

Abstract
We have demonstrated that amino acids E323, Y324, E330, and V331 from the factor Va heavy chain are required for the interaction of the cofactor with factor Xa and optimum rates of prothrombin cleavage. We have also shown that amino acid region 332−336 contains residues that are important for cofactor function. Using overlapping peptides, we identified amino acids D334 and Y335 as contributors to cofactor activity. We constructed recombinant factor V molecules with the mutations D334 → K and Y335 → F (factor VKF) and D334 → A and Y335 → A (factor VAA). Kinetic studies showed that while factor VaKF and factor VaAA had a KD for factor Xa similar to the KD observed for wild-type factor Va (factor VaWT), the clotting activities of the mutant molecules were impaired and the kcat of prothrombinase assembled with factor VaKF and factor VaAA was reduced. The second-order rate constant of prothrombinase assembled with factor VaKF or factor VaAA for prothrombin activation was ∼10-fold lower than the second-order rate constant for the same reaction catalyzed by prothrombinase assembled with factor VaWT. We also created quadruple mutants combining mutations in the amino acid region 334–335 with mutations at the previously identified amino acids that are important for factor Xa binding (i.e., E323Y324 and E330V331). Prothrombinase assembled with the quadruple mutant molecules displayed a second-order rate constant up to 400-fold lower than the values obtained with prothrombinase assembled with factor VaWT. The data demonstrate that amino acid region 334–335 is required for the rearrangement of enzyme and substrate necessary for efficient catalysis of prothrombin by prothrombinase.
The mainstay of the hemostasis process is the generation of thrombin, which in turn promotes the assembly of the fibrin plug following vascular injury. The prothrombinase complex is composed of the enzyme, factor Xa, and the protein cofactor, factor Va, associated in the presence of divalent metal ions on a membrane surface (1–3). This complex catalyzes the activation of prothrombin to its active form, thrombin. Factor Xa alone can activate prothrombin by two sequential proteolytic cleavages at Arg271 and Arg320 resulting in the intermediates fragment 1·2 and prethrombin 2 (4–14). The overall rate of this reaction is not compatible with survival; however, incorporation of the cofactor, factor Va, into prothrombinase reverses the order of the proteolytic cleavages and increases the catalytic activity of factor Xa by 5 orders of magnitude (7,8,15). The first, factor Va-dependent cleavage at Arg320 produces the intermediate meizothrombin. Further cleavage at Arg271 generates thrombin and fragment 1·2 (4–14). The regulation of this key step in the blood coagulation cascade has been under intense investigation for many years.
Plasma factor V circulates as an inactive protein cofactor with an Mr of 330000 that does not participate in the prothrombinase complex (16). Factor V must first be activated to factor Va by thrombin through three sequential proteolytic cleavages at Arg709, Arg1018, and Arg1545 to produce a heavy chain, composed of amino acid residues 1−709 (Mr = 105000) and a light chain composed of amino acid residues 1546−2196 (Mr = 74000) (17) (Figure 1). The crystal structure of the bovine factor Va molecule lacking the entire A2 domain and the acidic COOH terminus of the heavy chain (factor Vai) has been determined (18). Using this crystal structure as a template, we have provided a complete model of the human cofactor in solution (19). This model allows for the identification of the structural perturbations of point mutations on the overall tertiary cofactor structure.
Figure 1.

Proteolytic processing of human factor V. Factor V is composed of 2196 amino acids and has three A domains (A1, A2, and A3), a connecting B region, and two C domains (C1 and C2). The procofactor is activated following three cleavages by thrombin at Arg709, Arg1018, and Arg1545. The latter cleavage is the last to occur and is a prerequisite for light chain formation and maximum cofactor activity generation. These cleavages release the active cofactor composed of heavy and light chains associated in the presence of divalent metal ions, and two activation fragments. The mutations within a 45-amino acid region of the A2 domain of the heavy chain of the molecule that is involved in cofactor activity (hatched box) are indicated together with the designation for the recombinant mutant factor V molecules created and used throughout the paper. The black box preceding cleavage at Arg709 depicts the acidic COOH-terminal region of factor Va that is essential for optimum cofactor activity.
Both chains of factor Va have been shown to be required for the interaction of the cofactor with factor Xa (20,21). While the binding sites on the light chain remain to be identified, several binding sites for factor Xa on the factor Va heavy chain have been delineated (22–24). Analysis of the inactivation fragments of factor Va by plasmin and activated protein C (APC) demonstrated that amino acid region 307−348 is critical for factor Va cofactor function. A synthetic peptide spanning this amino acid region was found to be a good inhibitor of factor V/Va clotting activity [N42R (23)]. This region of factor Va is thought to be important for cofactor function since it is 82% conserved among human (25), bovine (26), mouse (27), and porcine (28) species. Using overlapping synthetic peptides from this region and recombinant factor Va molecules, we have demonstrated that amino acid region 323−331 of the factor Va heavy chain contains critical amino acid residues responsible for the direct interaction of the cofactor with factor Xa (23,29). In addition, our initial data also demonstrated that amino acid sequence 332IWDYA336, which is continuous to the factor Xa binding site, was also involved in the inhibition of prothrombinase activity (23). This study was undertaken to improve our understanding of the precise contribution of amino acid region 332−336 of the factor Va heavy chain to the catalytic efficiency of prothrombinase.
Experimental Procedures
Materials, Reagents, and Proteins
Diisopropyl fluorophosphate (DFP), O-phenylenediamine (OPD) dihydrochloride, N-(2-hydroxyethyl)piperazine-N′-2-ethanesulfonic acid (Hepes), Trizma (Tris base), and Coomassie Blue R-250 were purchased from Sigma (St. Louis, MO). Secondary anti-mouse and anti-sheep IgG coupled to peroxidase were purchased from Southern Biotechnology Associates Inc. (Birmingham, AL). l-α-Phosphatidylserine (PS)1 and l-α-phosphatidylcholine (PC) were from Avanti Polar Lipids (Alabaster, AL). Chemiluminescent reagent ECL+ and heparin-Sepharose were from AmershamPharmacia Biotech Inc. (Piscataway, NJ). Normal reference plasma and the chromogenic substrates Spectrozyme-TH and Spectrozyme-Xa were from American Diagnostica Inc. (Greenwich, CT). RecombiPlasTin was purchased from Instrumentation Laboratory (Lexington, MA). Human factor Xa was from Enzyme Research Laboratories (South Bend, IN). Factor V-deficient plasma was from Research Proteins Inc. (Essex Junction, VT). Dansylarginine-N-(3-ethyl-1,5-pentanediyl) amide (DAPA), RVV-factor V activator, human thrombin, and human prothrombin were purchased from Haematologic Technologies Inc. (Essex Junction, VT). Factor V cDNA (ATCC catalog no. 40515 pMT2-V) was from American Type Culture Collection (Manassas, VA). All restriction enzymes were from New England Biolabs (Beverly, MA). All molecular biology and tissue culture reagents and media were from Gibco, Invitrogen Corp. (Grand Island, NY), or as indicated. Monoclonal antibodies αHFVHC17 and αHFVLC9 and monoclonal antibody αHFV1 coupled to Sepharose were provided by K. G. Mann (Department of Biochemistry, University of Vermont, Burlington, VT).
Thrombin Generation Assay Using a Fluorescent Thrombin Inhibitor
Thrombin generation assays were performed using the fluorescent thrombin inhibitor DAPA as described previously (23). The buffer used was composed of 20 mM Tris, 0.15 M NaCl (pH 7.4), and 5 mM CaCl2 (TBS, Ca2+) and was made fresh and filtered before each use. Fluorescence was measured on a Perkin-Elmer LS-50B luminescence spectrometer (Perkin-Elmer Corp., Norwalk, CT) with a λex of 280 nm, a λem of 550 nm, and a 500 nm long-pass filter in the emission beam (Schott KV-500). The data points were captured using FL WinLab (Perkin-Elmer Corp.) and subsequently analyzed and plotted using Prizm (GraphPad, San Diego, CA). Peptides AP3 and AP4′ were made in the analytical facility of A. Kurosky (University of Texas, Medical Branch, Galveston, TX). Peptides IWDYA, AP5, AP4′, AP5m(DY→KF), AP6, and the control P15H were purchased from New England Peptide (NEP, Gardner, MA). Peptides were dissolved in water or buffer. Accurate peptide concentrations were determined by amino acid composition in the analytical facility of A. Kurosky as detailed previously (30).
