

Abstract
Background
Human metapneumovirus (hMPV) is a newly discovered paramyxovirus that causes acute respiratory illness. Despite apparent near-universal exposure during early childhood, immunity is transient.
Methods
An indirect screening ELISA using a recombinant, soluble, fusion (F) glycoprotein derived from hMPV was used to test for anti-F IgG in 1,380 acute and convalescent sera collected from children in Kamphaeng Phet, Thailand
Results
1,376 (99.7%) tested sera showed evidence of prior infection with hMPV. 67 children demonstrated a four-fold or greater rise in titer for an overall re-infection rate of 4.9%. Two children demonstrated evidence of an initial infection. 49 of the 69 new or re-infections occurred in 2000, accounting for 13.2% of all non-flaviviral febrile illnesses in the study population in that year. Of 69 positive cases, 89.9% reported a respiratory symptom compared to 69.2% of tested negative cases (p<.001). All positive specimens were also tested for an increase in titer to RSV F and 27% exhibited a four-fold or greater rise in titer.
Conclusion
These results demonstrate hMPV reinfection causing illness at rates equal to that seen for initial infections. hMPV may represent a more significant impact in older children than previously realized and may be the cause of significant outbreaks.
Keywords: Human metapneumovirus, fusion protein, reinfection, Thai children
INTRODUCTION
Human metapneumovirus (hMPV) is a recently discovered paramyxovirus that causes acute respiratory illness [1], and has been documented worldwide, including in Thailand [2-5]. Serological surveys have demonstrated greater than 90% of those over the age of 5 having antibodies to hMPV [1, 6]. Despite this apparent near-universal exposure during early childhood, reinfection with hMPV occurs, similar to that seen for respiratory syncytial virus (RSV) [7]. The largest epidemiological study to examine cases of reinfection revealed hMPV as the cause of approximately 4.5% of respiratory illness in 984 adult volunteers [8]. In addition, reinfection among children has also been documented in case reports and as incidental findings in larger prospective studies [9-14].
To retrospectively test for viral infection, serological assays can be performed using acute and convalescent patient sera. For hMPV, immunofluorescence assays using hMPV-infected cells [11, 15] and recombinant baculovirus infected cells expressing the fusion (F) protein [16] have been performed, as well as enzyme linked immunosorbent assays (ELISA) using hMPV infected cell lysates [17, 18], lysates from recombinant baculovirus infected cells expressing nucleoprotein (N) [19], purified N from a prokaryotic expression system [20], and a recombinant vesicular stomatitis virus expressing hMPV F [6]. However, to our knowledge, no serologic studies have been done using purified hMPV glycoproteins from a eukaryotic or human cell line system.
The membrane anchored envelope glycoproteins of paramyxoviruses are the principle antigenic determinant to which virus-neutralizing antibodies are directed. The F protein of hMPV is highly conserved amongst viral isolates and is a major immunologic determinant [21, 22]. Purified, soluble F has been produced and is immunogenic in animal models [23, 24]. Expression of hMPV F in a eukaryotic system should produce a closely related antigen to native protein and is the ideal antigen for a sensitive serologic assay. One concern in serologic tests is cross reactivity to related proteins. RSV is the genetically closest related human paramyxovirus to hMPV and currently all human data demonstrate a lack of cross-reactivity between conserved hMPV proteins and RSV antibodies to viral surface proteins [6, 20, 25].
A retrospective study in Bangladesh identified hMPV as the most common respiratory virus in children with cough and a fever based on an increase in titer between acute and convalescent samples in 107 paired sera tested [17]. This has been one of the limited numbers of studies to examine evidence of hMPV infection in older children, with 42 of those tested over the age of 5. Using archived serum specimens from 1,380 febrile children aged 7 to 16 in Thailand, we determined seroprevalence and reinfection rates for hMPV using a purified fusion protein as antigen in indirect ELISA.
SUBJECTS, MATERIALS AND METHODS
Subjects
The Armed Forces Research Institute of Medical Sciences (AFRIMS) and University of Massachusetts Medical School (UMMS) conducted a prospective study of dengue virus transmission and disease in primary school children in Kamphaeng Phet Province, Thailand from 1998 to 2002 [26, 27]. They enrolled children from 12 elementary schools, and performed active case surveillance from 1 June to 15 November each year, concurrent with peak dengue transmission. Village health workers investigated any school absence to determine if the child had a history of fever within seven days of the absence or an oral temperature of 38°C or greater. If the criteria were met, they administered a symptom questionnaire and took a blood sample then and approximately 14 days later. The Walter Reed Institute of Research Human Use Review Committee, the UMMS Human Subjects Committee and the Thai Ethical Review Committee, Ministry of Public Health, Nonthaburi, Thailand approved the protocol under which the samples were collected. The Uniformed Services University of the Health Sciences Institutional Review Board approved the use of the serum samples for this study as an exempt protocol.
