

Abstract
Background
In this study we examined the karyotypes of morphologically indistinguishable populations of the electric knifefish Gymnotus carapo sensu stricto from the Eastern Amazon of Brazil. These were identified unambiguously on the basis of external morphology, meristics, and pigmentation.
Results
Specimens from one of five localities exhibited a karyotype previously not documented for Gymnotus species in the Amazon basin: 2n = 40 (34M/SM+6ST/A). Samples from the other four localities exhibited a different karyotype: 2n = 42 (30M/SM+12ST/A), which we had previously described. Specimens from all five localities presented constitutive heterochromatin in the centromeric region of almost all chromosomes, including in the distal and interstitial regions. Staining with 4'6-Diamidino-2-phenylindole revealed C-positive banding. In both karyotypes the Nucleolar Organizer Region (NOR) was located on the short arm of pair 20, and Chromomycin A3 stained the NORs. Fluorescent in situ hybridization with telomeric probes showed an Interstitial Telomeric Sequence (ITS) in the proximal short arm of a metacentric pair in the 2n = 40 karyotype.
Conclusion
The difference between the two karyotypes on the diploid number and chromosome morphology can be explained by rearrangements of the fusion-fission type and also by pericentric inversions. The presence of ITS in a metacentric pair of the 2n = 40 karyotype suggests that the difference in the diploid number of the karyotypes results from a fusion. The consistent 2n = 42 karyotype at four localities suggests an interbreeding population. However, because fusion-fission and pericentric inversions of this nature typically result in reproductive isolation, we speculate that the form with the 2n = 40 karyotype is a different species to that of the 2n = 42 form. Nonetheless, we did not observe evident differences in external morphology, meristics and pigmentation between the two forms, which suggest that they represent cryptic sympatric species in the G. carapo species complex. We speculate that the chromosomal speciation occurred recently, allowing insufficient time for the fixation of other differences following post-zygotic isolation.
Background
Gymnotus (Gymnotiformes, Gymnotidae) is the most diverse known Neotropical electric knife fish genus. It currently holds 33 valid described species and many additional undescribed species are known from museum collections [1-4]. Gymnotus has the ability to generate a pulsed electrostatic field from a specialized electric organ and detect electrostatic fields with electroreceptors. These Electric Organ Discharges (EODs) permit electrolocation, the detection of objects within the electrostatic field, and also electrocommunication (review in [5]; [3]).
The diploid number of chromosomes in Gymnotus has been documented to vary from 2n = 39–40 (with sex chromosomes of the type X1X2Y) to 2n = 54, exhibiting variation in the karyotype formula, the quantity of heterochromatin, and the position of the Nucleolar Organization Region (NOR) [6-9].
The species Gymnotus carapo (L.) sensu stricto, as currently defined [10], occurs over large areas of northern South America: in the Amazon and Orinoco basins, the coastal drainages of the Guyanas, and some coastal basins of Northeastern Brazil. The Linnaean syntypes of G. carapo were collected in the first half of the 18th Century near Paramaribo, Surinam [10].
A complex of additional morphologically similar or cryptic species that closely resemble G. carapo sensu stricto are known to extend as far south as Northern Argentina (Albert, Crampton, pers. obs). Likewise, some populations within the currently defined range of G. carapo sensu stricto may also prove to be distinct species within this species-complex. This G. carapo species-complex is not to be confused with the G. carapo species-group, which currently comprises 18 well-defined species in which there are two (vs. one) laterosensory pores in the dorsoposterior portion of the preopercle (and which includes G. carapo sensu stricto). G. carapo senso stricto can be distinguished from all other members of the G. carapo species-group by a combination of characters that are listed in the description of the species [10]. It has a distinct color pattern comprising 16–27 dark obliquely oriented pigment bands or pand-pairs, with irregular wavy margins, often broken into spots above the lateral line. This distinct pigmentation pattern is shared only with G. arapaima and G. diamantinensis, from which G. carapo senso stricto can be distinguished on the basis of morphological and meristic characters.
At least four new species from the G. carapo species-group are currently being described from southern Brazil, Uruguay and Argentina (Crampton, Albert, Cognato and Richer-de-Forges, in review). However, many taxonomic uncertainties will be resolved only by using a combination of traditional taxonomy (based on morphology), molecular systematic techniques, cytogenetic analysis, and an examination of the diversity of EODs.