Fluorescence Anisotropy Measurements
The fluorescence anisotropy of [OG488]-EGR-hXa was measured using a Perkin-Elmer LS-50B luminescence spectrometer in L-format essentially as extensively described previously (23). Anisotropy measurements were performed in a quartz cuvette under constant stirring (low) with a λex of 490 nm, a λem of 520 nm, and a long-pass filter (Schott KV-520) in the emission beam. All experiments were performed in the dark. The data obtained were instantly analyzed using FL WinLab (Perkin-Elmer Corp.).
PCR Site-Directed Mutagenesis and Transient Expression of Factor V Molecules
Factor VKF was constructed using the mutagenic primers 5′-C ATT TGG AAG TTT GCA CCT G-3′ (forward) and 5′-C AGG TGC AAA CTT CCA AAT G-3′ (reverse) (bold underlined letters identify the mutated bases) in a two-stage PCR method and characterized as previously described (29). The resulting amplicon was subcloned into pGEM-T, and the mutations were confirmed by DNA sequencing (DNA Analysis Facility, Cleveland State University). The pGEM-T plasmid was digested with restriction enzymes Bsu36I and XcmI to remove the factor V insert, and the insert containing the mutation was ligated into pMT2-FV at the same restriction sites. The remaining recombinant plasmids were constructed using Stratagene’s QuikChange XL site-directed mutagenesis kit according to the manufacturer’s instructions. Factor VAA was constructed with the primers 5′-GAG GAA GTC ATT TGG GCC GCC GCA CCT GTA ATA-3′ (forward) and 5′-TAT TAC AGG TGC GGC GGC CCA AAT GAC TTC CTC-3′ (reverse). Factor VMI/KF was constructed with the primers 5′-GAA TAC TTC ATT GCT GCA GAG ATGGTC ATT TGG AAG TTT GCA CCT GTA ATA-3′ (forward) and 5′-TAT TAC AGG TGC AAA CTT CCA AAT GACCAT CTC TGC AGC AAT GAA GTA TTC-3′ (reverse). Factor VFF/KF was constructed with the primers 5′-CAC ATG AAG ATT TGG TTT TTC TTC ATT GCT GCA GAG GAA GTC ATT TGG AAG TTT GCA CCT GTA ATA-3′ (forward) and 5′-TAT TAC AGG TGC AAA CTT CCA AAT GAC TTC CTC TGC AGC AAT GAA GAA AAA CCA AAT CTT CAT GTG-3′ (reverse). Factor VMI/AA was constructed with the primers 5′-GAA TAC TTC ATT GCT GCA GAG ATGGTC ATT TGG GCC GCC GCA CCT GTA ATA-3′ (forward) and 5′-TAT TAC AGG TGC GGC GGC CCA AAT GACCAT CTC TGC AGC AAT GAA GTA TTC-3′ (reverse). Factor VFF/AA was constructed with the primers 5′-CAC ATG AAG ATT TGG TTT TTC TTC ATT GCT GCA GAG GAA GTC ATT TGG GCC GCC GCA CCT GTA ATA-3′ (forward) and 5′-TAT TAC AGG TGC AAA CTT CCA AAT GAC TTC CTC TGC AGC AAT GAA GACCAT CCA AAT CTT CAT GTG-3′ (reverse). Factor VI332A was constructed with the primers 5′-GCT GCA GAG GAA GTC GCT TGG GAC TAT GCA CCT GTA ATA-3′ (forward) and 5′-TAT TAC AGG TGC ATA GTC CCA AGC GAC TTC CTC TGC AGC-3′ (reverse). Factor VW333A was constructed with the primers 5′-GCT GCA GAG GAA GTC ATT GCG GAC TAT GCA CCT GTA ATA-3′ (forward) and 5′-TAT TAC AGG TGC ATA GTC CGC AAT GAC TTC CTC TGC AGC-3′ (reverse). In all the primers, the bases that are bold and underlined represent the mismatch. The mutations were confirmed by DNA sequencing (DNA Analysis Facility, Cleveland State University).
Transfection of Recombinant Factor V Molecules into COS-7L Cells
The COS-7L cell line was maintained in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% fetal bovine serum and the antibiotics streptomycin (100 µg/mL) and penicillin (100 IU/mL) in an atmosphere of 5% CO2 and 95% air at 37 °C. The purified wild-type and mutated plasmids were transfected into the COS-7L cells with Fugene 6 (Roche Diagnostics) as described previously (29,31).
Purification of Recombinant Proteins
The medium containing the recombinant proteins was centrifuged to remove any cellular debris. Recombinant factor VKF, factor VAA, factor VI332A, factor VW333A, and factor VWT were purified and quantified as recently described in detail (32). In some instances, the quadruple recombinant factor V mutants, factor VKF, as well as factor VWT were partially purified and quantified as extensively detailed previously (29,31). The activities of the partially purified recombinant factor Va molecules (29,31) and the activities of the recombinant cofactor molecules purified to homogeneity (32) were similar and are reported in Table 1. The proteins were stored at −80 °C in small aliquots to prevent repeated freeze−thaw cycles. The activity and integrity of the recombinant factor V molecules were confirmed by clotting assays using factor V-deficient plasma and Western blotting with monoclonal and polyclonal antibodies.
Table 1. Functional Properties of Recombinant Factor Va Molecules.
| factor Va species | clotting activity (units/mg)a | KDapp (nM)b | Km (µM)c |
|---|---|---|---|
| factor VaPLASMA | 2380 ± 150 | 0.35 ± 0.07 | 0.16 ± 0.1 |
| factor VaWT | 2390 ± 240 | 0.4 ± 0.1 | 0.13 ± 0.06 |
| factor VaWTd | − | 0.24 ± 0.12 | 0.16 ± 0.1 |
| factor VaKF | 440 ± 74 | 0.58 ± 0.2 | 0.42 ± 0.06 |
| factor VaKFd | − | 0.55 ± 0.13 | 0.6 ± 0.01 |
| factor VaAA | 550 ± 70 | 0.6 ± 0.15 | 0.62 ± 0.07 |
| factor VaFFde | − | 0.7 ± 0.3 | 0.10 ± 0.01 |
| factor VaMIde | − | 1.8 ± 0.7 | 0.14 ± 0.01 |
| factor VaFF/KFd | ∼30 | 1.3 ± 0.4 | 0.66 ± 0.2 |
| factor VaFF/AAd | ∼46 | 2.5 ± 0.8 | 0.6 ± 0.3 |
| factor VaMI/KFd | ≤30 | 5.5 ± 1 | 0.6 ± 0.35 |
| factor VaMI/AAd | ≤30 | 4.6 ± 0.8 | 0.4 ± 0.25 |
| factor VaI332A | 2103 ± 360 | 0.33 ± 0.06 | 0.39 ± 0.07 |
| factor VaW333A | 2150 ± 185 | 0.37 ± 0.08 | 0.35 ± 0.05 |
Two-stage clotting assays of recombinant factor V molecules was performed as described in .
Apparent dissociation constants of recombinant factor Va for plasma-derived factor Xa (KDapp) were determined as described in at limiting factor Xa concentrations (15 pM).
Apparent Km and kcat (shown in Figure 5B) values were determined as described in at limiting factor Xa concentrations (5 pM) and saturating concentrations of factor Va.
The recombinant molecules were partially purified and assayed as described in detail in ref (29).
The recombinant molecules were fully characterized in ref (31).
