

Abstract
We tested four chimpanzees in a self-control task in which food rewards accumulated as long as they were not eaten. In one condition, the chimpanzees had to perform a computer task that directly led to the delivery of the food rewards. In another condition, working on the computerized task was not required and any such work was not linked to the delivery of rewards. The third condition offered no computerized task (chimpanzees simply waited for food rewards to be delivered). Three of four chimpanzees showed no effect of the work scenario on delay of gratification. The one chimpanzee that showed an influence of work scenario on self-control was the overall poorest performing animal. This animal delayed gratification the longest, however, when work was required and reward delivery was directly linked to that work. Therefore, although there is little evidence linking delay of gratification to work requirements in chimpanzees, chimpanzees with lower overall self-control might benefit from having some work available if reward accumulation is contingent on performing that work.
Keywords: Chimpanzees, Delay of Gratification, Inhibition, Pan Troglodytes, Self-Control, Effort
Introduction
Comparative research suggests that there is continuity across species with regard to both self-control and impulsivity (e.g., Beran, Savage-Rumbaugh, Pate, & Rumbaugh, 1999; Evans & Westergaard, 2006; Logue, 1988; Logue, Forzano, & Ackerman, 1996; Rachlin, 2000; Stevens et al., 2005; Tobin et al., 1996; van Haaren, van Hest, & van De Poll, 1988). Delaying gratification (or postponing a response to a present reward for the sake of a future bigger or better reward) is one of the hallmark aspects of self-control (Mischel, Shoda, & Rodriguez, 1989). It sometimes requires an organism to maintain a course of action (or inaction) in the face of continual competition from the available, impulsive response so that a future, preferred outcome can be obtained (e.g., not spending all of one's money to increase the balance of a bank account or avoiding unhealthy foods when on a diet so as to maximize weight loss). In some cases, the delayed reward may be accumulating across time. This creates both an increase in the impulsive pull of that reward (as it grows, it becomes more tempting to remove the money and spend it) and an increase in the value of the reward (the longer it is left alone, the more there will finally be). This article is concerned with this form of delaying gratification in the face of increasingly more tempting rewards.
In the present study, we used a delay maintenance paradigm in which the amount of reward increased as long as a participant inhibited taking that reward. This paradigm has been useful for investigating the role that various environmental aspects and behavioral tendencies of the participant have on self-control, and it has been used with human children (Toner & Smith, 1977; Toner, Lewis, & Gribble, 1979) and nonhuman primates (e.g., Beran, 2002; Beran & Evans, 2006; Evans & Beran, 2007a, b). For example, through the presentation of this paradigm to human children, it has become well-known that reward-directed attention is detrimental to delay in situations where children simply wait during a delay interval (e.g., Mischel, 1974, 1981; Mischel & Baker, 1975; Mischel & Ebbeson, 1970).
In nearly all previous studies of delay maintenance, both with humans and nonhuman animals, the reward accumulation sequence has been out of the control of the participant. Delayed rewards accumulated no matter what participants did during that time, and so there was no way for participants to control the rate of accumulation or to feel any control over the delivery of delayed rewards. There have been some cases in which children engaged in distracting play or thoughts when faced with delay of gratification tasks (Cournoyer & Trudel, 1991; Miller & Karniol, 1976; Michel, Ebbesen, & Zeiss, 1972; Toner & Smith, 1977), but the children could only use these tactics to direct their attention and thoughts away from the tempting delayed rewards. Additionally, there is evidence that chimpanzees will distract themselves with task-irrelevant objects such as toys during a delay period, and such self-distraction prolongs their delay maintenance performance (Evans & Beran, 2007a). But, again, the chimpanzees could only extend control over their attention to the reward items, but had no way to control any aspect of the accumulation of rewards.