Cytogenetic variation has probably played an important role in the diversification of species in the G. carapo species-complex. Previous studies have indicated that forms identified as G. carapo exhibit different karyotypes (e.g. 2n = 48 in Amazonas, 2n = 42 in Pará, 2n = 54 and 2n = 52 from Southern Brazil) (reviewed in [9]). Whether this diversity is the manifestation of cryptic species diversity in a G. carapo species-complex or, alternatively, intraspecific cytogenetic diversity is currently unknown. Nonetheless, this question is of great relevance to our understanding of diversification in the genus. In this contribution, which forms the second in a series on the evolutionary cytogenetics of Gymnotus, we describe a novel karyotype in a population of Gymnotus carapo from the Eastern Amazon. We speculate that this may have evolved from chromosomal rearrangement of the karyotype of a more common, sympatrically co-occurring form.
Methods
We performed cytogenetic analysis on 17 specimens of Gymnotus from five localities in Pará State, Brazil, in the Eastern Amazon basin (Table 1, Figure 1). Morphological measurements and meristic counts were undertaken from specimens fixed in 10% formaldehyde, and then preserved in 70% ethanol. Morphometric data were taken as point-to-point linear distances from standardized landmarks on the left side of adult specimens using digital calipers to the nearest mm. Protocols for measurements follow those of [10,11] and [12] and abbreviations are given in Table 2. Two additional morphometric measurements were included: Head depth measured vertically at the mid point of the eye and Eye diameter measured horizontally from the anterior to posterior extremities of the orbital margin. Morphological measurements as a proportion of TL were not included for analysis in specimens with damage to the caudal appendage exceeding an estimated 5% of intact TL. Meristic and scale count procedures follow [11], and abbreviations are given in Table 3. We refer to the vertical bars of Gymnotus as 'bands', as has become the standard in the literature on this genus. Principal component analysis of morphological and meristic data was undertaken using Statistica 7.1 (Statsoft, Tulsa, OK). The sex of each specimen was determined by dissection under a stereo microscope. Female ovaries contain yellowish eggs, while male testes are smooth and pinkish white.
Table 1.
Sample localities in the State of Pará, Brazil
| Site | Lot | N | 2n | Locality | Darainage | Geographical Coordinates |
| 1 | MPEG 13332, 15099 | 2M | 42 | Ponta de Pedra | Rio Marajó-Ité | 01° 20' 25.4" S, 048° 58' 06.2" W |
| 2 | MPEG 13333 | 1M | 42 | São Miguel do Guamá | Rio Guamá | 01° 32' 09.5" S, 047° 36' 18.7" W |
| 3 | MPEG 13331, 15098 | 4M, 2F | 42 | Capanema | Lago Segredo and Lago Açaiteua | 01° 07' 30" S, 047° 07' 30" W |
| 4 | MPEG 13330 | 1M | 42 | Benfica | Rio Murini | 01° 16' 34.8" S, 048° 20' 17.0" W |
| 5 | MPEG 13329, 15100 | 1M, 6F | 40 | Almeirim | Rio Amazonas | 01° 31' 34.2" S, 052° 33' 37.9" W |
N = number of specimens submitted to cytogenetic analysis; F = Female, M = Male.
Figure 1.

Map showing the localities of species Gymnotus with known cytogenetic formulas. Data are taken from a review of the cytogenetic literature by Milhomem et al. (2007), from Margarido et al. (2007) and Lacerda & Maistro (2007), and from the species discussed in this paper. See Table 4 for chromosome numbers of these species, and further locality information. At some localities the coordinates are approximate because detailed information was not available from the respective papers.
Table 2.
Morphometric data for adult specimens of two distinct karyotypic forms belonging to the Gymnotus carapo species complex from the Eastern Amazon (EA) (G. carapo 2n = 42 and G. carapo 2n = 40).
