

Abstract
Choline is an important metabolite in all cells due to the major contribution of phosphatidylcholine to the production of membranes, but it takes on an added role in cholinergic neurons where it participates in the synthesis of the neurotransmitter acetylcholine. We have cloned a suppressor for a yeast choline transport mutation from a Torpedo electric lobe yeast expression library by functional complementation. The full-length clone encodes a protein with 10 putative transmembrane domains, two of which contain transporter-like motifs, and whose expression increased high-affinity choline uptake in mutant yeast. The gene was called CTL1 for its choline transporter-like properties. The homologous rat gene, rCTL1, was isolated and found to be highly expressed as a 3.5-kb transcript in the spinal cord and brain and as a 5-kb transcript in the colon. In situ hybridization showed strong expression of rCTL1 in motor neurons and oligodendrocytes and to a lesser extent in various neuronal populations throughout the rat brain. High levels of rCTL1 were also identified in the mucosal cell layer of the colon. Although the sequence of the CTL1 gene shows clear homology with a single gene in Caenorhabditis elegans, several homologous genes are found in mammals (CTL2–4). These results establish a new family of genes for transporter-like proteins in eukaryotes and suggest that one of its members, CTL1, is involved in supplying choline to certain cell types, including a specific subset of cholinergic neurons.
Cells contain large amounts of choline incorporated in their membranes, and all plant and animal cells, including unicellular organisms, absorb free choline as a nutrient. At cholinergic nerve terminals, the sodium-dependent high affinity choline uptake mechanism that is coupled to acetylcholine synthesis has been particularly well characterized at the functional level (1–3) but has thus far eluded diverse attempts at identification based on protein purification (4), even in conjunction with the use of a selective and irreversible ligand (5).
The yeast choline transporter has been isolated by homologous complementation by using a choline transport deficient yeast strain (6), and heterologous complementation of yeast mutants with a yeast expression library made with Arabidopsis thaliana cDNA has been used to identify the potassium transporter of plants (7). We decided to adapt the strategy of complementation cloning to the problem of neuronal choline transport by using choline transport deficient yeast with the highly cholinergic electric lobes of Torpedo as a source of cDNA. However, the single clone isolated, capable of partially restoring choline uptake in the mutant yeast, did not resemble the yeast choline transporter (8) and is thus considered to be a heterologous suppressor for the choline transport mutation. We report the sequences of genes both homologous and orthologous to the Torpedo choline transporter-like protein, tCTL1, and we begin the characterization of this new family of transmembrane proteins by studying the cellular distribution of CTL1 in the rat and its chromosomal localization in humans.
Materials and Methods
Complementation of the Choline Transport Mutation in Yeast.
The yeast strain (ctr ise URA3Δ) used in this study was obtained by crossing D308–14D (ctr ise leu2 his4) (6) with a URA3Δ strain (9). This strain will not grow in uracil-depleted medium, nor in high myoinositol unless both external choline and a choline uptake mechanism are made available. poly(A)+RNA was prepared from frozen electric lobes from Torpedo marmorata. The corresponding cDNAs prepared using the cDNA cloning kit from Amersham were inserted in the BstXI site of the plasmid pFL61 (9). Transformation of yeast was done by using an improved lithium acetate method (10). Transformed yeast were grown on solid medium under selective growth conditions (no uracil/20 μg/ml choline/20 μg/ml myoinositol); ise revertants were eliminated as being capable of growth in the absence of choline. After elimination of ctr revertants and yeast suppressor mutations by transformation of mutant yeast with isolated plasmids, five clones were retained and tested for choline uptake.
Choline Uptake by Pools of Transformed Yeast.
Aliquots of mutant yeast were transformed with individual isolated plasmids and, following amplification, tested for choline uptake in nitrogen-free medium (11) at 30°C for 10 or 30 min with 25 nM [3H]choline (0.1 μCi, 2.8 TBq/mmol, Amersham) and with the indicated concentrations of unlabeled choline and hemicholinium-3 (HC-3, Sigma), then filtered, and washed. The apparent Kt in the presence of the competitive inhibitor HC-3, determined by nonlinear regression, was used to calculate the inhibitory constant Ki as follows: Ki = [I]/[(Ktapp/Kt) − 1].