Protein Production
LLC-MK2 cells were infected with hMPV virus strain CAN97-83 from the A2 genotype (kindly provided by Kevin Yim, Virion Systems, Inc., Rockville, MD). RNA was extracted using the RNEasy Mini Kit (Qiagen) according to the manufacturer’s instructions and cDNA created. The soluble version of F is a truncate of F protein with an S-peptide tag that lacks the transmembrane domain and was made using cDNA and the following oligonucleotides: 5’-CCGCTCGAGATGTCTTGGAAAGTGGTGATCAT-3’ and 5’-GGATCCCTAGCTGTCCATGTGCTGGCGTTCGAATTTAGCAGCAGCGGTTTCTTTCCCT TTCTCTGCACTGCTTAGGATTCTG-3’. The construct was cloned into pCRII-Blunt-TOPO (Invitrogen) and further subcloned into phCMV1 (Genlantis, Inc.) as XhoI-BamHI fragments. Transfection of HEK 293 cells with the phCMV1 sFA2 vector resulted in high expression levels and secretion of sFA2 in the culture supernatant. Stable HEK 293 cell line secreting sFA2 were generated using geneticin selection and cloned by limited dilution.
293 cells expressing S-peptide tagged sFA2 were grown to confluency in 1700 cm2 tissue culture roller bottles in Dulbecco’s Modified Eagle’s Medium (Quality Biologicals) supplemented with 10% cosmic calf serum (Hyclone), 2 mM L-glutamine, 100 units/ml penicillin and streptomycin and 10 μg/ml gentamicin (Quality Biologicals), then with reduced serum Opti-MEM (Invitrogen) for 72 h at 37°C with CO2. The supernatant was clarified by centrifugation, supplemented with 0.1 M L-arginine (Sigma-Aldrich) and 0.1% Triton X-100, and filtered through a low-protein binding membrane before being passed over an XK26 column (Amersham Pharmacia Biotech) with 20 ml of S-protein agarose (Novagen, Inc.) at a flow rate of approximately 5 ml/min. The column was washed with phosphate buffered saline (PBS) with 0.1% Triton X-100 and 0.1 M L-arginine and then with PBS with 0.5 M NaCl, 0.5% Triton X-100, 0.1 M L-Arginine and 0.02 M Tris. The sF protein was eluted with 0.2 M L-Arginine, 0.2 M citric acid, pH 2 and neutralized with HEPES, pH 8. The eluate was then concentrated using 30 kDa Centricon centrifugal filter units (Millipore) and buffer exchanged into PBS with 0.2 M L-arginine and 0.01% Triton X-100. Protein concentrations were determined by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and Coomassie staining with SimplyBlue SafeStain (Invitrogen) and densitometry analysis with Scion Image software by comparison to known protein standards. The concentrated sFA2 glycoprotein had an approximate molecular size of 58 kDa.
Due to the large numbers of serum samples to be tested, and the need for additional protein antigen, we also obtained sF made from strains NL\1\00, an A1 genotype, and NL\1\99, a B1 genotype, from MedImmune, Inc. as described by Ulbrandt et al. [24]. All three versions of soluble F (sFA1, sFA2, sFB1) and RSV sF (also provided by MedImmune, Inc.) bound to positive human control sera in Western blot.
Western Blot
Purified proteins were separated by SDS-PAGE on a NuPAGE Novex 4-12% Bis-Tris Gel (Invitrogen), transferred to nitrocellulose membranes and then blocked with 5% milk in PBS with 0.5% Tween 20 (PBS-T) (Sigma-Aldrich) overnight at 4°C or for 1-2 h at room temperature. The membranes were incubated for 1 h at room temperature with antiserum at 1:25,000 dilution. Membranes were washed four times with PBS-T and incubated with horseradish peroxidase-conjugated ImmunoPure goat anti-human IgG (Pierce Biotechnology) at 1:25,000 dilution for 1 h at room temperature. The membranes were washed again four times with PBS-T and visualized by chemiluminescence (Super-Signal West Pico chemiluminescent substrate, Pierce Biotechnology).
Enzyme-linked Immunosorbent Assay (ELISA)
Ninety-six well Immulon II microtiter plates (Dynex Technologies) were coated with purified sF protein from hMPV or RSV in 50 mM sodium bicarbonate and 20 mM sodium carbonate buffer (pH 9.6) at a concentration of 50 ng of protein/well and incubated overnight at 4°C. The plates were washed with PBS-T and blocked overnight at 4°C or for 1 h at 37°C with PBS-T with 5% bovine serum albumin (BSA) (Sigma-Aldrich) added. The plates were washed with PBS-T and dilutions of serum in PBS-T containing 1% BSA were added and incubated for 1 h at 37°C. The plates were washed and incubated for 1 h at 37°C with alkaline phosphatase-conjugated goat anti-human IgG (Jackson ImmunoResearch) diluted 1:5000. The plates were washed again with the same solution and antibodies were detected by adding 100 μl of PNPP substrate (Pierce Biotechnology) to each well and incubating for 30 min at room temperature and then adding 50 μl of 2 N NaOH to each well to stop the reaction. The absorption at 405 nm was measured on a VersaMax microtiter plate reader (Molecular Devices).