| G. carapo 2n = 42 | G. carapo 2n = 40 | G. carapo EA | ||||
| Range | Mean | Range | Mean | Range | Mean | |
| TL | 160–365 (9) |
- | 240–300 (7) |
- | 165–253 (16) |
- |
| HL | 29.0–42.8 (9) |
- | 29.56–38.7 (7) | - | 21.0–34.5 (16) |
- |
| HL %* | 11.7–12.7 (4) | 12.3 | 12.1–12.8 (3) | 12.4 | 11.7–13.6 (15) |
12.7 |
| PR % | 34.0–38.5 (9) |
35.6 | 32.6–38.5 (7) | 35.6 | 32.9–35.7 (16) |
34.2 |
| MW % | 39.7–47.5 (9) |
43.8 | 40.6–46.5 (7) |
42.9 | 41.0–46.1 (16) |
43.7 |
| PO % | 58.7–63.4 (9) | 61.2 | 60.3–62.5 (7) |
61.5 | 60.0–65.5 (16) |
62.3 |
| IO % | 33.7–40.2 (9) | 37.4 | 34.5–38.9 (7) |
36.8 | 34.4–40.6 (16) |
37.1 |
| BD %* | 11.0–14.3 (4) |
12.4 | 13.5–14.3 (3) |
13.8 | 8.7–12.4 (16) |
10.1 |
| BW %* | 52.3–77.1 (4) | 65.2 | 56.5–67.9 (3) |
63.0 | 6.2–8.6 (16) |
7.2 |
| BW/BD | 0.59–0.73 (9) |
0.65 | 0.55–0.61 (7) |
0.58 | 0.67–0.76 (16) |
0.72 |
| HD % | 55.3–64.8 (9) |
60.0 | 58.6–63.2 (7) | 60.5 | 50.8–60.9 (16) |
55.2 |
| HD2 % | 36.0–40.6 (9) |
38.4 | 37.4–42.3 (7) |
40.7 | NA | NA |
| HW % | 56.2–66.8 (9) |
60.7 | 54.2–59.7 (7) |
58.0 | 53.3–64.4 (16) |
56.3 |
| PA % | 63.0–88.2 (9) |
78.8 | 64.2–89.5 (7) |
75.6 | 76.5–99.2 (16) |
89.0 |
| BO% | 31.8–47.3 (9) | 39.3 | 34.2–46.4 (7) |
41.2 | 34.7–38.1 (16) |
36.5 |
| P1 % | 44.4–50.1 (9) |
47.8 | 46.0–58.6 (7) | 51.8 | 42.7–49.1 (16) |
45.7 |
| AF %* | 81.1–81.9 (4) | 81.4 | 80.0–80.7 (3) | 80.5 | 69.4–86.6 (16) |
75.9 |
| ED % | 7.0–10.2 (9) |
8.4 | 8.0–9.1 (7) |
8.5 | NA | NA |
For reference we include data for EA populations of G. carapo taken from [10].
Abbreviations: TL: total length; HL: head length; PR: preorbital length; MW: mouth width; PO: postorbital length; IO: interorbital distance; BD: body depth; BW: body width; HD: head depth at nape; HD2: head depth at middle of eye; HW: head width; PA: preanal distance; BO: branchial opening; P1: pectoral-fin length; AF: anal fin base length; ED: eye diameter. TL and HL expressed in mm. Percentage measurements in HL or: if marked with an asterisk: in TL. BW/BD expressed as a ratio. N values (in parentheses) vary because measurements were excluded from specimens with damage or unusual preservation artifacts. NA = not available.
Table 3.
Meristic data for adult specimens of two distinct karyotypic forms belonging to the Gymnotus carapo species complex from the Eastern Amazon (EA) (G. carapo 2n = 42 and G. carapo 2n = 40).
| G. carapo 2n = 42 | G. carapo 2n = 40 | G. carapo EA | ||||
| Range | Median* Mode** |
Range | Median* Mode** |
Range | Median* Mode** |
|
| BND | 19–27 (9) |
22* | 14–21 (7) |
18.5* | 21–25 (16) |
23* |
| AFR | 170–220 (3) |
196.7* | 190–225 (2) |
202.5* | 190–235 (10) |
222* |
| P1R | 13–16 (9) |
14** | 15–17 (7) |
15** | 13–15 (16) |
14** |
| SAL | 6–8 (9) |
7** | 6–8 (7) |
7** | 6–7 (16) |
7** |
| CEP | 3–4 (3) |
3** | 3–4 (3) |
3** | 3–4 (16) |
3** |
| APS | 7–9 (9) |
9** | 8–10 (7) |
9** | 7–8 (7) |
8** |
| PCV | 33 (1) |
- | 34 (1) |
- | 32–35 (9) |
33 |
| PLR | 43–51 (9) |
47* | 40–48 (7) |
45* | 41–54 (15) |
48* |
| PLL | 87–104 (4) |
93.5* | 84–89 (3) |
84 | 68–110 (15) |
77* |
| VLR | 7–9 (9) |
9** | 8–10 (7) |
9** | 0–8 (10) |
4** |
For reference we include data for EA populations of G. carapo taken from [10].