Cloning and Sequence Analysis of CTL1 Orthologs and Homologs.
Full-length cDNAs for the electric organ suppressor of the yeast choline transport mutation, tCTL1, and its rat ortholog, rCTL1, were isolated from λZAP II (Stratagene) libraries constructed from the T. marmorata electric lobe and rat brain, respectively. The complete sequence of a 4.4-kb tCTL1 clone and a 2.9-kb rCTL1 clone were obtained by using both T7 Sequenase (Amersham) and an external sequencing service (ESGS-Cybergene, Evry). The predicted transmembrane domains (TMDs) of CTL proteins were assigned based on Kyte and Doolittle hydropathy plots. Homologous expressed sequence tags (ESTs) were gathered using blast programs. Sequences for the coding regions of the human ortholog, hCTL1 (Unigene accession no. Hs.179902), and its homolog hCTL2 (Unigene accession no.: Hs.167515 and Hs.105509) were completed by sequencing PCR products obtained using cDNA from the Ewing's sarcoma cell line, ICB 112 (12). Other full-length coding sequences were available as conceptual translations of genomic sequence from the MHC III region: NG22 from mouse (GenBank accession no. AAC84166) and human (GenBank accession no. AAD21813) are renamed here, respectively, mCTL4 and hCTL4; F35C8.7 from C. elegans; YOR161c from yeast; and F20M13.200 from A. thaliana. The alignment of selected sequences was done using gcg:pileup. EST analysis further indicated CTL1 homologs in Drosophila (GenBank accession no. AA697340), Dictyostelium (GenBank accession no. C24622), and another human homolog that we have called hCTL3 (GenBank accession no. Aa329432) and its equivalent in the mouse, mCTL3 (GenBank accession no. W64177). The clustering relationships between these proteins are presented as a dendrogram drawn using clustalw.
Northern Blot Analysis for rCTL1.
poly(A)+RNA (2 μg) extracted from various rat tissues were analyzed by Northern blot using a 5′-BamHI/EcoRI fragment from rCTL1 labeled with [32P]dCTP and then washed at high stringency. The blots were exposed for 7 then 40 (not shown) hr and reprobed with β-actin.
In Situ Hybridization (ISH).
ISH experiments were done as described (13, 14). A 1.6-kb PstI restriction fragment derived from rCTL1 was subcloned into pGEM-4Z and used to synthesize digoxigenin-labeled antisense and sense riboprobes. The double ISH protocol (15) was done using a fluorescein-labeled choline acetyltransferase riboprobe encoding a fragment corresponding to nucleotides 1–2,332 (16). The rCTL1 sense riboprobe gave no specific signal, whereas the antisense cRNA probe led to reaction products observed in a cytoplasmic rim around the cell nucleus.
Positioning of hCTL1 on Chromosome 9 Using Yeast Artificial Chromosomes (YACs).
A sequence tag site (WI-17320) has recently been linked to an EST corresponding to the 3′-coding region of hCTL1. YACs within the indicated region, 9q22/31, were obtained from the Centre d'Etudes du Polymorphisme Humain (Paris). The yeast harboring the YACs were used for PCR analysis. PCR primers for WI-17320 were as given (GenBank accession no. G24229), and for yeast actin (GenBank accession no. X61502) were forward: CAAAA TTGGC TAGAG AAACA ACCG and reverse: AAAGA ACAAT GGCCT TATAC AGG. The primers used to localize the coding region of the hCTL1 gene were forward: CATGT GGTGG TACCA TGTGG TGGG and reverse: CGAAT AAGGC GATTT ACTGA TGCC. The PCR product using the latter primers, CTL1-F3, was longer than predicted by the cDNA, 762 bases instead of 161, and was found to include an intron when sequenced.
Results
Isolation of a Torpedo Electric Lobe cDNA That Induces Choline Uptake in Mutant Yeast.