Paired acute- and convalescent-phase serum samples were assayed in duplicate on the same 96-well microtiter plate at 3 screening dilutions using sFA1 genotype as the antigen (1:100, 1:500 and 1:5000) against a standard dilution of hMPV antisera. Serial dilutions of known positive hMPV sera (reference sera kindly provided by Dr. Jeffrey Kahn, Yale University, New Haven, CT) were run on every ELISA plate and a standard curve generated. The positive cutoff was considered the first optical density (OD) value on the standard curve that was 1.5 times the previous dilution. All OD values used for screening and definitive testing were on the linear portion of the standard curve.
Samples were considered negative if the acute and convalescent sera tested negative at the 1:100 dilution and confirmed with repeat serial 2-fold diutions starting at 1:64. Samples were considered as evidence of a previous infection but no recent reinfection if both the acute and convalescent samples contained equivalent ELISA titers at all positive dilutions. If they were both positive at 1:5000, they were retested in serial 2-fold dilutions out to 1:262,144 to confirm. Samples were considered as possible reinfections if the convalescent titer was positive at 1:5000 and there was at least a 2-fold rise in the OD value between the acute and convalescent titer at 1:5000. Samples were considered as possible new infections when the acute samples showed no evidence of antibody and the convalescent titer was positive at 1:5000. All possible reinfections and new infections were retested in serial 2-fold dilutions. In the retest, a positive antibody response to hMPV was defined as a paired serum sample in which there was a ≥ 4-fold increase in the level of IgG antibodies between the acute-phase and the convalescent-phase serum sample.
All confirmed positive samples were also tested for evidence of increasing titers to hMPV sFA2 and sFB1 and RSV sF in serial 2-fold dilutions, with a ≥ 4-fold increase in titer considered positive. For the RSV control, we used serial dilutions of concentrated human RSV antibody (kindly provided by Dr. Jorge Blanco, Virion Systems, Inc., Rockville, MD) on each plate to generate a standard curve.
Statistical Analysis
Means were compared with Student’s t-test and proportions were compared using the χ2 test.
RESULTS
Study population demographics
In the initial dengue study, acute and convalescent serum samples were tested for evidence of recent infection with dengue and Japanese encephalitis viruses and 2,557 paired sera were negative. For these samples, the ages ranged from 7 to 16 years with an average age of 9 years, 11 months, 50.6% were male, 73.9% reported cough, rhinorrhea or both, and the average temperature at the acute visit was 38.1°C. We tested 1,380 (54%) of these samples for evidence of hMPV past and present exposure based on sample and reagent availability. These samples were from 1,049 children as 254 children had 2 to 6 febrile illnesses during the study period. All of the samples taken in 1999 and 2000 were tested, and 1/3 from 2001 and 1/2 from 2002 were randomly selected and tested. We tested the samples from 1998 that were available; from 1 June-29 July, 18-22 September and 21-29 October 1998. There were no statistically significant differences in gender or age of those tested and untested (table 1).
Table 1.
Characteristics of children with samples in the serum bank, those tested and those negative and positive for hMPV.
| Tested Samples | |||||
|---|---|---|---|---|---|
| Enrolled children |
Untested n=1177 |
Tested n=1380 |
Negative n=1312 |
Positive n=68 |
p-Value* |
| Age range (years) | 6-15 | 7-16 | 7-16 | 7-14 | |
| Mean age (years) (1 SD) | 10.1 (1.5) | 9.9 (1.6) | 9.9 (1.6) | 10 (1.6) | p > .05 |
| Male/female sex (ratio) |
594/583 (1.02) |
700/680 (1.03) |
664/648 (1.03) |
36/32 (1.13) |
p > .05 |
| Clinical symptoms+ | |||||
| Cough | 840/1162 (72.3%) |
860/1375 (62.5%) |
802/1307 (61.2%) |
58/68 (85.3%) |
p < .001 |
| Rhinorrhea | 661/1160 (57.0%) |
683/1371 (49.8%) |
638/1303 (49.0%) |
45/68 (66.2%) |
p < .01 |
| Mean temperature °C§ | 38.2 | 38.1 | 38.1 | 37.8 | p <.001 |
P-value is the same for comparison between untested and tested, and positive and negative for age and sex. P-value only for comparison between all tested and positive samples for clinical symptoms. χ2 test for categorical data and t-test for continuous variables.
Some variables were missing as indicated in the totals for cough and rhinorrhea.
Temperature at time of health care worker visit.