Abbreviations: BND: vertical bands; AFR: anal-fin rays; P1R: pectoral-fin rays; SAL: scale rows above lateral line; CEP: caudal electrocyte rows; APS: scale rows over anal-fin pterygiophores; PCV: pre-caudal vertebrae; PLR: pored lateral line scales to first ventral ramus of lateral line; PLL: total pored lateral line scales; VLR: ventral rami of lateral line. N values (in parentheses) vary because measurements were excluded from specimens with damage or unusual preservation artifacts. The mode and median are used as measures of central tendency following protocol for comparative studies of Gymnotus.
Metaphase chromosomal preparations were undertaken following the methods described by [13]. In brief: specimens were injected with a 0.025% colchicine solution in the proportion 0.5 ml/100 g body weight. After dissection, the kidney cells were suspended in 0.075 M KCl and incubated at 37°C for 30 min. The hypotonically treated cell suspension was then centrifuged and the pellet was re-suspended in fixative (3:1 methanol: acetic acid solution) and centrifuged twice. The final pellet was suspended in fresh fixative and dropped onto warmed slides. The slides were analyzed after conventional Giemsa staining, C-banding [14], silver staining of NORs [15], CMA3 staining [16], DAPI staining [17], and Fluorescente in situ Hybridization (FISH) with telomeric probes (All Telomere Probes, Oncor). Chromosomes were morphologically classified according to [18]. Fish specimens for which cytogenetic analysis was conducted were vouchered at the Museu Paraense Emilio Goeldi (MPEG) and the lot numbers are listed in Table 1.
Results
The examined specimens measured between 150 and 350 mm total length and weighed from 10 to 160 g. All had fully developed gonads and were easily sexed.
Around 20 metaphase plates for each individual specimen were examined using the procedures described above. We found two distinct karyotypes. Photographs of specimens with the two karyotypes described here are illustrated in Figure 2. At sites 1–4 (Table 1, Figure 1) all specimens presented a karyotype with a 2n (diploid number) of 42, and FN (Fundamental Number) of 72, of which 30 chromosomes were metacentric/submetacentric, and 12 subtelocentric/acrocentric (Figure 3a). At site 5 (Almeirim, Table 1, Figure 1) all specimens presented a karyotype with 2n = 40 and FN = 74, of which 34 chromosomes were metacentric/submetacentric, and 6 subtelocentric/acrocentric (Figure 4a).
Figure 2.

Specimens of: A. Gymnotus carapo from Capanema, Pará, Brazil (2n = 42) (MPEG 13331); B. Gymnotus carapo from Almeirim, Pará, Brazil (2n = 40) (MPEG 13329). Scale bar = 10 mm. Specimen A was photographed fresh, while specimen B was frozen and photographed after defrosting. This partially explains the color differences.
Figure 3.

Karyotype of Gymnotus carapo from Benfica, Pará, Brazil, with a diploid number of 2n = 42: A. Conventional Giemsa stained karyotype with the NOR pair (20); B. Sequenced C-banding; C. DAPI stained karyotype, the arrows indicate distal and interstitial markings; D. CMA3 stained metaphase (the arrows indicate the NOR pair). M-SM = Metacentric – Submetacentric; ST-A = Subtelocentric – Acrocentric.
Figure 4.

Karyotype of Gymnotus carapo from Almeirim, Pará, Brazil, with a diploid number of 2n = 40: A. Conventional Giemsa stained karyotype with the NOR pair (20); B. Sequenced C-banding; C. DAPI stained karyotype, the arrows indicate interstitial markings; D. CMA3 stained metaphase (the arrows indicate the NOR pair). M-SM = Metacentric – Submetacentric; ST-A = Subtelocentric – Acrocentric.
In both karyotypes the NOR is heteromorphic and is found in the short arm of pair 20. The 2n = 40 karyotype has an additional heterochromatin in the distal position of this pair (Figures 3a and 4a).
The C-banding in the 2n = 42 karyotype shows that the constitutive heterochromatin (CH) exhibits a centromeric location in all the chromosomes and interstitial regions on the short arms of chromosome 6 (Figure 3b). The C-banding in the 2n = 40 karyotype shows that the CH exhibits a centromeric location on almost all the chromosomes (with the exception of pairs 13, 14 and 17) and interstitial regions on the short arms of chromosome 9 (Figure 4b). One of the NOR-bearing chromosomes also possesses a block on the distal region of the short arm.