Complementation for choline-dependent growth under selective conditions by mutant yeast (ctr ise URA3Δ) transformed with a Torpedo electric lobe yeast expression library in pFL61 led to the isolation of an unique clone, named 4.17, associated with an increase in saturable [3H]choline uptake by mutant yeast beyond that observed with empty pFL61 (Fig. 1A). Sequencing of the cDNA corresponding to clone 4.17 indicated that it encodes a truncated transmembrane protein of 175 aa comprising three TMDs before the stop codon. A full-length cDNA coding for tCTL1 was cloned by hybridization with probes based on 4.17 from a λZAP electric lobe cDNA library and found to encode a protein with 10 putative TMDs (Fig. 2). When tCTL1 was expressed in mutant yeast, choline uptake was once again higher than for the empty plasmid (Fig. 1B). The analysis of the kinetic parameters for choline uptake showed that both yeast transformed with the empty plasmid and tCTL1 took up choline by high affinity mechanisms with Kms of ≈1 μM, but the maximal rate of choline uptake by yeast transformed with tCTL1 was 2.5 times greater than for yeast transformed with the plasmid itself. The tCTL1-transformed yeast were very sensitive to the competitive inhibitor HC-3 with a Ki of ≈0.5 μM (Fig. 1B). Yeast transformed with the control plasmid were similarly inhibited by 1 μM HC-3 (data not shown). The addition of 50 mM NaCl to the yeast medium, initially at 10 mM, had little effect on choline uptake by either group of yeast and had an inhibitory effect at 100 mM (data not shown). Choline uptake by mutant yeast expressing tCTL1 thus closely resembles the residual choline uptake in ctr yeast and partly resembles the neuronal choline transporter in cholinergic nerve terminals, known to have a high affinity for choline and to be inhibited by low concentrations of HC-3, but which is sodium dependent (1, 17). Only part of the tCTL1 proteins may be functional under these conditions because vertebrate transporters are not always readily sorted to the yeast plasma membrane (18).
Figure 1.

Suppression of a yeast choline transport mutation by heterologous expression of electric lobe cDNAs. (A) Pools of mutant yeast were transformed with individual plasmids isolated from colonies showing choline-dependent growth under selective conditions. Transformed yeast were incubated at 30°C for 30 min with 25 nM [3H]choline. Saturable choline uptake was measured as the difference in uptake in the presence and absence of 1 mM cold choline and normalized for growth (OD600); n.i. is a control plasmid without insert. Only a truncated clone, 4.17, had the capacity to induce choline uptake (means ± SEM for three to five independent transformations) above the low endogenous level. (B) The characterization of choline uptake by mutant yeast transformed with the full-length clone, tCTL1, corresponding to 4.17. Choline uptake by yeast transformed with tCTL1 was characterized by a Kt = 1.2 ± 0.6 μM and a Vmax = 211 ± 38 fmol/OD600 in 10 min. The apparent Kts in the presence of 0.5 and 1 μM HC-3 rose to 2.0 and 3.1 μM; respectively, the deduced Kis for HC-3 were 0.6 and 0.5 μM. Yeast transformed with a plasmid with no insert take up choline with a Kt = 1.7 ± 0.8 μM and Vmax = 82 ± 15 fmol/OD600 in 10 min. Results are presented as means ± SEM. from three independent transformations.
Figure 2.

Sequence alignments of CTL proteins and a model of their membrane topology. (A) Amino acid sequence alignment of CTL1 from Torpedo (tCTL1), rat (rCTL1), and human (hCTL1) with two homologous proteins, hCTL2 and hCTL4, and the single homologous C. elegans protein, F35C8.7. Black shading indicates 100% conservation and grey shading indicates 80% conservation (blosum 62). Stretches of hydrophobic amino acids that may form TMDs are indicated under the alignments and numbered. Potential N-glycosylation sites are underlined in each sequence. Conserved cysteines are marked with asterisks under the alignment. (B) Structural model of CTL1. (C) Dendrogram of tCTL1 and related proteins obtained by using clustalw. No prokaryote homologs have been found. The asterisks indicate that only partial protein sequences were used for the analysis.