The few studies that have given participants some way to influence the parameters of the delay task have involved human children and have focused on the relation between attention to delayed rewards and delay of gratification. In one study, Patterson and Carter (1979) tested children in one situation in which they had to work (i.e. fulfill a response requirement) to obtain a delayed reward. They also tested children in the typical self-control situation in which they simply waited for end of the delay period to obtain a reward. Patterson and Carter (1979) found that, contrary to the classic finding, attention to delayed rewards actually facilitated delay maintenance in the working situation. Peake, Hebl, and Mischel (2002) conducted a similar study, except that they tested children in an additional situation in which work was available, but was not required, in order to obtain the delayed reward. They found that, in the presence of rewards, children waited longer when work was available than when it was not, but there was no difference in delay times when obtaining the better reward was contingent on doing the work and when it was not. This suggested that the effects of working to obtain delayed rewards were the result of the work providing a distraction to the children.
Despite these findings in human children, it is unknown whether working to control the delivery of delayed rewards might increase or decrease delay of gratification in chimpanzees. For both species, self-distraction seems to facilitate increased delay of gratification (chimpanzees: Evans & Beran, 2007a; children: Miller & Karniol, 1976; Michel, Ebbeson & Zeiss, 1972; Toner & Smith, 1977). However, chimpanzees seem much less negatively affected by attention to delayed rewards in comparison to children (Beran, 2002; Beran & Evans, 2006). Thus, it is uncertain what effect a concurrent task that may increase or decrease chimpanzees' attention to delayed reward might have on chimpanzee delay maintenance. For the same reason, it is uncertain whether making that concurrent task a requirement or simply an option would affect delay maintenance differently in chimpanzees.
Having something to do during delay intervals can be beneficial to self-control of other non-primate animals. Grosch and Neuringer (1981) found that pigeons more often chose a more preferred, delayed reinforcer over a less preferred, immediate reinforcer when they could perform some unrelated activity during the delay interval compared to when they could not perform that activity. Presumably, this was because the activity offered a distraction that benefited self-control behavior. Chelonis, Logue, Sheehy, and Mao (1998) showed that, when they steadily increased the amount of force required for rats to select both an immediate small reward and a delayed large reward, the rats increasingly chose the delayed reward. Thus, actually requiring additional work to obtain any reward can facilitate self-control choice.
However, there is also evidence that some animals show decreases in self-control when increased effort is required. In one example, pigeons performed a task in which they chose between pecking a key a small number of times to receive a small reward versus pecking another key a larger number of times to receive a large reward (Grossbard & Mazur, 1986). As the required number of presses increased, the likelihood of choosing the smaller reward increased. Pigeons also seemingly revalued food rewards as a function of the amount of effort required to obtain those rewards (e.g., Clement, Feltus, Kaiser, & Zentall, 2000). Evans (2007) presented rhesus monkeys with a task in which delay interval and work effort were manipulated within a self-control task. The monkeys showed some evidence that increased effort led to decreased self-control, but the result was complicated by an interaction with overall delay length. This body of evidence leaves open the possibility that a work requirement or option associated with the accumulation of a delayed reward could have either a positive or negative effect overall on delay maintenance in chimpanzees.
To examine this question, we tested four chimpanzees in three different conditions in which candies were delivered into a container that was accessible to the chimpanzee throughout the session, and the session continued as long as a chimpanzee did not eat any of the candies. Two of the conditions included a concurrent computerized psychomotor task in addition to a delay maintenance requirement. This task was a highly familiar computer program in which the chimpanzees had to move the cursor into contact with a number of moving circles on the screen. In one of the two conditions involving the concurrent task, reward accumulation was contingent upon the work completed during the task. In the other it was not. The third condition did not include the concurrent task. We compared the number of reward items obtained by the chimpanzees and the duration of delay in these three conditions to examine what effect concurrent work and the requirement to perform that work had on delay maintenance.
We expected to see one of three effects of the working situation on chimpanzees' delay maintenance. One possibility was that the concurrent computer task would provide a distraction to chimpanzees, and they would use it to divert their attention away from the visible delayed rewards. If this were the case, the chimpanzees would show the shortest delay times in the no work condition, and equally high delay times in the two conditions involving the concurrent task. Also, in this case, we would expect to find a positive relationship between the percentage of delay time the chimpanzees spent working on the noncontingent task and the amount of rewards obtained in that condition. A second possibility was that the observable contingency between concurrent task activity and reward delivery would provide the chimpanzees with perceived control over the delayed reward accumulation. Nonhuman primates have exhibited enhanced performance in other test paradigms in response to perceived control over task parameters such as task order (Beran et al., 2007; Washburn, Hopkins, & Rumbaugh, 1990). Such an effect in this experiment would be evidenced by longer delay times in the contingent work condition than in either of the other two conditions, in which work was not required to obtain rewards. A third possibility was that persistent performance of the concurrent task would create an additional self-regulatory burden to delay maintenance. In this case, chimpanzees would maintain delay the longest in the no work condition and maintain delay the shortest in the contingent work condition. Also, in this case, we would expect the chimpanzees to exhibit a negative relationship between the percentage of delay time spent working on the noncontingent task and the amount of rewards obtained in that condition.