In both the 2n = 40 and 2n = 42 karyotypes, the CMA3 (Figures 3d and 4d) indicates the presence of a region rich in Guanine-Cytosine (G-C) base pairs coincident with the location of the NOR. The DAPI banding (Figures 3c and 4c) is coincident with the C-band. This demonstrates that the CH of these Gymnotus is rich in Adenine-Thymine (A-T) base pairs. FISH with telomeric probes hybridized all the telomeres. Additionally, in the 2n = 40 karyotype, there is an Interstitial Telomeric Sequence (ITS) in a metacentric pair on the short arm, close to the centromere (Figure 5).
Figure 5.
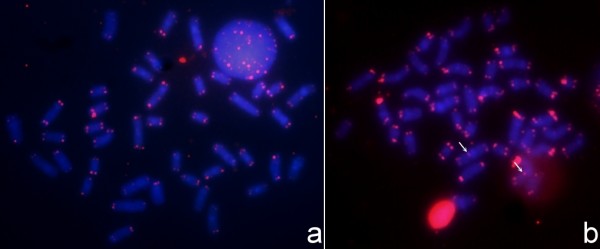
Telomeric FISH from: A. Gymnotus carapo from Benfica (2n = 42); B. Gymnotus carapo from Almeirim (2n = 40). The arrows indicate interstitial marking.
We observed no consistent differences in pigmentation or banding pattern between the animals with the two karyotypes described above (Figure 2). All exhibited the double bands with wavy indistinct margins characteristic of G. carapo senso stricto populations from the Eastern Amazon (Figure 2) (see [10], p. 10). Morphometric and meristic analysis of the two karyotypic forms indicated overlap for every measured parameter (Tables 2, 3). Principal component analysis of morphological and meristic analysis also failed to recover generalized differences between the two forms (Figure 6). Morphological measurements and meristic counts for each karyotypic form fell within the ranges published for Eastern Amazonian populations of Gymnotus carapo by [10], pp. 10–19 (Tables 2, 3). Morphological and meristic characters overlapped between the two forms. Finally, observation of the osteology of cleared and stained specimens did not reveal any obvious differences in the structure or organization of bony or cartilaginous elements. In sum, we found no evidence of morphological differentiation between the 2n = 40 and 2n = 42 karyotypic forms.
Figure 6.

Principal component scores from a correlation matrix of morphological and meristic data for two karyotypic forms of the Gymnotus carapo species complex from the Eastern Amazon. The first three principal components represent 39.9%, 67.7%, and 90.1% of cumulative variance respectively. In all combinations of these four axes the two karyotypes exhibited substantial overlap. The following meristic counts were eliminated from this analysis due to a lack of variance: P1R, BAN, SAL, APS, CEP (see Table 2 for abbreviations).
Discussion
The karyotype with the diploid number of 2n = 42 (30M/SM+12ST/A) found in G. carapo from localities 1–4 (Table 1) is the same as that previously characterized from Santa Cruz do Arari, Ilha do Marajó [9]. An almost identical karyotype, 2n = 42 (32M/SM+10ST/A), was also described [6] from G. carapo collected near Belém, Pará. The difference in morphology of one chromosomal pair may be attributed to a pericentric inversion. Alternatively, the other authors [6] may have interpreted the morphological classification of the pair differently, perhaps due to different degrees of condensation of the metaphase chromosomes. In either case, the data suggest that there is one common form of G. carapo with a homogenous 2n = 42 karyotype in the Eastern Amazon region of Belém and Marajó Island.
The 2n = 40 karyotype (28M/SM+12ST/A) from Almeirim (locality 5, Table 1) has not previously been documented from Gymnotus of the Amazon basin. Nonetheless, this diploid number has previously been documented for other species from more southerly regions of South America, but with variation in the karyotypic formula. G. sylvius (38M/SM+2ST/A [19]; 30M/SM+10ST/A, [20,21]; 36M/SM+4ST/A [22,23]; Gymnotus sp (14M/SM+26ST/A [7,8]) and G. pantanal (14M/SM+26ST/A [24]; 14M/SM+26ST/A in females and 15M/SM+24ST/A in males, with an multiple sex chromosome system X1X1X2 X2/X1X2Y [22]). The disparate phylogenetic positions of G. sylvius, G. pantanal and G. carapo senso stricto hypothesized by [1] indicates that the diploid number of 2n = 40 is homoplastic.
The location of NOR in both the 2n = 42 and 2n = 40 karyotypes were detected in the short arm of a unique pair of chromosomes, resembling the condition for karyotypes previously described in other species of Gymnotus [6,8,9,19,22,25,26].