Sequence Analysis and Alignment of the Members of the CTL Family.
A full-length cDNA coding for the rat ortholog (rCTL1, 69% identity with tCTL1) was cloned by hybridization from a rat brain cDNA library and sequenced. The protein sequences for the Torpedo and rat CTL1 genes as well as their human ortholog (hCTL1, 96% identity with the rat) are presented in Fig. 2A.
We further uncovered the first members of a family of several related proteins in humans and mice: hCTL4 (43% homology with hCTL1) and mCTL4, from conceptual translations of human and mouse genomic sequences (see Materials and Methods), and hCTL2 (43% homology with hCTL1 and 67% homology with hCTL4), for which we have completed the sequence corresponding to the coding region. All of these sequences are homologous to a single gene of this nature found in the genome of C. elegans, F35C8.7 (Fig. 2A). None of these proteins has either a known function or any apparent homology with other proteins. Neither domain analysis with blocks or prosite databases nor pattern searching are consensual, but transporters are frequently cited in alignments within TMDs 2 and 3. psi blast can be guided to list the majority of sodium-dependent transporters.
These proteins all show several transmembrane domains and can reasonably be thought to traverse the membrane 10 times (Fig. 2B). A first, large and variable loop between TMDs 1 and 2 is potentially extracellular and glycosylated. A highly conserved region covers the last four TMDs and includes the fourth extracellular loop that contains three of the 11 conserved cysteines and is potentially glycosylated only in the CTL1 orthologs. Interestingly, the conserved region includes the suppressor, 4.17, whose first ATG corresponds to M471 of tCTL1, suggesting that this region is sufficient to induce choline uptake. The proteins lack a clear signal peptide and are expected to be targeted to the plasma membrane.
Clustering relationships among these sequences and other partial sequences for another protein, CTL3, in mouse and human, as well as more distant homologs in other eukaryotes including conceptual translations from plant genomes are presented in Fig. 2C. The two plant proteins show only 26% homology with the hCTL1. Taken together, the information on the structure and sequence relationships indicates that this new transporter-like family has a widespread distribution among eukaryotes compatible with the nature of their proposed function as choline transporters.
Distribution of rCTL1 mRNA.
Northern blot analysis of rat mRNA distribution with an rCTL1 probe showed a particularly striking signal at 3.5 kb in the spinal cord and to a lesser extent in the brain, while a larger 5 kb form was prominent in the colon and notable in the lung and spinal cord (Fig. 3). A closer look at the tissues with strong expression by ISH showed that, indeed, several types of cells are expressing rCTL1 (Fig. 4). In the spinal cord, the highest expression was observed in large sized cells in the ventral horn, which were identified as motor neurons by double ISH experiments using, in addition, choline acetyltransferase cRNA antisense riboprobe (Fig. 4 E and F). Strong expression of rCTL1 transcripts was also observed in presumptive motor neurons present in the facial nucleus, suggesting an important role in these cholinergic neurons (data not shown). However, other neurons expressing choline acetyltransferase mRNA, such as those found in septal areas or the nucleus basalis, did not express high levels of rCTL1 mRNA (data not shown). Moreover, rCTL1 is not a specific marker for cholinergic neurons, even in the spinal cord, because we detected a high density of rCTL1 expressing cells scattered throughout both the gray and white matter (Fig. 4C). In the gray matter, the small, labeled cells might correspond to oligodendrocytes and interneurons. Indeed, in the brain, rCTL1 mRNA expression was particularly strong in myelinated fiber tracts. A high density of intensely labeled cells was observed in the cerebellar white matter, the brainstem, the corpus callosum, the fimbria of the hippocampus, and the lateral olfactory tract. In these areas, labeled cells appeared as short chains (Fig. 4B) as has been described for oligodendrocytes (19). Double ISH experiments using a riboprobe specific for myelin basic protein, a marker of oligodendrocytes (19), and rCTL1 performed at the level of the white matter of the cerebellum indicated that rCTL1 was indeed expressed in oligodendrocytes (data not shown). rCTL1 was also expressed at a lower level in neurons throughout the brain as exemplified in the hippocampal formation where rCTL1 transcripts were observed in pyramidal cells of the Ammon's horn and granule cells of the dentate gyrus (Fig. 4A), and also in dispersed cells throughout the hippocampus. Thus, rCTL1 appears to be expressed by both neurons and oligodendrocytes in the central nervous system.