Prothrombin Activation Analysis by Gel Electrophoresis
Prothrombin (1.4 µM) was incubated in a reaction mixture containing 20 µM PCPS, 50 µM DAPA, and various recombinant factor Va molecules at 10 nM (activated with thrombin at a 1/50 enzyme/substrate ratio) in TBS, Ca2+. The addition of factor Xa (1 nM) marked the start of the reaction. Aliquots of the reaction mixture were removed at selected time points and added to 2 volumes of 0.2 M glacial acetic acid and treated as described previously (31). The dried samples were reconstituted in 0.1 M Tris base (pH 6.8), 1% SDS, and 1% β-mercaptoethanol and heated for 75 s at 95 °C. A total of 5 µg of total protein was loaded per lane and analyzed via 9.5% SDS−PAGE.
Gel Electrophoresis and Western Blotting
Sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS−PAGE) analyses were carried out using 9.5% or 4 to 12% gradient gels following reduction with 2% β-mercaptoethanol as described previously (33). In some cases, the protein was transferred to polyvinylidene difluoride (PVDF) membranes as described previously (34). Factor Va heavy and light chains were probed with the appropriate monoclonal antibodies and visualized with chemiluminescence, or the protein was visualized by staining with Coomassie Brilliant Blue R-250, followed by destaining in a solution of methanol, acetic acid, and water. In some experiments, gels were analyzed by scanning densitometry using UN-SCAN-IT gel (Silk Scientific, Orem, UT) as described previously (30,31).
Assay Measuring Thrombin Formation
The ability of the recombinant factor V molecules to assemble in the prothrombinase complex was assessed in a discontinuous assay described in detail elsewhere (31). In short, all recombinant factor V molecules were activated with thrombin (1/50 enzyme/substrate ratio, 10 min at 37 °C) as described recently (35). Reaction mixtures contained PCPS vesicles (20 µM), DAPA (3 µM), factor Xa (varying concentrations), and recombinant factor Va species in reaction buffer [HEPES, 0.15 M NaCl, 50 nM CaCl2, and 0.01% Tween 20 (pH 7.40)]. DAPA (at twice the concentration of prothrombin) was included in all mixtures to prevent the action of thrombin during the course of the assay on factor Va, on prothrombin, and on itself. The rate of thrombin generation was subsequently measured using an excess of chromogenic substrate, Spectrozyme-TH (0.4 mM). The initial rate of thrombin generation was analyzed with Prizm (GraphPad), and kinetic constants were extracted directly from the graphs.
For the calculation of the KD between the factor Va molecules and factor Xa, assays were performed in the presence of a limiting factor Xa concentration (15 pM) and varying concentrations of the recombinant factor Va species (from 25 pM to 5 nM). For the determination of the kinetic constants of prothrombinase assembly, Km and kcat, experiments were executed with a limiting amount of factor Xa (5 pM) in the presence of a fixed amount of the various recombinant factor Va molecules (10 nM) and a varied amount of substrate (prothrombin). Experiments performed with the quadruple mutants used saturating concentrations of factor VFF/KF, factor VMI/KF, factor VFF/AA, and factor VMI/AA and varying amounts of the substrate prothrombin (from 25 nM to 4 µM). The concentration of each mutant cofactor molecule necessary to saturate factor Xa was calculated with the quadratic equation (eq 1) described in the literature (36,37) and given below.
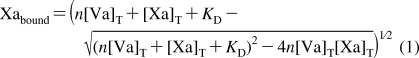 |
where [Va]T and [Xa]T are the total concentrations of factor Va and factor Xa, respectively. The apparent dissociation constant (KD) for the bimolecular interaction between factor Va and factor Xa on a phospholipid surface was obtained from the functional titration described above and the total concentration of ligand ([Va]T) modified as appropriate.
Additive Effect of the Mutations on Prothrombinase Function
The change in transition-state stabilization free energy, which measures the effect that mutations in the cofactor of the prothrombinase complex have on the catalytic site of the enzyme, was calculated for the quadruple mutants as extensively described previously (38–42), and recently (31). In brief, the perturbation to the function of prothrombinase assembled with wild-type factor Va (state A) caused by a mutation in factor Va (state B) affecting the transition state can be defined in general as follows:
Since prothrombinase activity (assembled with each of the recombinant factor Va proteins) is being measured against the same substrate (prothrombin), the transition-state stabilization free energy (ΔΔG⧧A→B) during catalysis induced by a mutation in factor Va can be determined from the following equation:
where R is the universal gas constant (2 cal K−1 mol−1), T is the absolute temperature (298 K in the experiments presented here), kcat is the turnover number, and Km is the Michaelis−Menten constant of the reaction. Once calculated, these values can be used to determine ΔΔGint, which reflects the exchange in free energy between the amino acid side chains manipulated in this study (E323F, Y324F, E330M, V331I, D334K, Y335F, D334A, and Y335A) by the following equation:
A positive value of ΔΔGint indicates that the interaction of these amino acid side chains reduces the catalytic efficiency of prothrombinase, while a negative value demonstrates that the mutations are better for prothrombinase, resulting in an increase in the catalytic efficiency of the enzyme. A value of zero would indicate no effect.
Molecular Dynamics Simulation Setup
Factor VaKF and factor VaAA were modeled using the recent factor Va model (PDB entry 1y61) (19). Briefly, amino acid residues Asp334 and Tyr335 were substituted with both lysine and phenylalanine, respectively, or with alanine. The amino acid change was performed using the “mutate” tool from the SwissPDBViewer package (43). Wild-type factor Va was left unchanged. The molecular dynamics simulation of each factor Va model consisted of a three-step setup: (1) the solvation of the protein followed by energy minimization, (2) position restraint molecular dynamics simulation, and (3) production run molecular dynamics simulation. Simulations were run and analyzed using GROMACS version 3.2.1 (44,45). The system was solvated using the SPC water model (46). The force used to describe the interaction parameters was set to GROMOS-87 implemented in GROMACS as “ffgmx” (47). Molecular topologies for both wild-type factor Va and factor VaKF were generated using the atom types described in the “ffgmx” force field. Bonds were constrained using the LINCS algorithm (48). Long-range electrostatic interactions were evaluated with the particle mesh Ewald summation method performed in all three dimensions and using a fourth-order Fourier interpolation on a 1.6 Å spaced grid (49,50). Following the minimization procedure, the systems were subjected to a 20 ps molecular dynamics simulation with the restriction that only water molecules were allowed to move freely and the protein was restrained. The simulation temperature was set to 300 K and was maintained using the Berendsen temperature coupling with the coupling parameter set to 0.1 ps (51). The constant reference pressure of 1 bar was maintained throughout the simulation using the Parrinello−Rahman isotropic pressure coupling scheme with the compressibility set to 4.5 × 10−5 bar−1(52). Simulations were run until the systems reached equilibrium. Equilibration of each system was confirmed when the root-mean-square displacement of the protein reached a plateau and the drift was less than 1 Å during a whole nanosecond simulation time. The last nanosecond of the equilibrated trajectories was analyzed using tools included in GROMACS (44,45).