Materials and methods
Subjects
Four chimpanzees were tested: Lana (37-year-old female), Sherman (34-year-old male), Panzee (22-year-old female), and Mercury (21-year-old male). The chimpanzees were familiar with the delay of gratification paradigm used in the present study (Beran & Evans, 2006; Evans & Beran, 2007a;) and were experienced users of the computer program and apparatus implemented in two of the three conditions.
Materials
The main test apparatus consisted of a mobile cart holding a personal computer connected to a 36-item universal feeder. The dispenser was constructed mainly of translucent materials, so all potentially available food rewards (fruit flavored candies) were visible at all times. The dispenser was located approximately 1.5 m above the floor. Dispensed candies dropped into a funnel that fed through a clear plastic tube that ran down to the floor. A clear container with a closed end was attached to the tube at the floor level. This tube could be disconnected from the plastic tube with a tug or pull by the chimpanzees, and the accumulation of candies then would be accessible to the chimpanzee for consumption. The dispenser cart was used in all three experimental conditions listed below.
A computer program controlled the delivery of candies via the universal dispenser. The program produced a tone each time a candy was dispensed, and it counted the number of candies dispensed across the session. A maximum of 36 candies could be delivered on a given trial. The experimenter observed test sessions via a closed circuit monitor from outside of the test area and controlled the dispenser apparatus via a remote keyboard.
Depending on the condition, the rate and number of candies delivered was determined by the chimpanzee's performance in the concurrent task in either the present or a previous test session (see design and procedure for details). The concurrent task was presented on a 17-inch monitor, and chimpanzees made responses with their hands by manipulating a joystick attached to their home cage. This joystick controlled a cursor on the computer screen. The concurrent task (and necessary input and display equipment) was presented either on the dispenser cart (and was controlled by the same computer that controlled the dispenser) or on a separate cart with a second computer, depending on the condition (see below for details). When the second cart was used, it was presented at a distance of approximately 2 ft from the dispenser cart.
Design and Procedure
Each chimpanzee was tested individually, and each animal completed one session per day. At the beginning of each session, the experimenter stocked the dispenser with 36 candies (the maximum number that it could hold). He then placed the collection container in the chimpanzee's enclosure so that the chimpanzee could disconnect it at any time.
Each session involved one of three conditions. In the contingent work condition, the experimenter began a computer program called CHASE on the computer cart that held the dispenser (only one computer cart was present during this condition). At the same time, he presented the candy collection tube to the chimpanzee. In each trial of the CHASE task, chimpanzees had to use a joystick to move a cursor on the computer screen so as to make contact with each of seven moving green circles on the screen. The circles moved in a randomly selected initial direction, and they independently deflected and rebounded off the edge of the screen when the edge was reached. Each circle disappeared when contacted, and when all circles had been contacted the dispenser dropped one candy into the collection container. A new CHASE trial then was presented, and the task continued until the chimpanzee disconnected the collection container and ate the candies. The CHASE program recorded the total duration of each trial (from the time that the cursor appeared until the point at which the last circle was contacted) and the total amount of time that a chimpanzee manipulated the joystick within a test session.