Figure 3.

Northern blot analysis of rCTL1 mRNA in adult rat tissues. poly(A)+ RNA (2 μg) from the specified tissues was hybridized with a probe for rCTL1 and exposed for 7 hr. The lower panel shows hybridization with a β-actin probe.
Figure 4.
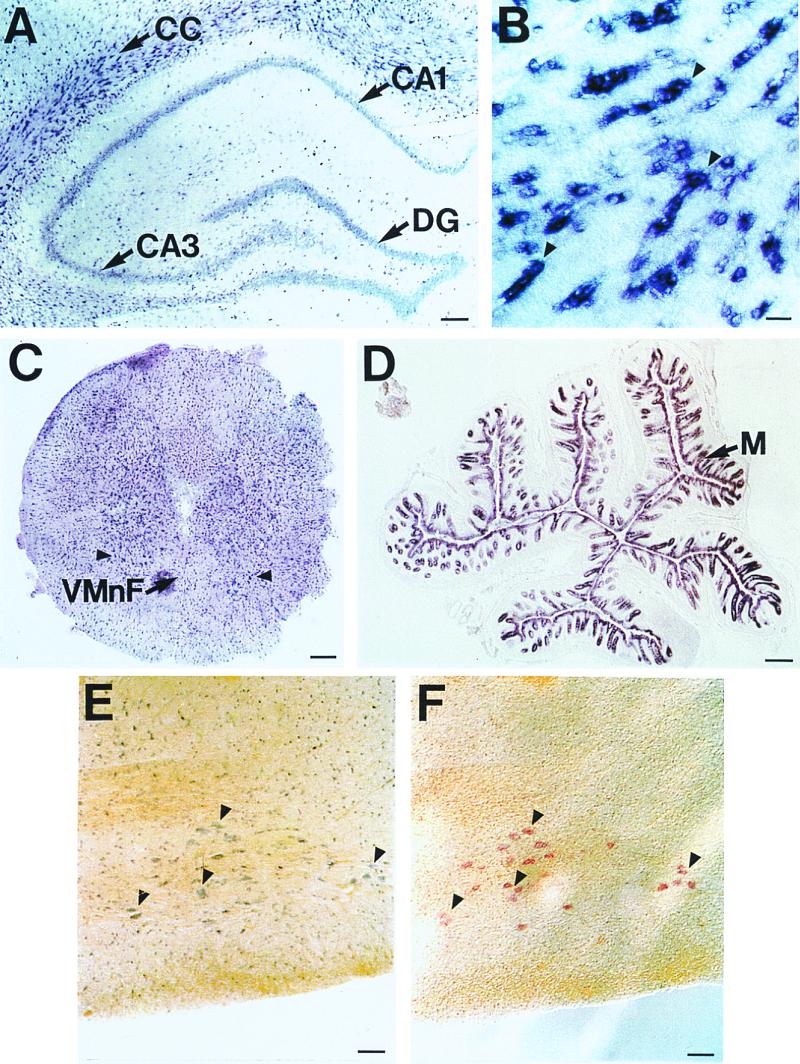
Distribution of rCTL1 in adult rat tissues by ISH. (A) Intense labeling with the antisense rCTL1 cRNA probe was observed within scattered cells present at a higher density in the corpus callosum (CC) than in cells located in the gray matter, as well as in the hippocampal neurons of the Ammon's horn (CA1–CA3) and of the dentate gyrus (DG). (B) A higher magnification of labeled cells within the CC showed chains of cells suggesting the labeling of oligodendrocytes (arrowheads). (C) A frontal section of cervical spinal cord showed a high density of small labeled cells in both white and gray matter, and larger labeled cells in the ventral horns (arrowheads); VMnF, ventral median fissure. (D) The cell layer constituting the colonic mucosa (M) expressed high levels of rCTL1 transcript. (E and F) Double ISH using the digoxigenin-labeled rCTL1 cRNA antisense probe (E) and a fluorescein-labeled choline acetyltransferase antisense riboprobe (F) identify the large cells expressing rCTL1 as motor neurons (arrowheads). Cryosections hybridized with digoxigenin-labeled and fluorescein-labeled sense probes showed no significant signal. Bars correspond to 200 μm (A, C, and D), 25 μm (B), and 100 μm (E and F).
