

Abstract
A sex difference on mental-rotation tasks has been demonstrated repeatedly, but not in children less than 4 years of age. To demonstrate mental rotation in human infants, we habituated 5-month-old infants to an object revolving through a 240° angle. In successive test trials, infants saw the habituation object or its mirror image revolving through a previously unseen 120° angle. Only the male infants appeared to recognize the familiar object from the new perspective, a feat requiring mental rotation. These data provide evidence for a sex difference in mental rotation of an object through three-dimensional space, consistently seen in adult populations.
Fifty years of research has confirmed that men typically outperform women in spatial-ability tests (Linn & Petersen, 1985; Voyer, Voyer, & Bryden, 1995). Such sex differences have been detected in 4-year-old children (Levine, Huttenlocher, Taylor, & Langrock, 1999); in children under 13, these differences are most often found on tasks requiring mental rotation (Voyer et al., 1995). Mental-rotation tasks revealing the largest sex differences require subjects to view a two-dimensional (2-D) representation of a three-dimensional (3-D) object, and to then recognize a novel 2-D representation of the same object rotated into a different orientation in 3-D space (Levine et al., 1999). Effect sizes for sex differences on such tasks are typically larger than the effects of sex on other behaviors (Collaer & Hines, 1995).
The current study examined the origins of mental rotation in infancy. Early work revealed that 4-month-old infants can detect the 3-D form of objects moving around two axes of rotation (Kellman, 1984; Kellman & Short, 1987). More recent studies presented infants with kinetic random-dot displays that specified rotating 3-D cubes (Arterberry & Yonas, 2000) and with video displays of partially occluded 3-D shapes rotating around a vertical axis (Johnson, Cohen, Marks, & Johnson, 2003); in both cases, 2-month-old infants appeared to perceive the 3-D shape of rotating objects.
Other studies found that 4-month-old infants form dynamic mental representations that allow them to both track the movement of a 2-D object rotating in the frontal plane and anticipate the object’s ultimate orientation (Rochat & Hespos, 1996; Hespos & Rochat, 1997). These results were interpreted as tentative evidence for rudimentary mental rotation in infants; however, although these results suggest that infants can use exposure to a moving 2-D object to help them predict how that object will look when rotated in a 2-D plane, full-blown mental rotation has traditionally been tested by requiring observers to mentally rotate 3-D stimuli through 3-D space and to discriminate the rotated object from its mirror image (Shepard & Cooper, 1982). No studies have yet provided evidence that infants can recognize a particular 3-D object—as distinct from its mirror image—after it has been rotated through 3-D space into a previously unseen perspective; such an ability would be indicative of mental rotation. Nor have any studies of infant perception of rotating objects documented sex differences.
Here, we evaluate the hypothesis that infants can mentally rotate visual stimuli through 3-D space and investigate possible sex differences in performance. We presented 5-month-old infants with a video representation of a 3-D habituation object (Fig. 1) revolving around the vertical axis in 3-D space, through a 240° angle. We hypothesized that infants would recognize the object in subsequent test trials in which it revolved through a previously unseen 120° angle. After habituation, each infant saw a series of alternating test trials presenting the original habituation object or its mirror image; in both cases, the test objects were shown revolving through the previously unseen angle.
Fig. 1.
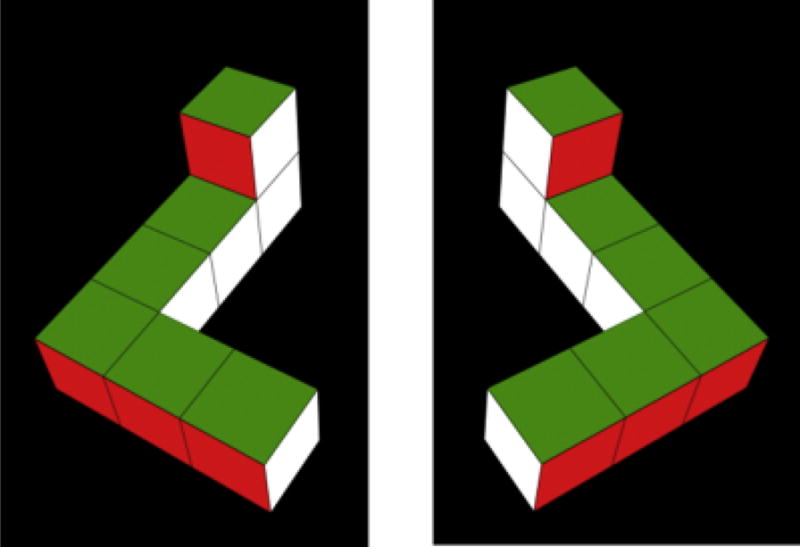
Shepard-Metzler object (Shepard & Metzler, 1971), which we arbitrarily called the L-object, and its mirror image, the R-object, pictured on the left and right, respectively.