In the noncontingent work condition, the CHASE program also was used except that the circles were purple instead of being green and the task was presented on a second cart positioned a short distance from the dispenser cart. These methodological differences were introduced to help the chimpanzees distinguish between the contingent and noncontingent task conditions that they would experience on different days (none of the chimpanzees had ever received rewards from a spatially separate computer cart in the past). Most importantly, performance on the CHASE task in this condition was not related at all to delivery of candies, and the chimpanzees did not have to engage the CHASE task at all to obtain the food rewards. Rather, the computer program running on the dispenser cart read the inter-item intervals from the most recent session in the contingent concurrent condition and dispensed a candy after each of those intervals. Thus, food delivery rate and inter-item intervals were identical between the contingent and noncontingent conditions with the only difference consisting of whether the chimpanzee controlled when the next item was dispensed by performing the CHASE task. Again, the task continued until the chimpanzee disconnected the collection container and ate the candies.
In the no work condition, all details were the same as in the contingent work condition except that there was no CHASE task available. The chimpanzees simply observed the collection container as candies were dispensed at the same rate, and with the same inter-item intervals, as in the most recent session in the contingent condition. The trial again ended when the chimpanzee disconnected the collection container and ate the candies.
Test sessions were presented to each chimpanzee in three-day blocks, in which one session of each type was conducted on a different day. The first session within a block always was the contingent work condition, and the remaining two sessions were ordered randomly. This was necessary so that the inter-item intervals could be perfectly controlled across all three conditions. Each chimpanzee completed 10 blocks for a total of 30 test days, and performance was compared within subjects across the three conditions.
Results
It is important first to note that, although the chimpanzees did not have to engage the CHASE task in the noncontingent condition, they all did so. Lana performed the CHASE task for 35% of the total trial time she completed in that condition. Panzee did do for 24% of the total time. Sherman did so for 14% of the total time, and Mercury did so for 53% of the total time. Therefore, the noncontingent condition was qualitatively different from the no work condition in that the chimpanzees were still working on the computer task for part of the time even though it was not linked to reward delivery. Also, there was no significant difference between the time each chimpanzee spent working on the noncontingent CHASE task when that condition directly followed the contingent task condition and when it directly followed the no work condition; Lana t(8) = 0.052, p = 0.96, Mercury t(8) = 0.98, p = 0.355, Panzee t(8) = 1.976, p = 0.08, Sherman t(8) = 0.671, p = 0.52 (Table 1). Therefore, it was unlikely that the chimpanzees engaged in the noncontingent task only because they confused the noncontingent condition with the contingent condition that they had experienced in the previous session.
Table 1.
Mean time-on-task as a function of experimental condition and order of presentation.
| Subject | Contingent | Noncontingent | |
|---|---|---|---|
| After Contingent | After No Work | ||
| Lana | 463.45 [161.90] | 266.28 [176.90] | 260.61 [166.32] |
| Mercury | 95.18 [33.95] | 67.58 [33.56] | 50.62 [19.16] |
| Panzee | 201.24 [89.68] | 249.96 [39.15] | 152.51 [103.09] |
| Sherman | 121.35 [101.75] | 99.08 [23.22] | 143.63 [146.68] |
Note. Values are given in seconds, and standard deviations are shown in the brackets.
Figure 1 presents the mean number of candies obtained in each of the three conditions by each chimpanzee. Only one chimpanzee, Mercury, showed a significant difference in the number of candies obtained across conditions, F(2, 18) = 17.67, p < 0.001, ηp2 = 0.66. Post hoc analyses using pair-wise t-tests indicated that Mercury obtained significantly more candies on trials in the contingent work condition compared to the two other conditions (contingent vs. no work: t(9) = 6.16, p < 0.001; contingent vs. noncontingent: t(9) = 4.21, p = 0.04), but those other two conditions did not differ from each other (t(9) = 0.81, p = 0.44). Although data from the other three chimpanzees seemed to indicate a negative influence of the contingent task condition on candies obtained, no statistically significant differences existed across conditions for these individuals; Lana F(2, 18) = 1.89, p = 0.18, ηp2 = 0.17; Sherman F(2, 18) = 1.28, p = 0.30, ηp2 = 0.12; Panzee F(2, 18) < 1.00, p = 0.76, ηp2 = 0.03. This was due to the large degree of variability in performance exhibited by these three chimpanzees within each condition.
Figure 1.