The banding with fluorescent stains CMA3 and DAPI corroborates results previously obtained in other species of Neotropical freshwater fishes, where CMA3 preferentially stains the NOR, and where DAPI exhibits a banding model similar to that of C-banding [27].
The occurrence and apparent abundance of the same 2n = 42 form of G. carapo at multiple localities around the mouth of the Amazon: Belém [6], Santa Cruz do Arari [9] and localities 1–4 (Table 1, Figure 1), and the absence of substantial morphological variation between or within these samples suggest that they together constitute an interbreeding population of a single species. This same situation has been reported by [19] for four species of Gymnotus occurring in southeast Brazilian drainages: G. "carapo" (in fact an undescribed species that is not G. carapo) from twelve localities; G. inaequilabiatus (in fact also an undescribed species that is not G. inaequilabiatus) from two localities, G. sylvius from eight localities, and G. pantherinus from four localities. The authors of this study also observed that individuals of the same species always possessed the same karyotype in different localities, for example, 2n = 54, 52M/SM+2ST/A in G. "carapo".
The difference in the diploid number (2n = 40 and 2n = 42) reported here can be explained by a fusion-fission rearrangement. The presence of an ITS in a metacentric pair in the 2n = 40 karyotype (Figure 5b) supports the fusion hypothesis, where two ST/A pairs from the 2n = 42 karyotype fused originating a M/SM pair in the 2n = 40 karyotype. The morphologic variation in the karyotypic formula (2n = 40: 34M/SM+6ST/A e 2n = 42: 30M/SM+12ST/A) can be explained also by pericentric inversions. According to [28], multiple pericentric inversions are an important post-zygotic reproduction isolation mechanism. The absence of morphologic differentiation suggests that the putative chromosomal speciation event occurred recently and that there was consequently insufficient time for the fixation of phenotypic differences. These considerations lead us to speculate that the Almeirim sample may be a cryptic species that is isolated by post-zygotic reproductive barriers from an extant sister taxon (the 2n = 42 form), and from which it derived by chromosomal rearrangement. This hypothesis is amenable to empirical test with molecular phylogenetic data and by examination of EOD signal variation.
A bibliographical survey of cytogenetic data for taxa assigned to G. carapo indicates a large amount of chromosomal variability (Figure 1 and Table 4) through Brazil. For instance a sample from Humaitá, in the Rio Madeira basin of Amazonas State exhibited a diploid number of 2n = 48 [6]. Several samples from the Brazilian state of São Paulo (at Jundiaí, Rio Claro, Americana, Botucatu, Paula Souza, Salto Grande, Primeiro de Maio, Pirassununga, Mococa, São Simão, Santa Maria da Serra and Jacareí) presented diploid numbers varying from 2n = 52 to 54 [6,19,21,26]. As discussed earlier, G. carapo senso stricto [10] and also closely related G. carapo-like forms from further south probably comprises a complex of morphologically similar or cryptic species (Crampton and Albert, pers. obs.), with species-level variation in karyotypes. The existence of a cryptic species of Gymnotus carapo (2n = 40) from Almeirim, in a region otherwise dominated by a single 2n = 42 taxon, is concordant with this emerging notion.
Table 4.
Localities and diploid number for karyotpes of Gymnotus species in Brazil (see also Figure 1).