Results
Inhibition of Prothrombinase Function by Synthetic Peptides from the Central Portion of the Factor Va Heavy Chain
We have previously shown that a peptide representing amino acid residues 323−331 inhibits factor Va cofactor activity (23). We have extensively characterized the important amino acids from this region by site-directed mutagenesis (29). We have also shown that under our experimental conditions, peptides encompassing residues 327−336 (AP5) and 332−341 (AP6) of the factor Va heavy chain also inhibit factor Va cofactor activity, while the pentadecapeptide P15H (representing amino acids 337−351 of factor V) did not have any effect on factor Va activity and was a good negative control. We have thus hypothesized that the overlapping sequence between AP5 and AP6 (amino acid motif 332IWDYA336) may contain amino acids responsible for the inhibition of prothrombinase. To investigate the role of these amino acids as it pertains to prothrombinase assembly and function, we have initiated a series of experiments using peptides from this region (Figure 2). The peptides were assayed for their ability to inhibit prothrombinase activity in an assay that uses purified reagents and a fluorescent thrombin inhibitor (Figure 3A). The data demonstrate that AP5 inhibits prothrombinase with an IC50 of ∼11 µM [Figure 3A (◆)], while the control peptide (P15H) had no effect on prothrombinase activity under the conditions employed [Figure 3A (◼)], even at concentrations as high as 500 µM (not shown). Complete inhibition of prothrombinase by AP5 occurred at 200 µM peptide (not shown). The Ki for the inhibition of prothrombinase by AP5 calculated from the IC50 was found to be 5.9 µM. This constant represents the dissociation constant of AP5 for prothrombinase. We have also assessed the effect of AP5 and AP6 for their ability to inhibit the direct binding of factor Va to [OG488]-EGR-hXa. Both peptides at concentrations as high as 100 µM were unable to inhibit the direct interaction of factor Va with [OG488]-EGR-hXa [Figure 3B (◆ and ●, respectively)]. In contrast, the nonapeptide control [Figure 3B, AP4′ (▼)] was a good inhibitor of the interaction of factor Va with [OG488]-EGR-hXa as previously described (23). In addition, AP3 that contains the NH2-terminal portion of AP4′ also inhibited the direct interaction of factor Va with [OG488]-EGR-hXa [Figure 3B (▲)], albeit less efficiently than AP4′. It is noteworthy that AP3 is a noncompetitive inhibitor of prothrombinase (Figure S1) with respect to substrate, like AP4′ (23). Kinetic analyses demonstrated that in the presence of increasing concentrations of inhibitor, the Km remained constant (0.4 ± 0.06) while the Vmax of the enzymatic reaction decreased (Figure S1A). Using the mathematical transformation associated with the Dixon plots, the apparent Ki of prothrombinase with AP3 was found to be ∼15.6 µM (Figure S1B). Finally, it is important to underline that control experiments demonstrated that AP5 did not have any effect on the catalytic activity of factor Xa, since high concentrations of peptide were unable to inhibit factor Xa activity toward both a chromogenic substrate and prothrombin (data not shown).
Figure 2.

Factor Va heavy chain peptides. At the top, the sequence of N42R is illustrated (part of the hatched box in Figure 1). The underlined sequence (323−335, 13 amino acids) is the regulatory amino acid sequence of the factor Va heavy chain. Overlapping peptides from the central portion of the factor Va heavy chain [AP3−AP7 (23)] are also shown. The role of the underlined amino acid region included in the box (i.e., Arg334-Tyr335) and common to IWDYA, AP5 and AP6, is under investigation here. Bold and underlined residues are the amino acid residues mutated in this study.
Figure 3.

Inhibitory potential of factor Va heavy chain peptides. (A) Inhibition of prothrombinase activity. Increasing concentrations of AP5, AP6, P15H, IWDYA, and AP5DY→KF were preincubated with factor Xa and assayed for prothrombinase activity as described in . P15H (◼) represents the control peptide containing amino acids 337−351 of the human factor Va heavy chain (see Figure 2 for details). AP5 (◆) represents amino acids 327−336 of factor Va. AP5DY→KF (△) represents amino acid residues 327−336 with residues 334 and 335 mutated from D and Y to K and F, respectively. IWDYA (◻) represents amino acid residues 332−336 of the human factor Va heavy chain. Data for AP6 are shown with ●. The concentration of peptide given on the x axis represents its final concentration in the prothrombinase mixture. The data represent the average of the results found in three independent experiments. The apparent inhibition constant (Ki) reported in the text is the value calculated from the formula IC50 = Ki(1 + S0/Km) (53), where Km is the Michaelis−Menten constant of the reaction in the absence of inhibitor, S0 is the concentration of prothrombin used, and IC50 is the half-maximal inhibition of prothrombinase by a given peptide. (B) Inhibition of prothrombinase assembly. Prothrombinase was assembled with [OG488]-EGR-hXa (10 nM), PCPS vesicles (10 µM), and increasing concentrations of human factor Va (up to 25 nM) in the presence of 2 mM Ca2+. Once a plateau was obtained, the preformed complexes were titrated with increasing concentrations of P15H (◼), AP3 (▲), AP4′ (▼), AP5 (◆), and AP6 (●). Δr was calculated as described in and plotted as extensively detailed previously (23). The data represent all the data points from two independent experiments performed in triplicate.
The mechanism of prothrombinase inhibition by AP5 was further investigated by examining the effect of the peptide on the kinetic parameters of prothrombinase assembly and function (Km and Vmax) in the presence of varying concentrations of the inhibitor (Figure 4). In the presence of increasing concentrations of peptide, both the apparent Km and Vmax changed; the inhibitor lowered the Vmax and increased the Km (Figure 4A). Analysis of the kinetic data using the Dixon plots demonstrated that AP5 inhibition of prothrombinase followed a model of linear mixed-type inhibition (Figure 4B). The apparent Ki of prothrombinase extrapolated from the graph was 10.5 µM (Figure 4B) which is in good agreement with the Ki value obtained from the IC50 value determined above (5.9 µM). The inhibition model of a mixed-type inhibitor is defined as unequal binding of the inhibitor to E and ES and suggests that AP5 that contains a portion of the factor Xa binding site may also interact with prothrombin in association with prothrombinase (53). However, it is also possible that binding of prothrombin to prothrombinase alters the affinity of AP5 for the enzyme.
Figure 4.

Kinetic analyses of prothrombin activation by prothrombinase in the presence of AP5. (A) Michaelis−Menten plots. Prothrombin generation experiments were performed in the absence (◼) and presence of increasing concentrations of AP5 [10 (▼), 15 (◆), and 20 µM (▲)], as described in using prothrombin concentrations varying from 40 nM to 1 µM. Initial rates of thrombin formation are plotted as a function of substrate concentration. (B) Analysis of the data using the Dixon plots. The data were analyzed and plotted as 1/Vo as a function of inhibitor concentration (AP5, Dixon plots). For mixed-type inhibition, a Dixon plot of 1/Vo as a function of increasing concentration of inhibitor is linear at a fixed enzyme and substrate concentration (53). The apparent inhibition constant (Ki) reported in the text is the value derived from the intercept of each of the four graphs (∼10.5 µM). The lines drawn represent the best fit through the points with an R2 varying from 0.9812 (worst) to 0.9881 (best). The following concentrations of prothrombin were used in the experiments: 80 (◆), 100 (●), 150 (◻), and 200 nM (△).
In view of these results, we have synthesized an additional peptide (IWDYA) and assayed its ability to inhibit prothrombinase activity. Peptide IWDYA inhibited cofactor activity, although not completely [Figure 3A (◻)]. Even at 500 µM peptide there was still approximately 25% cofactor activity remaining (not shown). The data demonstrated that IWDYA has an IC50 of 130 µM which corresponds to a Ki of 69 µM. Interestingly, AP6 [Figure 3A (●)] has an IC50 of ∼120 µM corresponding to a Ki of 64 µM. The similar Ki values between AP6 and IWDYA suggest that the inhibitory effect of the peptides is a consequence of the shared amino acid sequence 332IWDYA336 [Figure 2 (boxed sequence)]. However, it is important to note that the data provided in Figure 3A also demonstrate that while AP6 yields a Ki value similar to that of pentapeptide IWDYA for prothrombinase inhibition (∼60 µM), AP5 which also shares this sequence shows a 10-fold lower Ki value (∼6 µM). These data suggest that amino acid residues NH2-terminal to pentapeptide IWDYA are functionally important. This conclusion is strengthened by our earlier findings demonstrating the important contribution of amino acid stretch 227AAEEV331 to factor Va−factor Xa interaction (29). More recently, we have provided strong evidence supporting the notion that amino acids Glu330 and Val331 are significant contributors to prothrombinase complex function (31).
To identify the important amino acid residues of this region for cofactor activity, we have synthesized another peptide with the sequence DY replaced by KF (AP5DY→KF, AP5m) and assayed the peptide for prothrombinase inhibition. We have arbitrarily chosen to mutate these two amino acids because they are the only charged amino acid residues of this pentapeptide. However, the function of the other two amino acids (I332W333) was also established using recombinant technology and shown below. AP5m had an only weak effect on prothrombinase function with ∼60% cofactor activity remaining at concentrations of peptide as high as 500 µM [Figure 3A (△)], with a Ki value of ∼530 µM (assuming an IC50 of ∼1 mM). These data indicate that amino acid region 334–335 is important for the expression of the inhibitory potential of AP5. Taken together, the results indicate that (1) peptide AP5 from the central portion of the factor Va heavy chain is a potent inhibitor of factor Va cofactor activity and (2) amino acid region 323−335 (13 amino acids) from the factor Va heavy chain plays an important role in the optimal expression of prothrombinase activity (Figure 2).