rCTL1 expression also occurs outside of the central nervous system, especially in the colon where the larger transcript is prominent. In the colon, high levels of expression were observed in the mucosal cell layer both lining the crypts and exposed to the lumen (Fig. 4D).
Chromosomal Localization of hCTL1.
The analysis of human ESTs also indicated that sequences associated with hCTL1 included an sequence tag site, WI-17320, which has been mapped to 9q22/31. No YACs were known to carry this marker, so six YACs mapped to this locus were chosen for testing and two, 786_H_8 and 802_A_1, were found to be positive for WI-17320 by PCR analysis. Only these two YACs were also positive for a marker, CTL1-F3, defined for the 5′-coding region of hCTL1 (Fig. 5). Both YACs also carry the genetic marker D9S299, indicating a more precise localization of hCTL1 to 9q31.2. D9S299 is just proximal to the locus of mutation for familial dysautonomia (MIM 223900) (20).
Figure 5.

Colocalization of WI-17320 and hCTL1-F3 to YACs mapped to 9q31.2. Among the six YACs tested, only YACs 786_h_8 and 802_a_1 were found to be positive for the marker WI-17320 by PCR; the same YACs were also positive for a marker, CTL1-F3, for the 5′-coding region of hCTL1. Both of these YACs are also known to carry the genetic marker D9S299. Yeast actin is used as a positive control. The localization of the YACs on chromosome 9 (dotted lines leading to bars) is based on GenBank data.
Information on the chromosomal localization of hCTL4 and hCTL2 is also available; human NG22 (hCTL4) was sequenced as part of the major histocompatibility complex III at 6p21.3, and the assembled hCTL2 sequence includes two sequence tag sites independently mapped to 19p13.1. Hence, CTL resembles many other human genes with duplication between chromosomes 1, 6, 9, and 19 (21–23). More information on normal patterns of expression for these genes may be informative for potential links with genetic disorders.
Discussion
While looking for the neuronal choline transporter, we have isolated a Torpedo electric lobe clone, 4.17, capable of suppressing a choline transport mutation in yeast. High levels of expression of the rat homolog, rCTL1, are found in motor neurons and oligodendrocytes, but lower levels of expression also occurred in neuronal populations throughout the brain. The distribution of rCTL1 in the central nervous system, thus, clearly differs from that expected for a presynaptic cholinergic protein. In addition, a larger 5-kb rCTL1 transcript is associated with strong expression in the mucosal cell layer of the colon; given the stringency of the hybridation conditions, this mRNA probably represents a splice variant of rCTL1. CTL1 may provide choline for phospholipid synthesis rather than for acetylcholine synthesis. Nonetheless, cholinergic neurons may be particularly sensitive to any changes in choline metabolism because they need choline for both membrane and neurotransmitter synthesis (24, 25).
In ctr mutant yeast, the expression of tCTL1 is associated with an increase in sodium independent high affinity choline uptake, similar in nature to the choline uptake that remains in the mutant yeast. This observation is perhaps not so surprising since the yeast also have a gene, YOR161c, that is homologous to CTL1 and, hence, could act as a backup under these conditions.
The description of the CTL1 orthologs has further led to the description of a novel family of membrane proteins, which we call the CTL family. This family is characterized by 10 putative TMDs and 11 highly conserved cysteines. These proteins share limited structural homology with transporters, especially in TMDs 2 and 3, but none of the CTL family proteins has a known function. Plants and simple animals, such as C. elegans, have only one gene of this nature. Humans and mice, on the other hand, clearly have three, and probably four, different homologous genes located at chromosomal positions already known for grouping of duplicated genes.