| Species | Localities and 2n |
| G. carapo | Miracatu-SP [6], Botucatu-SP [6], Jundiaí-SP [19], Rio Claro-SP [19], Americana-SP [19], Salto Grande-SP [19], Primeiro de Maio-SP [19], Mococa-SP [19], São Simão-SP [19], Santa Maria da Serra-SP [19], Jacareí-SP [19] (2n = 54); Pirassununga-SP [26] (2n = 54 e 2n = 81); Brotas-SP [6], (2n = 52); Humaitá-AM [6] (2n = 48); Belém-PA [6], Benfica-PA1, São Miguel do Guamá-PA1, Capanema-PA1, Ponta de Pedras-PA1, Santa Cruz do Arari-PA [9], (2n = 42); Almeirim-PA1 (2n = 40). |
| G. sp. | Miracatu-SP [6] (2n = 52), Corrientes, Argentina [7] and Guaíra-PR [8], (2n = 39–40 X1X2Y), Alfenas-MG [23], (2n = 50). |
| G. sylvius | Miracatu-SP [20], São Simão-SP [19,20], Americana-SP [19], Represa de Capivara-SP [19], Sta. Maria da Serra-SP [19], Corumbataí-SP [19], Jacareí-SP [19], Paraibuna-SP [19] (2n = 40), Guaíra-PR [22], Alfenas-MG [23], (2n = 40). |
| G. inaequilabiatus | Rio Claro-SP [19], Represa de Porto Primavera-SP [19] (2n = 52). |
| G. pantherinus | Paranapiacaba-SP [32], Itanhaém-SP [32], Serra da Juréia-SP [32], (2n = 52). |
| G. pantanal | Mato Grosso do Sul-MS [24], (2n = 40), Guaíra-PR [22], (2n = 39–40 X1X2Y). |
| G. mamiraua | Sta. Cruz do Arari-PA [9], (2n = 54). |
| G. paraguensis | Guaíra-PR [22]; Alfenas-MG [23], (2n = 54). |
Brazilian state abbreviation are: AM = Amazonas; MS = Matogrosso do Sul; PA = Pará; PR = Paraná; SP = São Paulo; N = Sample; NI = Not informed; [6], (N = 16); [26], (N = 17); [19], (N = 166); [9], (N = 5); [7], (N = 20); [8], (2005) (N = 20); [20], (N = NI); [32], (N = NI); [24], (N = NI); [22], (N = 54); [23],12- Lacerda & Maistro (2007) (N = 27); 1- Present work (N = 17);
We expect that the wide geographic range, diversity of species, and cytogenetic diversity of the Gymnotus carapo species-complex will provide a good testing ground for exploring the role of chromosomal evolution in speciation. A holistic approach involving cytogenetics, phylogenetic systematics, population genetics, and the study of electric communication signals (putative pre-zygotic reproductive isolating barriers, see [29-31]) may allow us to trace individual speciation events (including those separating extant sister species) to documented chromosomal rearrangement events, such as the one we hypothesize here.
Conclusion
The difference in the diploid number and FN between a 2n = 42 form of Gymnotus from the Eastern Amazon, and a morphologically identical form with 2n = 40 can be explained by a fusion/fission and pericentric inversions rearrangements. We speculate that these two forms are cryptic species, isolated from each other by post-zygotic reproductive isolating barriers.
Authors' contributions
SSRM collected the samples, collaborated in all the cytogenetic procedures, undertook the bibliographic review, and coordinated the writing of this paper. JCP helped conceive the study, and participated in the development of laboratory techniques, cytogenetic analyses and writing. WGRC undertook morphological analyses, and wrote the taxonomic sections. DSS participated in collecting, and in the development of laboratory techniques. ACPS and JRC helped with collecting. CYN coordinated the study, helped in the development of laboratory techniques, cytogenetic analyses and reviewed the manuscript. All authors read and approved the final manuscript.
Acknowledgments
Acknowledgements
We acknowledge the Museu Paraense Emilio Goeldi (MPEG) for vouchering specimens. We thank Geoffrey Shetka for assistance with the morphological measurements (funded by NSF Research Experience for Undergraduates internship, DEB-0724280 to W. Crampton). Funds were supported by CNPq, CAPES, UFPA. W. Crampton received funding from National Science Foundation grant DEB-0614334 (Evolution of Species and Signal Diversity in the Neotropical Electric Fish Gymnotus). Collecting was authorized by IBAMA (Instituto Brasileiro do Meio Ambiente) permit 020/2005 (IBAMA Registration: 207419).
Contributor Information
Susana SR Milhomem, Email: susanamilhomem@yahoo.com.br.
Julio C Pieczarka, Email: juliopieczarka@pesquisador.cnpq.br.
William GR Crampton, Email: crampton@mail.ucf.edu.
Danillo S Silva, Email: dssufpa@yahoo.com.br.
Augusto CP De Souza, Email: augustoacari@yahoo.com.br.
Jaime R Carvalho, Jr, Email: jaimejr@ufpa.br.
Cleusa Y Nagamachi, Email: cleusanagamachi@pq.cnpq.br.