Expression and Activation of Recombinant Human Factor V Molecules
We next employed a recombinant protein scheme to further asses the importance of amino acid region 334–335 of the factor Va heavy chain for cofactor activity. We used one charge reversal mutation (D334K) and one conservative mutation (Y335F) to obtain recombinant factor VKF. We have also replaced both amino acids with alanine to produce factor VAA. In addition, we have combined these mutations with mutations at the extremities of the factor Xa binding site on factor Va. We have thus obtained the quadruple mutants, factor VMI/KF, factor VFF/KF, factor VMI/AA, and factor VFF/AA (see Figure 1 for more details) (31). To assess the importance of the other two amino acids of peptide IWDYA, i.e., I332 and W333, for cofactor function, we have created two additional mutant molecules with these amino acids changed to alanine (factor VI332A and factor VW333A). All eight recombinant factor V molecules along with wild-type factor V (factor VWT) were expressed, purified, and assayed in a two-stage clotting assay, and the results are reported in Table 1. Factor VaWT displayed normal clotting activity, compared to plasma-derived factor Va. Conversely, while factor VaKF and factor VaAA had 5-fold reduced clotting activity under similar experimental conditions, all the quadruple mutants were practically devoid of clotting activity (≤30 units/mg). We have previously demonstrated that factor VaFF and factor VaMI are impaired in their interaction with factor Xa (31). Results with these mutants are reported in Table 1 for comparison. The data demonstrate that while mutation of amino acid region 334−335 impair clotting activity, these mutations alone are not enough to completely abolish factor Va clotting activity. However, combining these mutations with mutations at the extremities of the previously described factor Xa binding site results in cofactor molecules that are devoid of clotting activity. The clotting data obtained with the recombinant proteins also demonstrate that amino acids I332 and W333 are not essential for expression of factor Va clotting activity as the mutant molecules have clotting capabilities comparable to that of the wild-type molecule.
Effect of Recombinant Factor Va Molecules on Prothrombin Activation by Prothrombinase
The capability of the recombinant factor Va molecules to be incorporated into prothrombinase and activate prothrombin was investigated by gel electrophoresis. Prothrombinase assembled with factor VaWT displayed normal activation pattern compared to the plasma-derived cofactor with initial cleavage at Arg320 producing fragment 1·2-A (Figure S2), indicative of meizothrombin generation. Activation of prothrombin by prothrombinase assembled with recombinant purified factor VaKF or factor VaAA showed delayed thrombin formation as evidenced by delayed formation of both fragment 1·2-A and B chain of thrombin (Figure S2). Under similar experimental conditions, prothrombinase assembled with factor VaFF/KF, factor VaMI/KF, factor VaFF/AA, and factor VaMI/AA did not activate prothrombin to any significant extent, following a 3 h incubation period (data not shown). These results indicate that substitution of amino acids 334 and 335 of the factor Va heavy chain results in delayed prothrombin activation by factor Xa within prothrombinase assembled with factor VaKF or factor VaAA.
Kinetic Analyses of Prothrombinase Assembled with Recombinant Factor Va
The ability of the recombinant factor Va molecules to be incorporated into prothrombinase was investigated using an assay employing purified reagents. Under similar experimental conditions, factor VaI332A, factor VaW333A, factor VaKF, and factor VaAA displayed KD values comparable to the value obtained with the wild-type molecule (Table 1). Conversely, the affinities of factor VaFF/KF, factor VaFF/AA, factor VaMI/KF, and factor VaMI/AA were 6-, 10-, 23-, and 20-fold, respectively, lower than the affinity of factor VaWT for factor Xa (Table 1). These data demonstrate that amino acids 332−335 do not participate in the interaction of the cofactor with factor Xa.
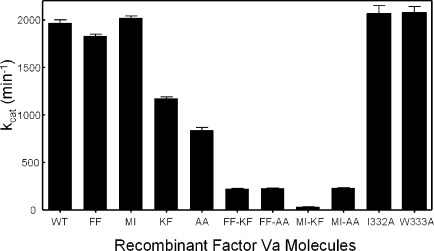
Figure 5 and Table 1 show the original raw data (Figure 5A), the apparent kcat (Figure 5B), the apparent Km (Table 1), and the second-order rate constants (Figure 5C) of prothrombinase assembled with the recombinant factor Va molecules. Knowing the KD of each recombinant molecule for factor Xa (Table 1), we calculated the appropriate amount of factor Va to saturate factor Xa using the quadratic equation (eq 1) described in the literature (36,37) and provided in . Thus, the titrations were performed under conditions where all factor Xa was saturated with factor Va, and as a consequence, any observed deficiency in the catalytic activity of prothrombinase was attributed to a defective factor Va molecule rather than to an impaired factor Va−factor Xa interaction. The data demonstrate that prothrombinase assembled with purified factor VaKF and factor VaAA was characterized by an ∼10-fold decrease in second-order rate constants (Figure 5B,C) compared to prothrombinase assembled with factor VaWT. Prothrombinase assembled with the quadruple mutant molecules, factor VaMI/KF, factor VaFF/KF, factor VaMI/AA, and factor VaFF/AA, demonstrated a significant decrease in the kcat and the second-order rate constant of the enzyme [up to 400-fold decrease (Figure 5C)]. The kinetic data obtained with the recombinant proteins also demonstrate that amino acids I332 and W333 are not required for expression of factor Va cofactor activity since the kinetic constants obtained with prothrombinase assembled with these two mutant molecules are similar to the data obtained with prothrombinase assembled with wild-type factor Va. Overall, the data show that substitution of amino acids 334 and 335 in factor Va has an effect on the catalytic efficiency of prothrombinase.
Figure 5.

Prothrombin titrations to determine the kinetic parameters of prothrombinase assembled with the various recombinant factor Va species. Thrombin generation experiments were carried out as described in by varying the substrate concentration. The raw data (hyperbolas) are shown in panel A: (◼ and ◻) two different preparations of wild-type factor Va, (○) factor VaAA, (●) factor VaFF-KF, (▽ and ▼) two different preparations of factor VaKF, (crossed boxes) factor VaMI-AA, (◆) factor VaMI-KF, (◼) factor VaFF-AA, (+) factor VaI332A, and (×) factor VaW333A. The Km values for all cofactor molecules are given in Table 1, while the kcat values are shown in panel B (min−1). Panel C provides the second-order rate constants (M−1 s−1). The data represent the average values found in at least three different titrations for each recombinant factor Va molecule.
Effect of the Mutations on Prothrombin Catalysis
Prothrombinase is composed of factor Va and factor Xa assembled on a membrane surface in the presence of divalent metal ions. We can thus hypothesize that prothrombinase is an enzyme composed of two subunits: a catalytic subunit (factor Xa) and a regulatory subunit (factor Va). Any perturbation in the interaction between the two subunits or any perturbations in the interaction of prothrombinase with the substrate caused by a mutation in the regulatory subunit may influence (modify) the stability of the catalytic site of the catalytic subunit. Thus, the consequences of mutations in factor Va affecting factor Xa catalytic efficiency can be measured relative to the change in the transition-state stabilization free energy of the enzyme. The kinetic data showed that the DY → KF/AA mutation in factor Va results in a decrease in the catalytic efficiency of factor Va which in turn is translated by positive values of ΔΔG⧧WT→KF and ΔΔG⧧WT→AA of 1.24 and 1.5 kcal/mol, respectively (Scheme 1). To quantify the interaction between these mutations and the mutations in the previously described factor Xa binding site (31) and their synergistic effects on prothrombinase function, we have calculated the difference in free energy of the transition-state analogue (ΔΔGint) for each of the quadruple mutants according to eqs 3 and 4. We have also constructed a thermodynamic cycle of prothrombinase (Scheme 1). The positive value of ΔΔGint for the combination of mutations at amino acids 323 and 324 and amino acids 330 and 331 with the DY → KF mutation indicates that these substitutions in the heavy chain of factor Va are detrimental to the catalytic activity of prothrombinase.