Cloning of this novel family of proteins opens the way to the development of pharmacological tools that might be useful for the treatment of diseases linked to perturbations of CTL functions in humans. The chromosomal localization of the hCTL1 in the vicinity of D9S299 at 9q31.2 places this gene in close proximity to the locus responsible for the genetic disorder familial dysautonomia. The strong expression of rCTL1 in motor neurons and oligodendrocytes, as well as the functional link to choline uptake make it an appealing candidate for the site of mutation causing familial dysautonomia, a disease that includes a peripheral cholinergic component (26) with both autonomic and motor manifestations at birth, and progressive demyelination in the central nervous system of adults (27). However, our present efforts to refine the genomic localization of hCTL1 combined with the recent narrowing in the range of markers neighboring with the locus associated with familial dysautonomia (20) indicate that the exonic region of hCTL1 is close, but proximal, to the site of mutation, leaving the possibility of a perturbation in the 5′ regulatory domain. Further genetic and functional studies should delineate the implication of hCTL1 in this or other disease states.
Acknowledgments
We thank Dr. M. Israël, in whose laboratory this work was done, for many helpful discussions. We thank Dr. F. Lacroute for his help in providing both the plasmid pFL61 and advice on heterologous complementation. The original yeast strain D308–14D was kindly provided by Dr. S. Yamashita. We are grateful to the Association Française contre les Myopathies for their financial support. M.R. is supported by a Junior group ATIPE and by the Association pour la Recherche sur le Cancer.
Abbreviations
- TMDs
transmembrane domains
- ISH
in situ hybridization
- HC-3
hemicholinium-3
- EST
expressed sequence tag
- YACs
yeast artificial chromosomes
Footnotes
This paper was submitted directly (Track II) to the PNAS office.
Data deposition: The sequences reported in this paper have been deposited in the GenBank database [accession nos. AJ245618 (tCTL1), AJ245619 (rCTL1), AJ245620 (hCTL1), and AJ245621 (hCTL2)].
Article published online before print: Proc. Natl. Acad. Sci. USA, 10.1073/pnas.030339697.
Article and publication date are at www.pnas.org/cgi/doi/10.1073/pnas.030339697
References
- 1.Yamamura H I, Snyder S H. J Neurochem. 1973;21:1355–1374. doi: 10.1111/j.1471-4159.1973.tb06022.x. [DOI] [PubMed] [Google Scholar]
- 2.Kuhar M J, Murrin L C. J Neurochem. 1978;30:15–21. doi: 10.1111/j.1471-4159.1978.tb07029.x. [DOI] [PubMed] [Google Scholar]
- 3.Jope R S. Brain Res. 1979;180:313–344. doi: 10.1016/0165-0173(79)90009-2. [DOI] [PubMed] [Google Scholar]
- 4.Knipper M, Kahle C, Breer H. Biochim Biophys Acta. 1991;1065:107–113. doi: 10.1016/0005-2736(91)90219-x. [DOI] [PubMed] [Google Scholar]
- 5.Rylett R J, Walters S A, Davis W. Brain Res Mol Brain Res. 1996;35:354–358. doi: 10.1016/0169-328x(95)00265-t. [DOI] [PubMed] [Google Scholar]
