Studies with human infants have shown that the experience of an action can significantly alter the perception of the same action. For example, Sommerville, Woodward and Needham (2005) presented infants at 3 months of age with an actor reaching for an attractive toy with a Velcro-covered mitten (a new, unfamiliar action). Infants who previously had been allowed to pick up objects with the same mitten looked at the actor for longer than infants who had not had an interactive experience with the mitten. While detailed studies of the link between action and perception are waiting to be conducted in a comparative framework, it is worth noting that capuchins tested by Sababtini and Visalberghi (2008) improved their performance after they themselves had interactive experience with objects that produce sounds when manipulated. It therefore seems possible that monkeys might have shown an increased understanding of acoustic cues following hands-on, interactive experiences with sounds and objects.
In summary, the results of the current study suggest that capuchins spontaneously use visual, but not acoustic information to successfully find hidden food items, in line with other studies highlighting the importance of visual information over other types of sensory information (e.g. Boner & Green, 1997). Being able to find food with the aid of visual cues might be based on attraction to baited cups and avoidance of empty cups rather than more cognitively complex processes. The observed discrepancy between visual and acoustic information is possibly linked to the monkeys’ limited experience with acoustic information indicating presence or absence of food items. Future studies should address whether self-generated acoustic information may be a more salient foraging cue than non-self-generated acoustic information as provided by an experimenter in the present study.
Acknowledgments
This research was supported by funds from the Division of Intramural Research, NICHD. We thank Pier Ferrari for critical comments on earlier drafts of this paper.
Footnotes
Publisher's Disclaimer: The following manuscript is the final accepted manuscript. It has not been subjected to the final copyediting, fact-checking, and proofreading required for formal publication. It is not the definitive, publisher-authenticated version. The American Psychological Association and its Council of Editors disclaim any responsibility or liabilities for errors or omissions of this manuscript version, any version derived from this manuscript by NIH, or other third parties. The published version is available at http://www.apa.org/journals/com/
References
- Boner RH, Green SM. Use of olfactory cues in foraging by owl monkeys (Aotus nancymai) and capuchin monkeys (Cebus apella) Journal of Comparative Psychology. 1997;111:152–158. doi: 10.1037/0735-7036.111.2.152. [DOI] [PubMed] [Google Scholar]
- Bräuer J, Kaminski J, Riedel J, Call J, Tomasello M. Making inferences about the location of hidden food: social dog, causal ape. Journal of Comparative Psychology. 2006;120:38–47. doi: 10.1037/0735-7036.120.1.38. [DOI] [PubMed] [Google Scholar]
- Call J. Inferences about the location of food in the great apes (Pan paniscus, Pan troglodytes, Gorilla gorilla, and Pongo pygmaeus) Journal of Comparative Psychology. 2004;118:232–241. doi: 10.1037/0735-7036.118.2.232. [DOI] [PubMed] [Google Scholar]
- Colombo M, D’Amato MR. A comparison of visual and auditory short-term memory in monkeys (Cebus apella) The Quarterly Journal of Experimental Psychology Section B. 1986;38:425–448. [PubMed] [Google Scholar]
- Fragaszy DM, Visalberghi E, Fedigan LM. The complete capuchin. Cambridge: Cambridge University Press; 2004. [Google Scholar]
- Fujita K, Kuroshima H, Asai S. How do tufted capuchin monkeys (Cebus apella) understand causality involved in tool use? Journal of Experimental Psychology: Animal Behavior Processes. 2003;29:233–242. doi: 10.1037/0097-7403.29.3.233. [DOI] [PubMed] [Google Scholar]
- Grether WF, Maslow AH. An experimental study of insight in monkeys. Journal of Comparative Psychology. 1937;24:127–134. [Google Scholar]
- Haake RJ, Somerville SC. Development of logical search skills in infancy. Developmental Psychology. 1985;21:176–187. [Google Scholar]
- Natale F, Antinucci F. Stage 6 object-concept and representation. In: Antinucci F, editor. Cognitive structure and development in nonhuman primates. Hillsdale, NJ: Lawrence Erlbaum associates; 1989. pp. 97–112. [Google Scholar]
- Paukner A, Anderson JR, Fujita K. Redundant food searches by capuchin monkeys (Cebus apella): a failure of metacognition? Animal Cognition. 2006;9:110–117. doi: 10.1007/s10071-005-0007-2. [DOI] [PubMed] [Google Scholar]
- Penn DC, Povinelli DJ. Causal cognition in human and nonhuman animals: a comparative, critical review. Annual Review of Psychology. 2007;58:97–118. doi: 10.1146/annurev.psych.58.110405.085555. [DOI] [PubMed] [Google Scholar]
- Phillips KA, Grafton BW, Haas ME. Tap-scanning for invertebrates by capuchins (Cebus apella) Folia Primatologica. 2003;74:162–164. doi: 10.1159/000070650. [DOI] [PubMed] [Google Scholar]
- Phillips KA, Shauver Goodchild LM, Haas ME, Ulyan MJ, Petro S. Use of visual, acoustic, and olfactory information during embedded invertebrate foraging in brown capuchins (Cebus apella) Journal of Comparative Psychology. 2004;118:200–205. doi: 10.1037/0735-7036.118.2.200. [DOI] [PubMed] [Google Scholar]
- Piaget J. The construction of reality in the child. Routledge; 1954. [Google Scholar]
- Rice WR. Acoustic location of prey by the marsh hawk: adaptation to concealed prey. The Auk. 1982;19:403–413. [Google Scholar]
- Sabbatini G, Visalberghi E. Inferences about the location of food in capuchin monkeys (Cebus apella) in two sensory modalities. Journal of Comparative Psychology. 2008;122:156–166. doi: 10.1037/0735-7036.122.2.156. [DOI] [PubMed] [Google Scholar]
- Sommerville JA, Woodward AL, Needham A. Action experience alters 3-month-old infants’ perception of others’ actions. Cognition. 2005;96:B1–B11. doi: 10.1016/j.cognition.2004.07.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swaisgood RR, Howe MP, Owings DH. Assessment of rattlesnake dangerousness by California ground squirrels: exploitation of cues from rattling sounds. Animal Behavior. 1999;57:1301–1310. doi: 10.1006/anbe.1999.1095. [DOI] [PubMed] [Google Scholar]
- Visalberghi E, Limongelli L. Lack of comprehension of cause-effect relations in tool-using capuchin monkeys (Cebus apella) Journal of Comparative Psychology. 1992;108:15–22. doi: 10.1037/0735-7036.108.1.15. [DOI] [PubMed] [Google Scholar]
- Visalberghi E, Neel C. Tufted capuchins (Cebus apella) use weight and sound to choose between full and empty nuts. Ecological Psychology. 2003;15:215–228. [Google Scholar]
- Visalberghi E, Trinca L. Tool use in capuchin monkeys: distinguishing between performing and understanding. Primates. 1989;30:511–521. [Google Scholar]