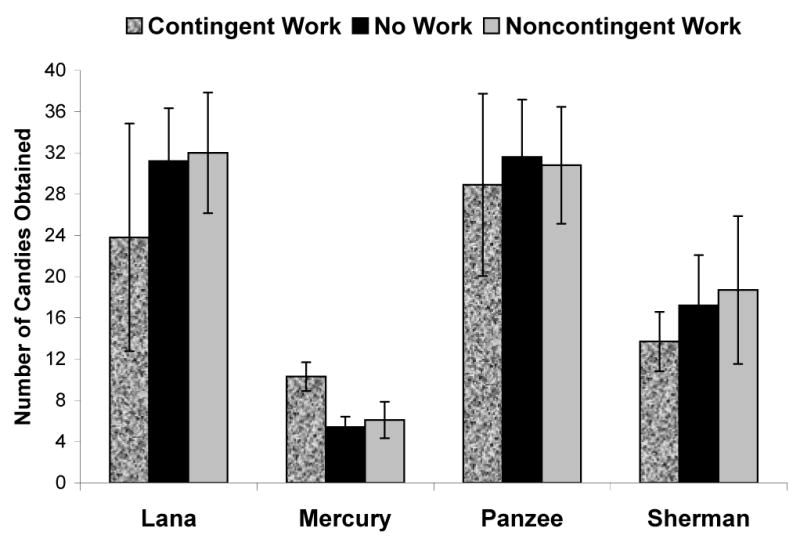
The mean number of candies obtained in each of the three conditions by each chimpanzee. Bars indicate 95% confidence intervals.
Similar analyses using duration of delay showed identical results with only Mercury producing a significant difference in the duration of delay maintenance across condition, F(2, 18) = 14.44, p < 0.001, ηp2 = 0.62; Lana F(2, 18) = 2.16, p = 0.16, ηp2 = 0.18; Sherman F(2, 18) < 1.00, p = 0.52, ηp2 = 0.07; Panzee F(2, 18) < 1.00, p = 0.67, ηp2 = 0.04. Post hoc analyses using pair-wise t-tests again indicated that Mercury maintained delay of gratification significantly longer on trials in the contingent work condition compared to the two other conditions (contingent vs. no work: t(9) = 5.34, p < 0.001; contingent vs. noncontingent: t(9) = 3.79, p = 0.004), but those other two conditions did not differ from each other (t(9) = 0.86, p = 0.412). Those data are presented in Figure 2.
Figure 2.
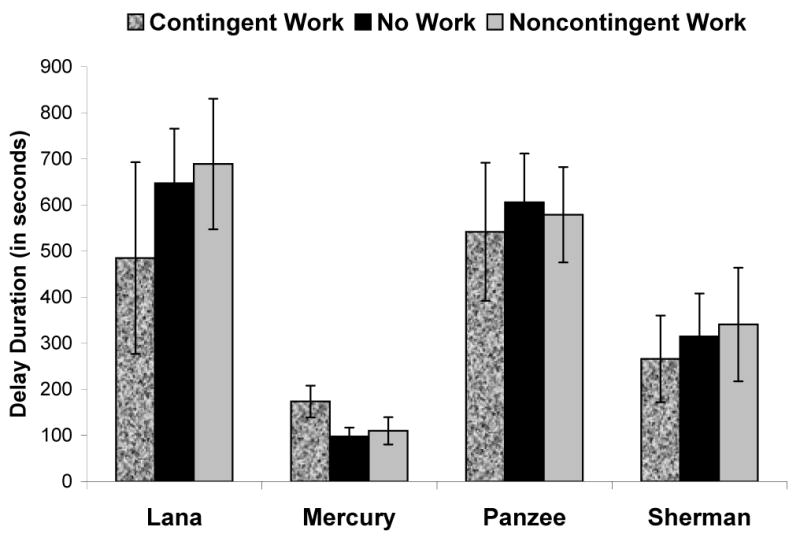
The mean duration of delay maintenance in each of the three conditions by each chimpanzee. Bars indicate 95% confidence intervals.
Finally, there was no relationship between the percentage of time each chimpanzee spent working on the concurrent task and the number of rewards items obtained in the noncontingent condition, Lana r = 0.119, df = 8, p = 0.74, Mercury r = 0.155, df = 8, p = 0.67, Panzee r = 0.095, df = 8, p = 0.79, Sherman r = -0.389, df = 8, p = 0.26.