Scheme 1. Thermodynamic Cycle for Prothrombinase Activity following Substitutions in the Factor Va Heavy Chain.

ΔΔGint is the free energy of interaction between the side chains of D334Y335 and E323Y324/E330V331 of the factor Va heavy chain and was calculated with eqs 3 and 4 described in . The values illustrated on the arrows are the ΔΔG⧧ values between recombinant species as indicated. Parallel arrows show the difference in catalytic efficiency between prothrombinase assembled with different mutant factor Va molecules. Different forms of prothrombinase are compared for their ability to activate prothrombin.
Structural Consequences of the Mutations
We next used the model of the factor Va molecule to understand the structural consequences of the mutations at positions D334 and Y335. It is important to underline that our MD simulations are tentative, especially with reference to interactions within the A2 domain, as this region is not present in any crystal structures and is derived from modeling (19). The MD simulation of factor VaWT reached equilibrium after 1 ns and was extended for an additional 1 ns for analysis. The simulation of factor VaKF reached equilibrium after 2.5 ns, and the simulation was extended to 3.5 ns for analysis of the last equilibrated nanosecond. The simulation of factor VaAA reached equilibrium at 2 ns, and the last 500 ps was used for analysis. Figure 6A shows the final snapshot of factor VaWT with amino acid residues D334 and Y335 shown as blue sticks. The corresponding amino acids in factor VaKF [K334 and F335, respectively (Figure 6B)] and in factor VaAA [A334 and A335, respectively (Figure 6C)] are also represented as blue sticks. The MD data show that the antiparallel β-sheet contained in the region from R321 to V331 (first strand) and K365 to T369 (second strand) is preserved in the case of factor VaWT during the simulation. In contrast, in the case of factor VaAA and factor VaKF, the β-sheets are extended at the V331 and K365 ends by three amino acids. This β-sheet contains the critical amino acid sequence 323−331 previously shown to regulate factor Va cofactor activity (22,23,29). Elongation of the R321−V331/K365−T369 β-sheet to R321−A334/H362−T369 in the factor VaAA molecule positioned H362 in a conformation that ensured hydrogen bonding with F538, thus directly affecting the stability of the S528−F538 α-helix. The S528−F538 helix is a continuation of the amino acid stretch that contains a secondary binding site of the cofactor for factor Xa that is located between amino acids 493 and 506 (24).
Figure 6.

Structural analyses of the factor V heavy chain. A portion of the prothrombin interactive site containing amino acids D695−Q69930,54,55 is shown as magenta spheres. The α-helix contained in S528−F538 is colored red. This region contains a secondary binding site for factor Xa (24). The β-sheet contained in the R321−V331/K365−T369 sequence that is important for expression of factor Va cofactor activity (22,23,29) is colored yellow. The loop contained within the sequence of amino acid residues 400−420 is shown with cyan ribbons. Amino acid residues F538 and H362 are shown as green sticks. S412 is shown as cyan sticks. N534 and K364 are shown as lime sticks. Amino acid residues Y530 and E330 are shown as olive sticks, and E323 is shown as orange sticks. The final snapshot at 2 ns of factor VaWT is shown in panel A with amino acid residues D334 and Y335 represented as blue sticks. Panel B shows the final snapshot at 3.5 ns of factor VaKF with the mutated amino acid residues K334 and Phe335 as blue sticks and R413 and S412 as cyan sticks. Panel C is a snapshot at 2 ns of factor VaAA with the mutated amino acid residues A334 and A335 colored blue.
Discussion
Collectively, through a systematic approach, using both kinetic studies with synthetic peptides and recombinant proteins, our data demonstrate that amino acid region 334−335 plays an important role in the expression of factor Va cofactor activity. Our data also demonstrate the cofactor requirement for the efficient rearrangement of enzyme (factor Xa) and substrate (prothrombin) within prothrombinase necessary for efficient catalysis of prothrombin at two spatially distinct sites. However, it is important to note that our study does not provide a definitive mechanism detailing the involvement of amino acid region 334−335 of factor Va in cofactor activity.
The mechanism of inhibition of prothrombinase function by AP5, AP5DY→KF, AP6, and IWDYA was investigated by assessing factor Va cofactor activity in the presence of increasing concentrations of synthetic peptide. These data show that the IWDYA motif shared by AP5 and AP6 contains the amino acids that account for the inhibitory effect of the peptides. This is evident in the similar Ki values of peptides AP6 and IWDYA. AP5 has an ∼10-fold lower Ki value than AP6 and IWDYA, because it also contains amino acids Glu330 and Val331, which have been previously shown to be involved in factor Xa binding (29). Analysis of the mode of inhibition of prothrombinase by AP5 suggests that the peptide is a mixed-type inhibitor and interacts with both prothrombinase and prothrombinase bound to the substrate (prothrombin). Thus, while it is possible that AP5 interacts with prothrombinase in the presence and absence of prothrombin, the possibility that AP5 also binds prothrombin when the enzyme is in complex with the substrate must be kept in mind. Finally, within the peptide sequence IWDYA, amino acids D and Y are contributing to its function, since substitution of these two amino acids with K and F, respectively (AP5DY→KF), results in the almost complete loss of the inhibitory potential of AP5.
Site-directed mutagenesis was used to assess the importance of amino acid region 334−335 from the factor Va heavy chain during prothrombin catalysis. The data show that mutating this amino acid region results in a factor Va molecule that when incorporated into prothrombinase produces an enzyme with an overall 10-fold decrease in its second-order rate constant. Since the KD values of factor VaKF and factor VaAA for factor Xa are similar to the KD of factor VaWT for the enzyme, the data strongly suggest that the inability of prothrombinase assembled with factor VaKF and factor VaAA to function optimally can be explained by the inability of factor Xa to efficiently convert prothrombin to thrombin because of a diminished number of productive collisions.
The results obtained from the MD simulations of wild-type factor Va, factor VaKF, and factor VaAA were able to offer an explanation both for the similar KD values found for the interaction of wild-type factor Va and factor VaKF (or factor VaAA) with factor Xa and for the impaired clotting and cofactor activity of both mutant molecules. The modeling data demonstrate that the distances between the Cα atoms of amino acids 334 and 323, 334 and 324, 335 and 323, and 335 and 324 analyzed separately do not change significantly when comparing wild-type factor Va with the mutant molecules. Thus, there are no changes in the overall conformation of the segment of amino acids 323−331 because of the mutations. The combined data rather suggest that the difference in the distance of the centers of mass between the two amino acid segments observed is due to the presence of different amino acid side chains that interact with each other. Coincidentally, the thermodynamic and kinetic data obtained herein also suggest that combination of the mutations at the extremities of the factor Xa binding site in factor Va, 323EY324 (factor VaFF) and 330EV331 (factor VaMI), with mutations in the 334DY335 region (factor VaKF or factor VaAA) have an additive detrimental effect on prothrombinase with respect to prothrombin activation. It is also important to note that replacement of an aspartic acid with an alanine appeared to be more detrimental to cofactor activity than replacement with lysine. These data would suggest that the negative charge per se at residue 334 is not critical for cofactor activity. Finally, we must point out that tentative explanations using our theoretical model are suggestive rather than conclusive and await verification that will be obtained from the crystal structure of the recombinant mutated cofactor molecule.