We reasoned that evidence for mental rotation would be revealed in the test trials by a preference for the mirror-image object; such a preference would imply recognition that the other test object is the original habituation object, now rotating through a novel angle. A preference for the mirror-image stimulus would be inexplicable in terms of lower-order stimulus characteristics (e.g., brightness, amount of contour, etc.) because the test stimuli were identical in all respects other than their enantiomorphic properties. Recognizing the habituation object from the new perspective would require infants to rotate a mental representation of either the habituation object or the visible test stimulus.
METHOD
We tested 20 male and 20 female full-term 5-month-old infants (mean age =153.2 days, SD = 10). Five additional infants were observed but excluded from the analysis, four due to fussiness, and one due to sleepiness.
Each infant was tested individually while sitting on a parent’s lap in a darkened room, 100 cm from a 53-cm monitor screen, on which all stimuli were presented. A Macintosh G5 running Habit software (Cohen, Atkinson, & Chaput, 2002) presented stimuli, timed trials, calculated the habituation criterion, and stored data. A trained observer, not visible to the infant and blind to the stimulus shown, viewed the infant via a closed-circuit camera and used the computer’s keyboard to initiate trials and record the infant’s fixations and their durations. Parents kept their eyes closed throughout the procedure.
The stimuli were 2-D video representations of unfamiliar 3-D objects presented on a black background. Maximum vertical and horizontal dimensions of the objects during rotation were 15.5° and 12.2° of visual angle, respectively. The stimuli represented a simplified Shepard-Metzler object (Shepard & Metzler, 1971), which we arbitrarily called the L-object, and its mirror image, the R-object. Each stimulus object was constructed of seven cubes attached rigidly with 90° bends at its top and bottom; a two-cube bar (x-axis) was attached at the bottom of a straight central bar formed of four cubes (y-axis), and a single-cube bar (z-axis) was attached to the top of this central bar. If viewed from above, all visible faces of the objects were yellow; if viewed from below, all visible faces were red. Viewed from the front, right, back, and left, the faces were purple, blue, white, and green, respectively.
Two habituation videos and two test videos were used. The habituation videos presented the L- or R-object rotating at 48° per second around the vertical axis, through a 240° arc. On reaching its maximum extent of rotation, the object rotated back to its starting point. The stimuli were looped, so the image rotated continuously back and forth. The L- and R-test videos continued the rotation of the L- and R-objects, respectively, through the previously unseen 120° of arc; like the habituation stimuli, the test stimuli continuously rotated back and forth. No still frame of the habituation stimuli matched any still frame of the test stimuli.
Trials were preceded by the presentation of an attention-getter, which drew the infant’s attention to the screen. The observer began each trial with a key press. Trials ended (i.e., the stimulus was terminated) either 2 s after the observer released a key to indicate that the infant was no longer fixating the display, or after 60 s. If the infant returned attention to the stimulus in the 2-s interval, the trial continued. Otherwise, the attention-getter appeared between trials; when the infant looked, a new trial was initiated.
Each infant initially saw a series of identical habituation trials presenting the L- or R-habituation stimulus; infants were randomly assigned to the L- or R-habituation group. Average fixation duration during the first four habituation trials was computed on-line for each infant, and habituation was evaluated by comparing this average with fixation durations averaged across succeeding four-trial blocks. Infants were deemed habituated when fixation of the habituation stimuli declined in a given four-trial block to 50% of looking in the first four trials. Test trials were presented after infants habituated, or after 12 habituation trials, whichever came first.
Each infant then saw a series of six test trials. Twenty infants were randomly assigned to see the L-test stimulus in the first test trial; the others saw the R-test stimulus first. The test stimulus presented in subsequent trials was alternated thereafter.