- 6.Nikawa J, Tsukagoshi Y, Yamashita S. J Bacteriol. 1986;166:328–330. doi: 10.1128/jb.166.1.328-330.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Sentenac H, Bonneaud N, Minet M, Lacroute F, Salmon J M, Gaymard F, Grignon C. Science. 1992;256:663–665. doi: 10.1126/science.1585180. [DOI] [PubMed] [Google Scholar]
- 8.Nikawa J, Hosaka K, Tsukagoshi Y, Yamashita S. J Biol Chem. 1990;265:15996–16003. [PubMed] [Google Scholar]
- 9.Minet M, Dufour M E, Lacroute F. Plant J. 1992;2:417–422. doi: 10.1111/j.1365-313x.1992.00417.x. [DOI] [PubMed] [Google Scholar]
- 10.Gietz R D, Schiestl R H, Willems A R, Woods R A. Yeast. 1995;11:355–360. doi: 10.1002/yea.320110408. [DOI] [PubMed] [Google Scholar]
- 11.Hosaka K, Yamashita S. J Bacteriol. 1980;143:176–181. doi: 10.1128/jb.143.1.176-181.1980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.O'Regan S, Diebler M-F, Meunier F-M, Vyas S. J Neurochem. 1995;64:69–76. doi: 10.1046/j.1471-4159.1995.64010069.x. [DOI] [PubMed] [Google Scholar]
- 13.Schaeren-Wiemers N, Gerfin-Moser A. Histochemistry. 1993;100:431–440. doi: 10.1007/BF00267823. [DOI] [PubMed] [Google Scholar]
- 14.Traiffort E, Charytoniuk D A, Watroba L, Faure H, Sales N, Ruat M. Eur J Neurosci. 1999;11:3199–3214. doi: 10.1046/j.1460-9568.1999.00777.x. [DOI] [PubMed] [Google Scholar]
- 15.Arce V, Pollock R A, Philippe J M, Pennica D, Henderson C E, deLapeyriere O. J Neurosci. 1998;18:1440–1448. doi: 10.1523/JNEUROSCI.18-04-01440.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Brice A, Berrard S, Raynaud B, Ansieau S, Coppola T, Weber M J, Mallet J. J Neurosci Res. 1989;23:266–273. doi: 10.1002/jnr.490230304. [DOI] [PubMed] [Google Scholar]
- 17.Haga T, Noda H. Biochim Biophys Acta. 1973;291:564–575. doi: 10.1016/0005-2736(73)90508-7. [DOI] [PubMed] [Google Scholar]
- 18.Hogue D L, Ellison M J, Young J D, Cass C E. J Biol Chem. 1996;271:9801–9808. doi: 10.1074/jbc.271.16.9801. [DOI] [PubMed] [Google Scholar]
- 19.Shiota C, Miura M, Mikoshiba K. Brain Res Dev Brain Res. 1989;45:83–94. doi: 10.1016/0165-3806(89)90010-2. [DOI] [PubMed] [Google Scholar]
- 20.Blumenfeld A, Slaugenhaupt S A, Liebert C B, Temper V, Maayan C, Gill S, Lucente D E, Idelson M, MacCormack K, Monahan M A, et al. Am J Hum Genet. 1999;64:1110–1118. doi: 10.1086/302339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Katsanis N, Fitzgibbon J, Fisher E C. Genomics. 1996;35:101–108. doi: 10.1006/geno.1996.0328. [DOI] [PubMed] [Google Scholar]
- 22.Sugaya K, Sasanuma S, Nohata J, Kimura T, Fukagawa T, Nakamura Y, Ando A, Inoko H, Ikemura T, Mita K. Gene. 1997;189:235–244. doi: 10.1016/s0378-1119(96)00857-8. [DOI] [PubMed] [Google Scholar]
- 23.Hughes A L. Mol Biol Evol. 1998;15:854–870. doi: 10.1093/oxfordjournals.molbev.a025990. [DOI] [PubMed] [Google Scholar]
- 24.Blusztajn J K, Wurtman R J. Science. 1983;221:614–620. doi: 10.1126/science.6867732. [DOI] [PubMed] [Google Scholar]
- 25.Wurtman R J. Trends Neurosci. 1992;15:117–122. doi: 10.1016/0166-2236(92)90351-8. [DOI] [PubMed] [Google Scholar]
- 26.Mittag T W, Mindel J S, Green J P. Ann NY Acad Sci. 1974;228:301–306. doi: 10.1111/j.1749-6632.1974.tb20518.x. [DOI] [PubMed] [Google Scholar]
- 27.Pearson J, Pytel B A. J Neurol Sci. 1978;39:47–59. doi: 10.1016/0022-510x(78)90187-9. [DOI] [PubMed] [Google Scholar]