SD = Standard deviation
Serosurvey
Out of the 1,049 patients tested, all but 3 (99.7%) had evidence of prior exposure to hMPV. One child had a fever twice in 1998 and tested negative for prior exposure for hMPV both times. Of the 1,380 acute and convalescent serum samples tested, we found 74 that met our screening criteria for a potential new or reinfection and an additional 21 that had both acute and convalescent sera positive at the 1:5000 dilution. Of the 74 potential infections, 64 were positive for a reinfection and 2 for a new infection. Of the 21 indeterminate screening tests, 2 were positive for reinfection, for a total of 66 presumptive reinfections (4.8%) and 2 new infections (0.1%). A total of 66 children made up the 68 positive cases, with two children demonstrating reinfections in 1998 and again in 2000. Table 2 lists the results by year. As the samples were screened and retested using protein provided by MedImmune, Inc. (sFA1), all of the non-exposed, new infections and reinfections were then retested using sFA2 in the same assay conditions with the same results (data not shown).
Table 2.
Results of human serosurvey
| Year | Number paired sera tested of total collected | ≥ 4-fold rise in titer | |||
|---|---|---|---|---|---|
| 1998 | 390/1,083 (36.0%) | 14 (3.6%)* | |||
| 1999 | 326/341 (95.6%) | 1 (0.3%) | |||
| 2000 | 364/372 (97.8%) | 48 (13.2%)* | |||
| 2001 | 157/461 (34.1%) | 2 (1.3%) | |||
| 2002 | 143/300 (47.7%) | 3 (2.1%) | |||
| Total | 1,380/2,557 (54.0%) | 68 (4.9%) | |||
One case a presumptive new infection with negative acute sera.
The acute specimens that were negative for hMPV sF ELISA and the paired specimens that showed a potential new infection were further tested by Western blotting. A representative example of each one is shown in figure 1. In each case acute sera from these samples were negative for the three different subtypes of hMPV F and positive for antibody to RSV F. The two new infections (figure 1A) demonstrated antibody to all three F subtypes in the convalescent sera. Although we cannot exclude the possibility of the presence of antibody below the limits of detection in the acute sera, it appears that 4 samples were negative for exposure in the acute sample, and two of these demonstrated a new infection with hMPV in the convalescent sample, confirming the ELISA results.
Figure 1.
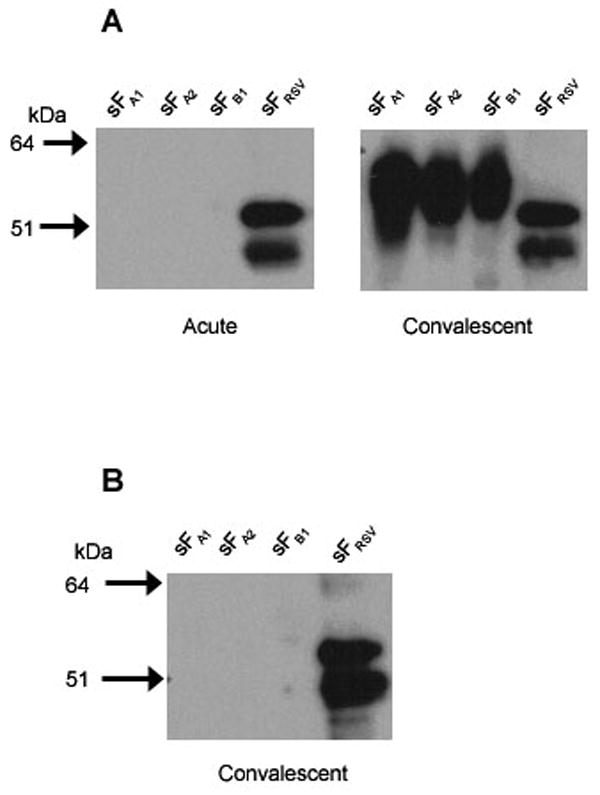
Western blot of presumptive hMPV negative and new infection sera. Purified hMPV sFA1, sFA2, and sFB1 and RSV sF were resolved on 4-12% Nu-PAGE gels and transferred to nitrocellulose membranes. The membranes were probed with human sera as follows: (A) acute and convalescent sera from a patient after the acute sample tested negative but the convalescent sample tested positive for hMPV sF antibody in ELISA, or (B) RSV positive convalescent sera from a patient that was ELISA negative for hMPV sF in both acute and convalescent sera. Bound antibodies were reacted with a peroxidase conjugated goat anti-human IgG and visualized by chemiluminescence.
If A and B represent different serologic subtypes, it is possible that the ELISA could be negative if the patients were infected with lineage B and sF lineage A was used as the antigen. Leung et al. demonstrated no difference in ELISA reactivity using cell lysates infected with recombinant VSV expressing A or B lineages as antigen [6]. To verify that our assay was also equally sensitive for the two lineages, we further tested 12 paired sera with evidence of reinfection and 48 paired sera with only evidence of a prior exposure using the sFB1 protein as antigen and obtained identical results to the sFA1 protein (data not shown).