RESULTS
Male 5-month-old infants looked longer at the mirror-image test object than at the familiar object rotating through the novel angle, t(19) =4.07, p < .001, prep = .99, d = 0.61 (Fig. 2); 70% of these infants preferred the mirror-image test stimulus (Wilcoxon Z = 3.02, p < .003, prep = .98). In contrast, female 5-month-old infants looked at the test stimuli about equally, t(19) = 0.35, p = .73, prep = .33, d = 0.06; 45% of these infants preferred the mirror-image stimulus (Wilcoxon Z = 0.26, p = .79, prep = .28). Additionally, the male infants’ novelty preference was significantly greater than that of the females, t(38) = 2.07, p = .045, prep = .89, d = 0.66. Male and female infants did not differ in accumulated habituation times, t(38) = 1.60, p = .12, prep = .80, d = 0.50, or in number of trials to habituation, t(38) = 0.86, p = .39, prep = .58, d = 0.27.
Fig. 2.
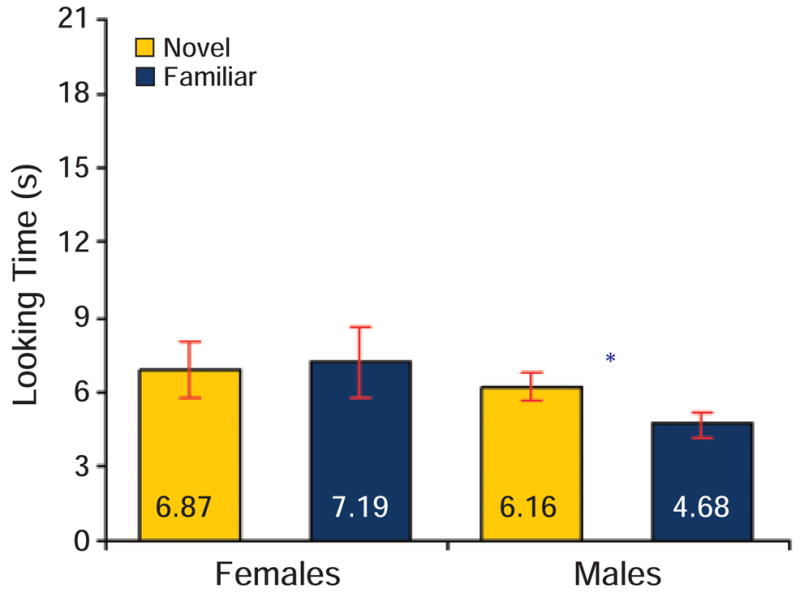
Five-month-old infants’ mean looking times at novel and familiar stimuli. Error bars indicate standard errors of the means, and an asterisk indicates statistical significance, p < .001, two-tailed t test.
In summary, whereas female infants treated the test stimuli similarly, male infants treated the habituation and familiar test objects similarly but discriminated between the habituation and mirror-image test objects.
DISCUSSION
Only the male infants discriminated the habituation object from its mirror image. Although infants can fail to demonstrate discrimination for many reasons, the males’ attention to the mirror-image stimulus permits the inference that they recognized that the other stimulus represented the familiar habituation object, now rotating through a novel angle. Such recognition requires mental rotation of a mental representation of the habituation object (allowing comparison to the visible test stimulus), of the visible test stimulus (allowing comparison to a mental representation of the habituation object), or of both. Thus, these findings represent the first demonstration of mental rotation of an object through 3-D space by human infants. Quinn and Liben (2008), likewise, report mental rotation in 3- to 4-month-old male infants; their stimuli were static drawings of a 2-D object rotated in a 2-D (frontal) plane.
These data provide evidence for the early appearance of a sex difference in cognition consistently seen in older human populations (Levine et al., 1999; Linn & Petersen, 1985; Voyer et al., 1995). Some theorists have argued that sex differences in spatial cognition reflect selective evolutionary pressures (Ecuyer-Dab & Robert, 2004; Gaulin & FitzGerald, 1986), but regardless, all phenotypes emerge in ontogeny (Lickliter & Berry, 1990). Consequently, understanding the roots of this difference in development will be important, if challenging.
Multiple factors contribute to sex differences in spatial cognition, including the effects of gonadal hormones, sex differences in maturation rates of the brain’s hemispheres, and experiences with particular stimuli and tasks. Androgen levels are related to mental-rotation ability in older populations, having both organizational and activational influences on its development. Prior exposure to androgens during gestation is positively associated with 7- to 12-year-old girls’ performances on spatial reasoning tasks (Grimshaw, Sitarenios, & Finegan, 1995; Hampson, Rovet, & Altmann, 1998); these hormones appear to contribute to spatial abilities by influencing the organization of the developing nervous system in utero. Likewise, normal variations across the menstrual cycle in testosterone and estradiol levels are correlated with women’s performance on mental-rotation tasks, suggesting that these hormones also influence spatial ability via an activating role in the nervous system (Hausmann, Slabbekoorn, Van Goozen, Cohen-Kettenis, & Güntürkün, 2000; but see Schöning et al., 2007). Finally, in a double-blind placebo-controlled study, one sublingual dose of 0.5 mg of testosterone significantly improved healthy young women’s performances on a standard mental-rotation task (Aleman, Bronk, Kessels, Koppeschaar, & van Honk, 2004).