Discussion
This experimental paradigm provided an assessment of the influence of concurrent task activity and response-reward contingency on chimpanzee self-control. Importantly, all chimpanzees spent at least some time working on the concurrent task even when it was not linked to the delivery of rewards. This made the noncontingent work condition truly different from the no work condition. We also can conclude that the chimpanzees' work on the noncontingent task was not the result of confusion between task conditions, given that: 1.) The noncontingent task was presented on a cart that was clearly spatially separate from the reward dispenser and the chimpanzees had never received rewards from a spatially separate computer cart in the past; and 2.) All four chimpanzees spent a similar amount of time on the noncontingent task whether it followed the contingent condition or no work condition within the 3-sessions block order.
Despite the validity of this method, three of four chimpanzees showed no significant increase or decrease in delay maintenance across the three conditions. Although, on average, they seemed to perform poorer in the contingent task condition compared to the other two conditions, their behavior varied greatly within conditions, and this apparent trend was not supported by statistical analysis. Further, there was no relationship between the length of time they spent working on the noncontingent concurrent task and the number of delayed rewards they obtained in that condition to support this apparently negative influence of concurrent task work on their delay maintenance. Had they exhibited such a pattern, this would have given support to the hypothesis that the response requirement provided an additional self-regulatory load to already having to wait to consume candies, which in turn reduced the chimpanzees' overall ability to delay gratification. However, working to accumulate food rewards was no different than passively waiting for those rewards to accumulate for those three individuals. Nevertheless, this is a marked difference from the reported performance of human children (Peake et al., 2002).
For two of the chimpanzees (Lana and Panzee), the lack of an effect of work on delay maintenance might have been the result of their overall high levels of self-control in this task. Had we presented a more challenging baseline delay task, these individuals may have produced clearer behavior patterns (though not necessarily in the same direction as the apparent trend exhibited in this experiment). On the other hand, Sherman clearly had room to improve in terms of the number of items he could obtain (see Figure 1). He did not, however, show any effects of having a concurrent task or having that task performance tied to delivery of the accumulated rewards.
The chimpanzee Mercury showed an entirely different result. This animal accumulated the most rewards when the accumulation of those rewards was contingent upon his task responding. He did not show any elevation in performance (compared to the no task condition) when he had access to the computer task but the reward delivery rate was not tied to his work, and there was no relationship between the percentage of time he spent working in the noncontingent condition and the quantity of rewards he obtained in this condition. This pattern of performance indicates that the contingent relation between Mercury's computer work and the delivery of rewards allowed him to delay gratification for longer than in the baseline (no task) condition. It was not simply the presence of the potentially distracting concurrent task because Mercury delayed longer when his task performance was linked directly to reward delivery than when the task was independent of reward delivery. Given the highly observable contingency between the completion of each concurrent task trial and the delivery of each delayed reward item in this paradigm, Mercury may have perceived some level of control over the delay scenario that enhanced his delay maintenance performance. Indeed, perceived control over task parameters has led to positive changes in performance by nonhuman primates in other test paradigms. For example, Washburn, Hopkins, and Rumbaugh (1990) reported that rhesus monkeys (Macaca mulatta) that were allowed to select computer tasks from a menu before completing those tasks showed better individual task performance than when the same tasks were assigned by the experimenter. Capuchin monkeys (Cebus apella) showed some similar benefits of perceived control when selecting the order in which they completed a set of required tasks compared to when task completion order was out of their control (Beran et al., 2007).
Mercury's results contrast with the results of Peake et al. (2002) who tested children in a similar situation. The children in that study delayed gratification longest when they had access to a concurrent task, but they performed equally well whether their task performance was or was not linked to the delivery of rewards. Thus, the distracting nature of the task allowed the children to delay longer. However, it is important to note that there were methodological differences between these studies that may have accounted for the behavioral differences. The chimpanzees were tested in a repeated-measures design whereas the children were tested with a between subjects design. Also, the children of the Peake et al. (2002) study were only told that receiving the reward later was contingent upon their concurrent task activity, whereas Mercury could observe a portion of the rewards being delivered each time he completed a computer trial. Thus, the response-reward contingency was potentially more apparent to Mercury than it was to the children. So, if this were the underlying cause for Mercury's performance pattern, it might help explain why he did not perform more similarly to the previously tested human children.