At this point, it is important to underline the complementarities of all the data provided in this paper. Our findings using the MD simulations demonstrate that in the presence of the mutations there is a relocation of the COOH-terminal region of factor VaKF and factor VaAA as compared with the wild-type cofactor molecule. However, these calculations are unable to distinguish whether the changes will enhance or decrease the catalytic efficiency of the prothrombinase complex assembled with the mutated cofactor molecule. On the other hand, the kinetic, thermodynamic, and molecular biology data demonstrate a detrimental effect of the mutations on cofactor activity without providing a structural explanation. Together, the data suggest that the internal rearrangement at the COOH terminus of factor VaKF and factor VaAA is detrimental to the overall catalytic efficiency of factor Xa as part of prothrombinase. Therefore, the MD simulations using the only complete structure of factor Va in solution available in the literature are useful in providing a theoretical explanation for the decrease in the second-order rate constant of prothrombinase assembled with the mutant cofactor molecules.
Acknowledgments
We thank Dr. Ken Mann and Dr. Tom Orfeo from the University of Vermont for providing the monoclonal antibodies to factor V and Dr. Alex Kurosky and Steve Smith from the University of Texas, Medical Branch, for quantitative amino acid analysis and mass spectrometry analyses of the synthetic peptides.
Supporting Information Available
Figure S1 shows the kinetic analyses of prothrombin activation by prothrombinase in the presence of AP3. Panel A shows the Michaelis−Menten plots, while panel B shows the mathematical transformation of the data using Dixon plots. Figure S2 shows prothrombin activation by prothrombinase assembled with recombinant factor Va molecules. This material is available free of charge via the Internet at http://pubs.acs.org.
Funding Statement
National Institutes of Health, United States
Footnotes
Abbreviations: PS, l-α-phosphatidylserine; PC, l-α-phosphatidylcholine; PCPS, small unilamellar phospholipid vesicles composed of 75% PC and 25% PS (w/w); HPLC, high-performance liquid chromatography; SDS-PAGE, sodium dodecyl sulfate−polyacrylamide gel electrophoresis; PVDF, polyvinylidene difluoride; [OG488]-EGR-hXa, factor Xa labeled in the active site with Oregon Green 488; factor VaWT, recombinant wild-type human factor Va; factor VaKF, recombinant human factor Va with the mutations D334 → K and Y335 → F; factor VaAA, recombinant human factor Va with the mutations D334 → A and Y335 → A; factor VaFF, recombinant human factor Va with the mutations E323 → F and Y324 → F; factor VaMI, recombinant human factor Va with the mutations E330 → M and V331 → I; factor VI332A, recombinant human factor Va with the mutation I332 → A; factor VW333A, recombinant human factor Va with the mutation W333 → A; factor VaFF/KF, quadruple mutant of recombinant human factor Va with the mutations E323 → F, Y324 → F, D334 → K, and Y335 → F; factor VaMI/KF, quadruple mutant of recombinant human factor Va with the mutations E330 → M, V331 → I, D334 → K, and Y335 → F; factor VaFF/AA, quadruple mutant of recombinant human factor Va with the mutations E323 → F, Y324 → F, D334 → A, and Y335 → A; factor VaMI/AA, quadruple mutant of recombinant human factor Va with the mutations E330 → M, V331 → I, D334 → A, and Y335 → A.
Supplementary Material
References
- Kalafatis M.; Egan J. O.; van’t Veer C.; Cawthern K. M.; Mann K. G. (1997) The regulation of clotting factors. Crit. Rev. Eukaryotic Gene Expression 7, 241–280. [DOI] [PubMed] [Google Scholar]
- Kalafatis M.; Mann K. G. (2001) Factor V: Dr. Jeckyll and Mr. Hyde. Adv. Exp. Med. Biol. 489, 31–43. [DOI] [PubMed] [Google Scholar]
- Rosing J.; Zwaal R. F.; Tans G. (1986) Formation of meizothrombin as intermediate in factor Xa-catalyzed prothrombin activation. J. Biol. Chem. 261, 4224–4228. [PubMed] [Google Scholar]
- Owen W. G.; Esmon C. T.; Jackson C. M. (1974) The conversion of prothrombin to thrombin. I. Characterization of the reaction products formed during the activation of bovine prothrombin. J. Biol. Chem. 249, 594–605. [PubMed] [Google Scholar]
- Esmon C. T.; Owen W. G.; Jackson C. M. (1974) The conversion of prothrombin to thrombin. II. Differentiation between thrombin- and factor Xa-catalyzed proteolyses. J. Biol. Chem. 249, 606–611. [PubMed] [Google Scholar]
- Esmon C. T.; Jackson C. M. (1974) The conversion of prothrombin to thrombin. III. The factor Xa-catalyzed activation of prothrombin. J. Biol. Chem. 249, 7782–7790. [PubMed] [Google Scholar]
- Krishnaswamy S.; Church W. R.; Nesheim M. E.; Mann K. G. (1987) Activation of human prothrombin by human prothrombinase. Influence of factor Va on the reaction mechanism. J. Biol. Chem. 262, 3291–3299. [PubMed] [Google Scholar]
- Nesheim M. E.; Taswell J. B.; Mann K. G. (1979) The contribution of bovine Factor V and Factor Va to the activity of prothrombinase. J. Biol. Chem. 254, 10952–10962. [PubMed] [Google Scholar]
- Nesheim M. E.; Mann K. G. (1983) The kinetics and cofactor dependence of the two cleavages involved in prothrombin activation. J. Biol. Chem. 258, 5386–5391. [PubMed] [Google Scholar]
- Esmon C. T.; Owen W. G.; Jackson C. M. (1974) A plausible mechanism for prothrombin activation by factor Xa, factor Va, phospholipid, and calcium ions. J. Biol. Chem. 249, 8045–8047. [PubMed] [Google Scholar]
- Downing M. R.; Butkowski R. J.; Clark M. M.; Mann K. G. (1975) Human prothrombin activation. J. Biol. Chem. 250, 8897–8906. [PubMed] [Google Scholar]
- Butkowski R. J.; Elion J.; Downing M. R.; Mann K. G. (1977) Primary structure of human prethrombin 2 and α-thrombin. J. Biol. Chem. 252, 4942–4957. [PubMed] [Google Scholar]
- Bajaj S. P.; Butkowski R. J.; Mann K. G. (1975) Prothrombin fragments. Ca2+ binding and activation kinetics. J. Biol. Chem. 250, 2150–2156. [PubMed] [Google Scholar]
- Myrmel K. H.; Lundblad R. L.; Mann K. G. (1976) Characteristics of the association between prothrombin fragment 2 and α-thrombin. Biochemistry 15, 1767–1773. [DOI] [PubMed] [Google Scholar]
- Brufatto N.; Nesheim M. E. (2003) Analysis of the kinetics of prothrombin activation and evidence that two equilibrating forms of prothrombinase are involved in the process. J. Biol. Chem. 278, 6755–6764. [DOI] [PubMed] [Google Scholar]
- Suzuki K.; Dahlback B.; Stenflo J. (1982) Thrombin-catalyzed activation of human coagulation factor V. J. Biol. Chem. 257, 6556–6564. [PubMed] [Google Scholar]
- Nesheim M. E.; Foster W. B.; Hewick R.; Mann K. G. (1984) Characterization of Factor V activation intermediates. J. Biol. Chem. 259, 3187–3196. [PubMed] [Google Scholar]
- Adams T. E.; Hockin M. F.; Mann K. G.; Everse S. J. (2004) The crystal structure of activated protein C-inactivated bovine factor Va: Implications for cofactor function. Proc. Natl. Acad. Sci. U.S.A. 101, 8918–8923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Orban T.; Kalafatis M.; Gogonea V. (2005) Completed three-dimensional model of human coagulation factor Va. Molecular dynamics simulations and structural analyses. Biochemistry 44, 13082–13090. [DOI] [PubMed] [Google Scholar]