Nevertheless, antibodies can cross-react to shared epitopes and we considered the possibility of potential cross-reactivity to RSV as the RSV F glycoprotein possesses 30-43% homology to hMPV F [28]. Therefore, we assayed the 68 hMPV reinfection positive paired sera against RSV F. Among these, only 18 (26%) had a four-fold or greater rise in antibody titer to the RSV antigen, and only 3 of the 18 had a greater titer increase to RSV than hMPV (data not shown). While we cannot rule out cross-reactivity, it is also possible that at least some of those positive for both hMPV and RSV reinfection represent coinfections with more than one virus.
Epidemiology of hMPV in the study population
We found the majority of reinfections in 1998 and 2000. We were not able to test a representative sample of the study period in 1998, but 48 of the 68 new or re-infections occurred in 2000, accounting for 13.2% of all non-flaviviral febrile illnesses in the study population in that year. Thirty-one of these cases occurred in a discrete 2 week time period (figure 2), suggesting a localized outbreak. Twenty of these cases were from 3 of 12 participating schools, with 3 other schools accounting for the remaining 11 cases. Although clinical information on the patients used in the present study was limited, and a majority of all study children had respiratory symptoms, a statistically significant higher number of hMPV-positive patients complained of rhinorrhea or cough when compared to all of the patients tested (table 1), consistent with a respiratory disease causing their illness.
Figure 2.
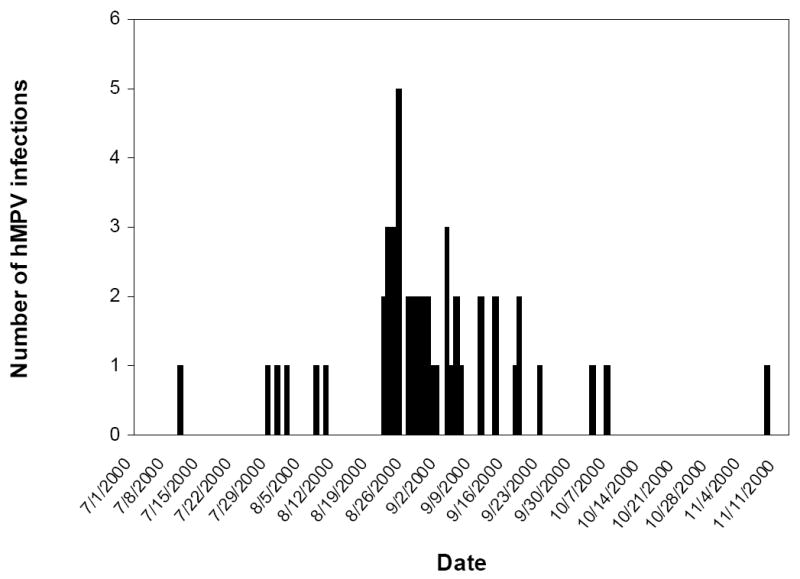
Epidemiologic curve of cases with a 4-fold or greater rise in IgG titer to hMPV sF between acute and convalescent serum samples representing reinfection with hMPV in the year 2000 in the study population.
DISCUSSION
This is the first report of the seroepidemiology of hMPV in Thailand, and the first study to systematically examine reinfection rates among children. Using our ELISA with recombinant F glycoprotein as antigen to detect hMPV antibodies, we found the seroprevalence in this population nearly 100%. The reinfection rate (average 4.9%/year) was similar to that seen in adults [18]. We believe that this number is probably an underestimate for a number of technical reasons. Firstly, we used very conservative criteria to determine cut-offs for potential reinfections in the initial screen. Many paired sera had higher OD values for convalescent sera than acute sera, but did not meet the criteria of having the convalescent sera positive at 1:5000 or did not have at least 2 times the OD reading at this dilution (e.g., 257 convalescent sera had an OD reading between 1.5 and 2 times the acute sera). Secondly, for a blood sample to be taken, a child had to have a history of fever. Several published reports quote rates of fever in hMPV infections near 50% or lower [13, 14, 17, 18, 29, 30]. Especially given that these illnesses are almost entirely reinfections, it is possible that there were other cases of hMPV that were symptomatic, but did not meet the fever criteria. Thirdly, it is possible that the peak season for hMPV is in the winter months as discussed below, and we did not have samples from this time of year. Finally, it is possible that patients may mount such a rapid IgG response to a reinfection that the ability to detect a change between acute and convalescent titers could be diminished. The average length of time between onset of illness and drawing the acute sample was 1.2 days in all of the tested patients. Since IgG levels typically increase within 2 days following re-exposure [31], this could occur before symptoms are even apparent, and may result in increased titers in the acute specimen. Two of 7 patients with known hMPV reinfections by viral isolation in another study did not develop a four-fold rise in titer between acute and convalescent samples [10].