Interestingly, Mercury showed the overall lowest levels of self-control of all the chimpanzees tested in the present study. This was true not only in the present study but also in previous investigations of delay maintenance in these chimpanzees (Beran, 2002; Beran & Evans, 2006; Evans & Beran, 2007). This suggests that there may be individual differences not only in general self-control ability but also with regard to the influence of response-reward contingency on chimpanzees' delayed gratification capacity. For chimpanzees that were good at delaying gratification, there seemed to be no difference in how many candies they obtained across the conditions. However, for the one chimpanzee that was a poor performer in terms of delay maintenance, having a task to work on and having that work directly linked to reward delivery apparently increased delay maintenance. We hypothesize that this may occur because individuals with low self-control benefit more from attendant forms of control over the environment that may facilitate internal self-control mechanisms. Given the small number of chimpanzees tested in this investigation, it is important to replicate this performance pattern with other animals across variable degrees of overall self-control capacity. Future studies should also focus on the apparent difference between chimpanzees and human children with regard to the relation between working/waiting for reward delivery and self-control. More extensive analyses of these behavior patterns will be informative with regard to the emergence and degree of self-control in humans and other species.
Acknowledgments
The authors thank Sarah Hunsberger and John Kelley for their assistance during experimental trials. This research was supported by grant HD-38051 from the National Institute of Child Health and Human Development.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Beran MJ. Maintenance of self-imposed delay of gratification by four chimpanzees (Pan troglodytes) and an orangutan (Pongo pygmaeus) J Gen Psychol. 2002;129:49–66. doi: 10.1080/00221300209602032. [DOI] [PubMed] [Google Scholar]
- Beran MJ, Evans TA. Maintenance of delay of gratification by four chimpanzees (Pan troglodytes): The effects of delayed reward visibility, experimenter presence, and extended delay intervals. Behav Process. 2006;73:315–324. doi: 10.1016/j.beproc.2006.07.005. [DOI] [PubMed] [Google Scholar]
- Beran MJ, Klein ED, Evans TA, Chen B, Antworth B. Perceived control, motivation, and task performance in capuchin monkeys. In: Zelick PR, editor. Issues in the Psychology of Motivation. Hauppauge, NY: Nova Science Publishers; 2007. pp. 171–185. [Google Scholar]
- Beran MJ, Savage-Rumbaugh ES, Pate JL, Rumbaugh DM. Delay of gratification in chimpanzees (Pan troglodytes) Dev Psychobiol. 1999;34:119–127. doi: 10.1002/(sici)1098-2302(199903)34:2<119::aid-dev5>3.0.co;2-p. [DOI] [PubMed] [Google Scholar]
- Chelonis JJ, Logue AW, Sheehy R, Mao J. Effects of response effort on self-control in rats. Anim Learn Behav. 1998;26:408–415. [Google Scholar]
- Clement TS, Feltus JR, Kaiser DH, Zentall TR. “Work ethic” in pigeons: Reward value is directly related to the effort or time required to obtain the reward. Psychon Bull Rev. 2000;7:100–106. doi: 10.3758/bf03210727. [DOI] [PubMed] [Google Scholar]
- Cournoye M, Trudel M. Behavioral correlates of self-control at 33 months. Infant Behav Devel. 1991;14:497–503. [Google Scholar]
- Evans TA. Performance in a computerized self-control task by rhesus macaques (Macaca mulatta): The combined influence of effort and delay. Learn Motiv. 2007;38:342–357. doi: 10.1016/j.lmot.2007.02.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evans TA, Beran MJ. Chimpanzees use self-distraction to cope with impulsivity. Bio Lett-UK. 2007a;3:599–602. doi: 10.1098/rsbl.2007.0399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evans TA, Beran MJ. Delay of gratification and delay maintenance by rhesus macaques (Macaca mulatta) J Gen Psychol. 2007b;134:199–216. doi: 10.3200/GENP.134.2.199-216. [DOI] [PubMed] [Google Scholar]