- Kalafatis M.; Xue J.; Lawler C. M.; Mann K. G. (1994) Contribution of the heavy and light chains of factor Va to the interaction with factor Xa. Biochemistry 33, 6538–6545. [DOI] [PubMed] [Google Scholar]
- Luckow E. A.; Lyons D. A.; Ridgeway T. M.; Esmon C. T.; Laue T. M. (1989) Interaction of clotting factor V heavy chain with prothrombin and prethrombin 1 and role of activated protein C in regulating this interaction: Analysis by analytical ultracentrifugation. Biochemistry 28, 2348–2354. [DOI] [PubMed] [Google Scholar]
- Kalafatis M.; Mann K. G. (2001) The Role of the Membrane in the Inactivation of Factor Va by Plasmin: Amino Acid Region 307−348 of Factor V Plays a Critical Role in Factor Va Cofactor Function. J. Biol. Chem. 276, 18614–18623. [DOI] [PubMed] [Google Scholar]
- Kalafatis M.; Beck D. O. (2002) Identification of a binding site for blood coagulation factor Xa on the heavy chain of factor Va. Amino acid residues 323−331 of factor V represent an interactive site for activated factor X. Biochemistry 41, 12715–12728. [DOI] [PubMed] [Google Scholar]
- Heeb M. J.; Kojima Y.; Hackeng T. M.; Griffin J. H. (1996) Binding sites for blood coagulation factor Xa and protein S involving residues 493−506 in factor Va. Protein Sci. 5, 1883–1889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jenny R. J.; Pittman D. D.; Toole J. J.; Kriz R. W.; Aldape R. A.; Hewick R. M.; Kaufman R. J.; Mann K. G. (1987) Complete cDNA and derived amino acid sequence of human factor V. Proc. Natl. Acad. Sci. U.S.A. 84, 4846–4850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guinto E. R.; Esmon C. T.; Mann K. G.; MacGillivray R. T. (1992) The complete cDNA sequence of bovine coagulation factor V. J. Biol. Chem. 267, 2971–2978. [PubMed] [Google Scholar]
- Yang T. L.; Cui J.; Rehumtulla A.; Yang A.; Moussalli M.; Kaufman R. J.; Ginsburg D. (1998) The structure and function of murine factor V and its inactivation by protein C. Blood 91, 4593–4599. [PubMed] [Google Scholar]
- Grimm D. R.; Colter M. B.; Braunschweig M.; Alexander L. J.; Neame P. J.; Kim H. K. (2001) Porcine factor V: cDNA cloning, gene mapping, three-dimensional protein modeling of membrane binding sites and comparative anatomy of domains. Cell. Mol. Life Sci. 58, 148–159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh L. S.; Bukys M. A.; Beck D. O.; Kalafatis M. (2003) Amino acids Glu323, Tyr324, Glu330, and Val331 of factor Va heavy chain are essential for expression of cofactor activity. J. Biol. Chem. 278, 28335–28345. [DOI] [PubMed] [Google Scholar]
- Bukys M.; Kim P. Y.; Nesheim M. E.; Kalafatis M. (2006) A Control Switch for Prothrombinase. Characterization of a Hirudin-Like Pentapeptide From the COOH Terminus of Factor Va Heavy Chain that Regulates the Rate and Pathway for Prothrombin Activation. J. Biol. Chem. 281, 39194–39204. [DOI] [PubMed] [Google Scholar]
- Bukys M. A.; Blum M. A.; Kim P. Y.; Brufatto N.; Nesheim M. E.; Kalafatis M. (2005) Incorporation of factor Va into prothrombinase is required for coordinated cleavage of prothrombin by factor Xa. J. Biol. Chem. 280, 27393–27401. [DOI] [PubMed] [Google Scholar]
- Erdogan E.; Bukys M. A.; Orfeo T.; Mann K. G.; Kalafatis M. (2007) Identification of an inactivating cleavage site for α-thrombin on the heavy chain of factor Va. Thromb. Haemostasis 98, 998–1006. [PubMed] [Google Scholar]
- Laemmli U. K. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227, 680–685. [DOI] [PubMed] [Google Scholar]
- Towbin H.; Staehelin T.; Gordon J. (1979) Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. U.S.A. 76, 4350–4354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Erdogan E.; Bukys M. A.; Kalafatis M. (2008) The contribution of amino acid residues 1508−1515 of factor V to light chain generation. J. Thromb. Haemostasis 6, 118–124. [DOI] [PubMed] [Google Scholar]
- Krishnaswamy S. (1992) The interaction of human factor VIIa with tissue factor. J. Biol. Chem. 267, 23696–23706. [PubMed] [Google Scholar]
- Krishnaswamy S.; Williams E. B.; Mann K. G. (1986) The binding of activated protein C to factors V and Va. J. Biol. Chem. 261, 9684–9693. [PubMed] [Google Scholar]
- Ackers G. K.; Smith F. R. (1985) Effects of site-specific amino acid modification on protein interactions and biological function. Annu. Rev. Biochem. 54, 597–629. [DOI] [PubMed] [Google Scholar]
- LiCata V. J.; Ackers G. K. (1995) Long-range, small magnitude nonadditivity of mutational effects in proteins. Biochemistry 34, 3133–3139. [DOI] [PubMed] [Google Scholar]
- Wells J. A. (1990) Additivity of mutational effects in proteins. Biochemistry 29, 8509–8517. [DOI] [PubMed] [Google Scholar]
- Mildvan A. S.; Weber D. J.; Kuliopulos A. (1992) Quantitative interpretations of double mutations of enzymes. Arch. Biochem. Biophys. 294, 327–340. [DOI] [PubMed] [Google Scholar]
- Mildvan A. S. (2004) Inverse thinking about double mutants of enzymes. Biochemistry 43, 14517–14520. [DOI] [PubMed] [Google Scholar]
- Guex N.; Peitsch M. C. (1997) SWISS-MODEL and the Swiss-PdbViewer: An environment for comparative protein modeling. Electrophoresis 18, 2714–2723. [DOI] [PubMed] [Google Scholar]
- Berendsen H. J. C.; van der Spoel D.; van Drunen R. (1995) GROMACS: A message-passing parallel molecular dynamics implementation. Comput. Phys. Commun. 91, 43–56. [Google Scholar]
- Lindahl E.; Hess B.; van der Spoel D. (2001) GROMACS 3.0: A package for molecular simulation and trajectory analysis. J. Mol. Model. 7, 306–317. [Google Scholar]
- Berendsen H. J. C., Postma J. P. M., van Gunsteren W. F., and Hermans J. (1981) Interaction models for water in relation to protein hydration, in Intermolecular Forces (Pullman B., and Reidel D., Eds.) pp 331−342, Kluwer Academic Publishers, Dordrecht, The Netherlands. [Google Scholar]
- van Gunsteren W. F., and Berendsen H. J. C. (1987) Gromos-87 Manual, Biomos BV, Groningen, The Netherlands. [Google Scholar]
- Hess B.; Bekker H.; Berendsen H. J. C.; Fraaije J. G. E. M. (1997) LINCS: A linear constraint solver for molecular simulations. J. Comput. Chem. 18, 1463–1472. [Google Scholar]
- Darden T.; York D.; Pedersen L. (1993) Particle mesh Ewald: An N·log(N) method for Ewald sums in large systems. J. Chem. Phys. 98, 10089–10092. [Google Scholar]
- Essmann U.; Perera L.; Berkowitz M. L.; Darden T.; Lee H.; Pedersen L. G. (1995) A smooth particle mesh Ewald method. J. Chem. Phys. 103, 8577–8593. [Google Scholar]
- Berendsen H. J. C.; Postma J. P. M.; van Gunsteren W. F.; DiNola A.; Haak J. R. (1984) Molecular dynamics with coupling to an external bath. J. Chem. Phys. 81, 3684–3690. [Google Scholar]
- Parrinello M.; Rahman A. (1980) Structure and pair potentials: A molecular-dynamics study. Phys. Rev. Lett. 45, 1196–1199. [Google Scholar]
- Segal I. H. (1993) Enzyme Kinetics, pp 170−178, John Wiley & Sons, Inc., New York. [Google Scholar]
- Kalafatis M.; Beck D. O.; Mann K. G. (2003) Structural requirements for expression of factor Va activity. J. Biol. Chem. 278, 33550–33561. [DOI] [PubMed] [Google Scholar]
- Beck D. O.; Bukys M. A.; Singh L. S.; Szabo K. A.; Kalafatis M. (2004) The contribution of amino acid region ASP695-TYR698 of factor V to procofactor activation and factor Va function. J. Biol. Chem. 279, 3084–3095. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.