By 3 months after birth, male and female brains differ structurally (de Lacoste, Horvath, & Woodward, 1991) and functionally (Shucard, Shucard, Cummins, & Campos, 1981), and male and female infants have already experienced a social world that treats them differently (Donovan, Taylor, & Leavitt, 2007; Stern & Karraker, 1989). It is unknown what experiential factors contribute to the development of mental-rotation ability in infants, but studies of older populations have confirmed that experience influences performance on spatial-ability tests. A meta-analysis addressing this question found that participation in activities requiring spatial competence was associated with improved spatial abilities, and that training improves performance on spatial-ability tests for males and females (Baenninger & Newcombe, 1989). In addition, individuals who prefer sports that provide opportunities to exercise spatial skills outperform other individuals on mental-rotation tasks (Voyer, Nolan, & Voyer, 2000), and training experiences with 2-D representations of 3-D objects can improve mental-rotation performance (Sanz de Acedo Lizarraga & García Ganuza, 2003).
The precise roles of the factors that contribute to the development of mental-rotation ability must still be determined. Explaining the emergence of a sex difference in this ability by 5 months of age will require research specifically designed to explore how these factors interact to influence the development of mental-rotation performance.
Acknowledgments
This work was supported by Pitzer College’s R&A Committee (D.S.M.) and National Institutes of Health Grants R01-HD40432 and R01-HD48733 (S.P.J.). We thank our participants, and Dona Bailey, Robert Ditto, and Dawn Jones for invaluable help with this work.
References
- Aleman A, Bronk E, Kessels RPC, Koppeschaar HPF, van Honk J. A single administration of testosterone improves visuospatial ability in young women. Psychoneuroendocrinology. 2004;29:612–617. doi: 10.1016/S0306-4530(03)00089-1. [DOI] [PubMed] [Google Scholar]
- Arterberry ME, Yonas A. Perception of three-dimensional shape specified by optic flow by 8-week-old infants. Perception & Psychophysics. 2000;62:550–556. doi: 10.3758/bf03212106. [DOI] [PubMed] [Google Scholar]
- Baenninger M, Newcombe N. The role of experience in spatial test performance: A meta-analysis. Sex Roles. 1989;20:327–344. [Google Scholar]
- Cohen LB, Atkinson DJ, Chaput HH. Habit 2002: A new program for obtaining and organizing data in infant perception and cognition studies (Version 1.0) [Computer software] Austin: University of Texas; 2002. [Google Scholar]
- Collaer ML, Hines M. Human behavioral sex differences: A role for gonadal hormones during development? Psychological Bulletin. 1995;118:55–107. doi: 10.1037/0033-2909.118.1.55. [DOI] [PubMed] [Google Scholar]
- de Lacoste MC, Horvath DS, Woodward DJ. Possible sex-differences in the developing human fetal brain. Journal of Clinical and Experimental Neuropsychology. 1991;13:831–846. doi: 10.1080/01688639108405101. [DOI] [PubMed] [Google Scholar]
- Donovan W, Taylor N, Leavitt L. Maternal sensory sensitivity and response bias in detecting change in infant facial expressions: Maternal self-efficacy and infant gender labeling. Infant Behavior and Development. 2007;30:436–452. doi: 10.1016/j.infbeh.2006.12.008. [DOI] [PubMed] [Google Scholar]
- Ecuyer-Dab I, Robert M. Spatial ability and home-range size: Examining the relationship in Western men and women (Homo sapiens) Journal of Comparative Psychology. 2004;118:217–231. doi: 10.1037/0735-7036.118.2.217. [DOI] [PubMed] [Google Scholar]
- Gaulin SJC, FitzGerald RW. Sex differences in spatial ability: An evolutionary hypothesis and test. American Naturalist. 1986;127:74–88. [Google Scholar]