- Evans TA, Westergaard GC. Self-control and tool use in tufted capuchin monkeys (Cebus apella) J Comp Psychol. 2006;120:163–166. doi: 10.1037/0735-7036.120.2.163. [DOI] [PubMed] [Google Scholar]
- Grosch J, Neuringer A. Self-control in pigeons under the Mischel paradigm. J Exp Anal Behav. 1981;35:3–21. doi: 10.1901/jeab.1981.35-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grossbard CL, Mazur JE. A comparison of delays and ratio requirements in self-control choice. J Exp Anal Behav. 1986;45:305–315. doi: 10.1901/jeab.1986.45-305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Logue AW. Research on self-control: An integrating framework. Behav Brain Sci. 1988;11:665–709. [Google Scholar]
- Logue AW, Forzano LB, Ackerman KT. Self-control in children: Age, preference for reinforcer amount and delay, and language ability. Learn Motiv. 1996;27:260–277. [Google Scholar]
- Miller DT, Karniol R. Coping strategies and attentional mechanisms in self-imposed and externally imposed delay situations. J Pers Soc Psychol. 1976;34:310–316. [Google Scholar]
- Mischel W. Processes in delay of gratification. In: Berkowitz L, editor. Advances in Experimental Social Psychology. New York: Academic Press; 1974. pp. 249–292. [Google Scholar]
- Mischel W. Objective and subjective rules for delay of gratification. In: d'Ydewalle G, Lens W, editors. Cognition in Human Learning and Motivation. Hillsdale, NJ: Erlbaum; 1981. pp. 33–58. [Google Scholar]
- Mischel W, Baker N. Cognitive appraisals and transformations in delay behavior. J Pers Soc Psychol. 1975;31:254–261. [Google Scholar]
- Mischel W, Ebbesen EB. Attention in delay of gratification. J Pers Soc Psychol. 1970;16:329–337. doi: 10.1037/h0032198. [DOI] [PubMed] [Google Scholar]
- Mischel W, Ebbesen EB, Zeiss A. Cognitive and attentional mechanisms in delay of gratification. J Pers Soc Psychol. 1972;21:204–218. doi: 10.1037/h0032198. [DOI] [PubMed] [Google Scholar]
- Mischel W, Shoda Y, Rodriguez ML. Delay of gratification in children. Science. 1989;244:933–938. doi: 10.1126/science.2658056. [DOI] [PubMed] [Google Scholar]
- Patterson CJ, Carter DB. Attentional determinants of children's self-control in waiting and working situations. Child Dev. 1979;50:272–275. [Google Scholar]
- Peake PK, Hebl M, Mischel W. Strategic attention deployment for delay of gratification in working and waiting situations. Dev Psychol. 2002;38:313–326. doi: 10.1037//0012-1649.38.2.313. [DOI] [PubMed] [Google Scholar]
- Rachlin H. The Science of Self-control. Cambridge, MA: Harvard University Press; 2000. [Google Scholar]
- Stevens JR, Rosati AG, Ross KR, Hauser MD. Will travel for food: Spatial discounting in two New World monkeys. Curr Biol. 2005;15:1855–1860. doi: 10.1016/j.cub.2005.09.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tobin H, Logue AW, Chelonis JJ, Ackerman KT, May JG. Self-control in the monkey Macaca fascicularis. Anim Learn Behav. 1996;24:168–174. [Google Scholar]
- Toner IJ, Lewis BC, Gribble CM. Evaluative verbalization and delay maintenance behavior in children. J Exp Child Psychol. 1979;28:205–210. [Google Scholar]
- Toner IJ, Smith RA. Age and overt verbalization in delay-maintenance behavior in children. J Exp Child Psychol. 1977;24:123–128. [Google Scholar]
- van Haaren F, van Hest A, Van de Poll NE. Self-control in male and female rats. J Exp Anal Behav. 1988;49:201–211. doi: 10.1901/jeab.1988.49-201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Washburn DA, Hopkins WD, Rumbaugh DM. Perceived control in rhesus monkeys (Macaca mulatta): Enhanced video-task performance. J Exp Psychol Anim Behav Proc. 1991;17:123–129. doi: 10.1037//0097-7403.17.2.123. [DOI] [PubMed] [Google Scholar]