We could not confirm that elevations of antibody reactive to hMPV F glycoprotein correlated with actual reinfections with the virus, since we did not have respiratory samples. Indeed, we did see that 27% of all hMPV reinfection positive samples reacted to RSV F. Although all existing human data demonstrate a lack of cross-reactivity between conserved hMPV proteins and RSV antibodies [6, 20, 25], there is the potential that cross-reactivity to RSV could be causing some of the hMPV positive results. It may be possible to use sera from known hMPV and RSV cases and compare their ability to neutralize recombinant viruses expressing the hMPV fusion protein to determine the extent of their ability to bind to F. We also did not conduct parallel testing using hMPV infected cell lysates as the antigen in ELISA, and this could assist in confirming our results.
There are many reports of coinfections of various respiratory viruses, including hMPV. Although most have reported low rates of coinfection, some studies have reported 16-30% of hMPV-positive samples co-infected with other viral or bacterial pathogens [32-34]. Therefore, while unlikely that all specimens positive for both RSV and hMPV represent coinfections, it is possible that some of them are true coinfections. It is also important to document the potential for cross-reactivity or non-specific antibody increases as serological tests are developed for pneumoviruses.
Two different genotypes of hMPV have been documented, and some studies suggest they may represent two different serotypes based on neutralization capability [35]. However, we have demonstrated a 100% concordance using both A and B lineages of hMPV F by ELISA, demonstrating that significant cross-reactivity occurs between the two F glycoproteins. If they are separate serotypes, they cannot be distinguished by reactivity to the F glycoprotein, and since F is the major antigenic determinant of virus-neutralizing antibodies, our results do not support a serotype distinction of the hMPV genotypes.
The seasonality of hMPV epidemics appears to peak in the winter to spring in temperate regions [2, 36], although there are reports of summer outbreaks [37]. There are few published studies regarding transmission of hMPV in tropical areas, but one study in Hong Kong demonstrated hMPV activity only from January to July [38]. Another study in New Delhi detected the majority of hMPV infections from December through February [39], and a report of samples collected during July and August in central India showed 5 of 26 positive [40]. A retrospective serosurvey in Bangladesh demonstrated all cases of hMPV occurring from January to July with a peak in April [17]. Interestingly, in one study in Thailand, all 6 hMPV cases detected during a 2 year period occurred between August and November [5]. Here we also demonstrate active infection throughout the summer and fall, with an outbreak occurring in August. However, given the usually reported pattern, if samples had been taken in the winter through spring months, it is possible that infection or reinfection with hMPV may have accounted for an even greater percentage of illnesses in this population.
Taken together, our findings demonstrate that recombinant hMPV F is an antigenic glycoprotein and useful for the detection of antibodies to hMPV. This is the first study to use purified, recombinant F by ELISA for screening and determination of infection rates amongst a human cohort. We have obtained evidence that reinfection of hMPV is a significant cause of illness in older pediatric populations. Further studies are required to clearly define the role hMPV plays in this population.
Acknowledgments
The authors would like to thank Dr. James Young of MedImmune, Inc., for helpful discussions, Dr. Ed Mitre for assistance with standard curve generation and analysis and Dr. Cara Olsen for statistical assistance.
This project and publication were supported by NIH Grant AI34533 and the United States Army Medical Research and Materiel Command, Ft Detrick MD.
Footnotes
The authors declare that they have no competing or commercial interests.
The views expressed in the manuscript are solely those of the authors, and do not represent official views or opinions of the Uniformed Services University, the Department of Defense, or the National Institutes of Health.
Portions of this work have been presented at the American Society of Tropical Medicine and Hygiene 55th Annual Meeting, November 2006, Atlanta, GA and the Infectious Disease Society of America 2007 Annual Meeting, October 2007, San Diego, CA.