- Grimshaw GM, Sitarenios G, Finegan JK. Mental rotation at 7 years: Relations with prenatal testosterone levels and spatial play experiences. Brain and Cognition. 1995;29:85–100. doi: 10.1006/brcg.1995.1269. [DOI] [PubMed] [Google Scholar]
- Hampson E, Rovet JF, Altmann D. Spatial reasoning in children with congenital adrenal hyperplasia due to 21-hydroxylase deficiency. Developmental Neuropsychology. 1998;14:299–320. [Google Scholar]
- Hausmann M, Slabbekoorn D, Van Goozen SHM, Cohen-Kettenis PT, Güntürkün O. Sex hormones affect spatial abilities during the menstrual cycle. Behavioral Neuroscience. 2000;114:1245–1250. doi: 10.1037//0735-7044.114.6.1245. [DOI] [PubMed] [Google Scholar]
- Hespos SJ, Rochat P. Dynamic mental representation in infancy. Cognition. 1997;64:153–188. doi: 10.1016/s0010-0277(97)00029-2. [DOI] [PubMed] [Google Scholar]
- Johnson SP, Cohen LB, Marks KH, Johnson KL. Young infants’ perception of object unity in rotation displays. Infancy. 2003;4:285–295. [Google Scholar]
- Kellman PJ. Perception of three-dimensional form by human infants. Perception & Psychophysics. 1984;36:353–358. doi: 10.3758/bf03202789. [DOI] [PubMed] [Google Scholar]
- Kellman PJ, Short KR. Development of three-dimensional form perception. Journal of Experimental Psychology: Human Perception and Performance. 1987;13:545–557. doi: 10.1037//0096-1523.13.4.545. [DOI] [PubMed] [Google Scholar]
- Levine SC, Huttenlocher J, Taylor A, Langrock A. Early sex differences in spatial skill. Developmental Psychology. 1999;35:940–949. doi: 10.1037//0012-1649.35.4.940. [DOI] [PubMed] [Google Scholar]
- Lickliter R, Berry TD. The phylogeny fallacy: Developmental psychology’s misapplication of evolutionary theory. Developmental Review. 1990;10:348–364. [Google Scholar]
- Linn MC, Petersen AC. Emergence and characterization of sex differences in spatial ability: A meta-analysis. Child Development. 1985;56:1479–1498. [PubMed] [Google Scholar]
- Quinn PC, Liben LS. A sex difference in mental rotation in young infants. Psychological Science. 2008;19:1067–1070. doi: 10.1111/j.1467-9280.2008.02201.x. [DOI] [PubMed] [Google Scholar]
- Rochat P, Hespos SJ. Tracking and anticipation of invisible spatial transformations by 4- to 8-month-old infants. Cognitive Development. 1996;11:3–17. [Google Scholar]
- Sanz de Acedo Lizarraga ML, García Ganuza JM. Improvement of mental rotation in girls and boys. Sex Roles. 2003;49:277–286. [Google Scholar]
- Schöning S, Engelien A, Kugel H, Schäfer S, Schiffbauer H, Zwitserlood P, et al. Functional anatomy of visuo-spatial working memory during mental rotation is influenced by sex, menstrual cycle, and sex steroid hormones. Neuropsychologia. 2007;45:3203–3214. doi: 10.1016/j.neuropsychologia.2007.06.011. [DOI] [PubMed] [Google Scholar]
- Shepard RN, Cooper LA. Mental images and their transformations. Cambridge, MA: MIT Press; 1982. [Google Scholar]
- Shepard RN, Metzler J. Mental rotation of three-dimensional objects. Science. 1971;171:701–703. doi: 10.1126/science.171.3972.701. [DOI] [PubMed] [Google Scholar]
- Shucard JL, Shucard DW, Cummins KR, Campos JJ. Auditory evoked potentials and sex-related differences in brain development. Brain and Language. 1981;13:91–102. doi: 10.1016/0093-934x(81)90131-0. [DOI] [PubMed] [Google Scholar]
- Stern M, Karraker KH. Sex stereotyping of infants: A review of gender labeling studies. Sex Roles. 1989;20:501–522. [Google Scholar]
- Voyer D, Nolan C, Voyer S. The relation between experience and spatial performance in men and women. Sex Roles. 2000;43:891–915. [Google Scholar]
- Voyer D, Voyer S, Bryden MP. Magnitude of sex differences in spatial abilities: A meta-analysis and consideration of critical variables. Psychological Bulletin. 1995;117:250–270. doi: 10.1037/0033-2909.117.2.250. [DOI] [PubMed] [Google Scholar]