References
- 1.van den Hoogen BG, de Jong JC, Groen J, et al. A newly discovered human pneumovirus isolated from young children with respiratory tract disease. Nat Med. 2001;7:719–24. doi: 10.1038/89098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Kahn JS. Epidemiology of human metapneumovirus. Clin Microbiol Rev. 2006;19:546–57. doi: 10.1128/CMR.00014-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Samransamruajkit R, Thanasugarn W, Prapphal N, Theamboonlers A, Poovorawan Y. Human metapneumovirus in infants and young children in Thailand with lower respiratory tract infections; molecular characteristics and clinical presentations. J Infect. 2006;52:254–63. doi: 10.1016/j.jinf.2005.07.001. [DOI] [PubMed] [Google Scholar]
- 4.Thanasugarn W, Samransamruajkit R, Vanapongtipagorn P, et al. Human metapneumovirus infection in Thai children. Scand J Infect Dis. 2003;35:754–6. doi: 10.1080/00365540310000094. [DOI] [PubMed] [Google Scholar]
- 5.Teeratakulpisarn J, Ekalaksananan T, Pientong C, Limwattananon C. Human metapneumovirus and respiratory syncytial virus detection in young children with acute bronchiolitis. Asian Pac J Allergy Immunol. 2007;25:139–45. [PubMed] [Google Scholar]
- 6.Leung J, Esper F, Weibel C, Kahn JS. Seroepidemiology of human metapneumovirus (hMPV) on the basis of a novel enzyme-linked immunosorbent assay utilizing hMPV fusion protein expressed in recombinant vesicular stomatitis virus. J Clin Microbiol. 2005;43:1213–9. doi: 10.1128/JCM.43.3.1213-1219.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Henderson FW, Collier AM, Clyde WA, Jr, Denny FW. Respiratory-syncytial-virus infections, reinfections and immunity. A prospective, longitudinal study in young children. N Engl J Med. 1979;300:530–4. doi: 10.1056/NEJM197903083001004. [DOI] [PubMed] [Google Scholar]
- 8.Falsey AR, Walsh EE. Viral pneumonia in older adults. Clin Infect Dis. 2006;42:518–24. doi: 10.1086/499955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Ebihara T, Endo R, Ishiguro N, Nakayama T, Sawada H, Kikuta H. Early reinfection with human metapneumovirus in an infant. J Clin Microbiol. 2004;42:5944–6. doi: 10.1128/JCM.42.12.5944-5946.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ebihara T, Endo R, Kikuta H, et al. Human metapneumovirus infection in Japanese children. J Clin Microbiol. 2004;42:126–32. doi: 10.1128/JCM.42.1.126-132.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ebihara T, Endo R, Kikuta H, et al. Seroprevalence of human metapneumovirus in Japan. J Med Virol. 2003;70:281–3. doi: 10.1002/jmv.10391. [DOI] [PubMed] [Google Scholar]
- 12.Pelletier G, Dery P, Abed Y, Boivin G. Respiratory tract reinfections by the new human Metapneumovirus in an immunocompromised child. Emerg Infect Dis. 2002;8:976–8. doi: 10.3201/eid0809.020238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Williams JV, Harris PA, Tollefson SJ, et al. Human metapneumovirus and lower respiratory tract disease in otherwise healthy infants and children. N Engl J Med. 2004;350:443–50. doi: 10.1056/NEJMoa025472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Williams JV, Wang CK, Yang CF, et al. The role of human metapneumovirus in upper respiratory tract infections in children: a 20-year experience. J Infect Dis. 2006;193:387–95. doi: 10.1086/499274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Chan PK, Tam JS, Lam CW, et al. Human metapneumovirus detection in patients with severe acute respiratory syndrome. Emerg Infect Dis. 2003;9:1058–63. doi: 10.3201/eid0909.030304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ishiguro N, Ebihara T, Endo R, et al. Immunofluorescence assay for detection of human metapneumovirus-specific antibodies by use of baculovirus-expressed fusion protein. Clin Diagn Lab Immunol. 2005;12:202–5. doi: 10.1128/CDLI.12.1.202-205.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Brooks WA, Erdman D, Terebuh P, et al. Human metapneumovirus infection among children, Bangladesh. Emerg Infect Dis. 2007;13:1611–1613. doi: 10.3201/eid1310.070337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Falsey AR, Erdman D, Anderson LJ, Walsh EE. Human metapneumovirus infections in young and elderly adults. J Infect Dis. 2003;187:785–90. doi: 10.1086/367901. [DOI] [PubMed] [Google Scholar]
- 19.Liu L, Bastien N, Sidaway F, Chan E, Li Y. Seroprevalence of human metapneumovirus (hMPV) in the Canadian province of Saskatchewan analyzed by a recombinant nucleocapsid protein-based enzyme-linked immunosorbent assay. J Med Virol. 2007;79:308–13. doi: 10.1002/jmv.20799. [DOI] [PubMed] [Google Scholar]
- 20.Hamelin ME, Boivin G. Development and validation of an enzyme-linked immunosorbent assay for human metapneumovirus serology based on a recombinant viral protein. Clin Diagn Lab Immunol. 2005;12:249–53. doi: 10.1128/CDLI.12.2.249-253.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Skiadopoulos MH, Biacchesi S, Buchholz UJ, et al. Individual contributions of the human metapneumovirus F, G, and SH surface glycoproteins to the induction of neutralizing antibodies and protective immunity. Virology. 2006;345:492–501. doi: 10.1016/j.virol.2005.10.016. [DOI] [PubMed] [Google Scholar]
- 22.Skiadopoulos MH, Biacchesi S, Buchholz UJ, et al. The two major human metapneumovirus genetic lineages are highly related antigenically, and the fusion (F) protein is a major contributor to this antigenic relatedness. J Virol. 2004;78:6927–37. doi: 10.1128/JVI.78.13.6927-6937.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Cseke G, Wright DW, Tollefson SJ, Johnson JE, Crowe JE, Jr, Williams JV. Human metapneumovirus fusion protein vaccines that are immunogenic and protective in cotton rats. J Virol. 2007;81:698–707. doi: 10.1128/JVI.00844-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ulbrandt ND, Ji H, Patel NK, et al. Isolation and characterization of monoclonal antibodies which neutralize human metapneumovirus in vitro and in vivo. J Virol. 2006;80:7799–806. doi: 10.1128/JVI.00318-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Alvarez R, Jones LP, Seal BS, Kapczynski DR, Tripp RA. Serological cross-reactivity of members of the Metapneumovirus genus. Virus Res. 2004;105:67–73. doi: 10.1016/j.virusres.2004.04.019. [DOI] [PubMed] [Google Scholar]
- 26.Endy TP, Chunsuttiwat S, Nisalak A, et al. Epidemiology of inapparent and symptomatic acute dengue virus infection: a prospective study of primary school children in Kamphaeng Phet, Thailand. Am J Epidemiol. 2002;156:40–51. doi: 10.1093/aje/kwf005. [DOI] [PubMed] [Google Scholar]
- 27.Endy TP, Nisalak A, Chunsuttiwat S, et al. Spatial and temporal circulation of dengue virus serotypes: a prospective study of primary school children in Kamphaeng Phet, Thailand. Am J Epidemiol. 2002;156:52–9. doi: 10.1093/aje/kwf006. [DOI] [PubMed] [Google Scholar]
- 28.Bastien N, Normand S, Taylor T, et al. Sequence analysis of the N, P, M and F genes of Canadian human metapneumovirus strains. Virus Res. 2003;93:51–62. doi: 10.1016/S0168-1702(03)00065-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Baer G, Schaad UB, Heininger U. Clinical findings and unusual epidemiologic characteristics of human metapneumovirus infections in children in the region of Basel, Switzerland. Eur J Pediatr. 2007 doi: 10.1007/s00431-007-0427-x. [DOI] [PubMed] [Google Scholar]
- 30.Manoha C, Espinosa S, Aho SL, Huet F, Pothier P. Epidemiological and clinical features of hMPV, RSV and RVs infections in young children. J Clin Virol. 2007;38:221–6. doi: 10.1016/j.jcv.2006.12.005. [DOI] [PubMed] [Google Scholar]
- 31.Birdsall H. Antibodies. In: Mandell GL, Bennett JE, Dolin R, editors. Mandell, Douglas and Bennett’s Principles and Practice of Infectious Disease. 6. Vol. 1. Philadelphia: Elsevier; 2005. pp. 52–68. [Google Scholar]
- 32.Boivin G, Abed Y, Pelletier G, et al. Virological features and clinical manifestations associated with human metapneumovirus: a new paramyxovirus responsible for acute respiratory-tract infections in all age groups. J Infect Dis. 2002;186:1330–4. doi: 10.1086/344319. [DOI] [PubMed] [Google Scholar]
- 33.Garcia-Garcia ML, Calvo C, Casas I, et al. Human metapneumovirus bronchiolitis in infancy is an important risk factor for asthma at age 5. Pediatr Pulmonol. 2007;42:458–64. doi: 10.1002/ppul.20597. [DOI] [PubMed] [Google Scholar]
- 34.Bosis S, Esposito S, Niesters HG, Crovari P, Osterhaus AD, Principi N. Impact of human metapneumovirus in childhood: comparison with respiratory syncytial virus and influenza viruses. J Med Virol. 2005;75:101–4. doi: 10.1002/jmv.20243. [DOI] [PubMed] [Google Scholar]
- 35.van den Hoogen BG, Herfst S, Sprong L, et al. Antigenic and genetic variability of human metapneumoviruses. Emerg Infect Dis. 2004;10:658–66. doi: 10.3201/eid1004.030393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Peret TC, Hall CB, Hammond GW, et al. Circulation patterns of group A and B human respiratory syncytial virus genotypes in 5 communities in North America. J Infect Dis. 2000;181:1891–6. doi: 10.1086/315508. [DOI] [PubMed] [Google Scholar]
- 37.Louie JK, Schnurr DP, Pan CY, et al. A summer outbreak of human metapneumovirus infection in a long-term-care facility. J Infect Dis. 2007;196:705–8. doi: 10.1086/519846. [DOI] [PubMed] [Google Scholar]
- 38.Peiris JS, Tang WH, Chan KH, et al. Children with respiratory disease associated with metapneumovirus in Hong Kong. Emerg Infect Dis. 2003;9:628–33. doi: 10.3201/eid0906.030009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Banerjee S, Bharaj P, Sullender W, Kabra SK, Broor S. Human metapneumovirus infections among children with acute respiratory infections seen in a large referral hospital in India. J Clin Virol. 2007;38:70–2. doi: 10.1016/j.jcv.2006.07.003. [DOI] [PubMed] [Google Scholar]
- 40.Rao BL, Gandhe SS, Pawar SD, Arankalle VA, Shah SC, Kinikar AA. First detection of human metapneumovirus in children with acute respiratory infection in India: a preliminary report. J Clin Microbiol. 2004;42:5961–2. doi: 10.1128/JCM.42.12.5961-5962.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]