References
- Albert JS, Crampton WGR, Thorsen DT, Lovejoy NR. Phylogenetic systematics and historical biogeography of the Neotropical electric fish Gymnotus (Teleostei: Gymnotiformes) Systematics and Biodiversity. 2005;2:375–417. [Google Scholar]
- Albert JS, Crampton WGR. Diversity and phylogeny of Neotropical electric fishes (Gymnotiformes) In: Bullock TE, Hopkins CD, Popper AN, Fay FR, editor. Electroreception. New York: Springer; 2005. pp. 360–403. [Google Scholar]
- Crampton WGR, Albert JS. Evolution of electric signal diversity in Gymnotiform fishes. I. Phylogenetic systematics, ecology and biogeography. In: Ladich F, Collin SP, Moller P, Kapoor BG, editor. Communication in Fishes. Enfield-NH: Science Publishers; 2006. pp. 647–696. [Google Scholar]
- Cognato D, Richer-de-Forges MM, Albert JS, Crampton WGR. Gymnotus chimarrao : a new species of Neotropical electric fish (Gymnotiformes: Gymnotidae) from Southern Brazil. Ichthyol Exploration in Freshwaters. 2007;18:375–382. [Google Scholar]
- Caputi AA, Carlson BA, Macadar O. Electric organs and their control. In: Bullock TE, Hopkins CD, Popper AN, Fay FR, editor. Electroreception. New York: Springer; 2005. pp. 410–452. [Google Scholar]
- Foresti F, Almeida-Toledo LF, Toledo-Filho SA. Chromosome studies in Gymnotus carapo and Gymnotus sp. (Pisces, Gymnotidae) Caryologia. 1984;37:141–146. [Google Scholar]
- Sánchez S, Laudicina A, Jorge LC. A New Report of Multiple Sex Chromosome System in the Order Gymnotiformes (Pisces) Cytologia. 2004;69:155–160. [Google Scholar]
- Silva EB, Margarido VP. An X1X1X2X2/X1X2Y multiple sex chromosome system in a new species of the genus Gymnotus (Pisces, Gymnotiformes) Environmental Biology of Fishes. 2005;73:293–297. [Google Scholar]
- Milhomem SSR, Pieczarka JC, Crampton WGR, de Souza ACP, Carvalho JR, Nagamachi CY. Differences in karyotype between two sympatric species of Gymnotus (Gymnotiformes: Gymnotidae) from the Eastern Amazon of Brazil. Zootaxa. 2007;1397:55–62. [Google Scholar]
- Albert JS, Crampton WGR. Seven new species of the Neotropical electric fish Gymnotus (Teleostei: Gymnotiformes) with a redescription of G. carapo (Linnaeus) Zootaxa. 2003;28:1–54. [Google Scholar]
- Albert JS, Crampton WGR. Five new species of Gymnotus (Gymnotiformes, Teleostei) from an Upper Amazon floodplain, with descriptions of electric organ discharges and ecology. Ichthyol Exploration in Freshwaters. 2001;12:241–266. [Google Scholar]
- Crampton WGR, Thorsen DH, Albert JS. Three new species from a diverse and sympatric assemblage of the electric fish Gymnotus (Ostariophysi: Gymnotidae) in the lowland Amazon Basin, with notes on ecology. Copeia. 2005;1:82–99. [Google Scholar]
- Bertollo LAC, Takashi CS, Moreira-Filho O. Citotaxonomic considerations on Hoplias lacerdae (Pisces, Erythrinidae) Brazilian Journal of Genetics. 1978;2:103–120. [Google Scholar]
- Sumner AT. simple technique for demonstrating centromeric heterochromatin. Experimental Cell Research. 1972;75:304–306. doi: 10.1016/0014-4827(72)90558-7. [DOI] [PubMed] [Google Scholar]
- Howell WM, Black DA. Controlled silver-staining of nucleolus organizer regions with a protective colloidal developer: a 1-step method. Experientia. 1980;36:1014–1015. doi: 10.1007/BF01953855. [DOI] [PubMed] [Google Scholar]
- Schweizer D. Simultaneous fluorescent staining of R bands and specific heterochromatic regions (DA/DAPI Bands) in human chromosomes. Cytogenetics and Cell Genetics. 1980;27:190–193. doi: 10.1159/000131482. [DOI] [PubMed] [Google Scholar]
- Pieczarka JC, Nagamachi CY, Souza ACP, Milhomem SSR, Castro RR, Nascimento AL. An adaptation to DAPI-banding to fishes chromosomes. Caryologia. 2006;59:43–46. [Google Scholar]
- Guerra MS. Reviewing the chromosome nomenclature of Levan et al. Brazilian Journal of Genetics. 1986;9:741–743. [Google Scholar]
- Fernandes-Matioli FMC, Marchetto MCN, Almeida-Toledo LF, Toledo-Filho SA. High intraspecific karyological conservation in four species of Gymnotus (Pisces: Gymnotiformes) from Southeastern Brazilian basins. Caryologia. 1998;51:221–234. [Google Scholar]
- Albert JS, Fernandes-Matiole FMC, Almeida-Toledo LF. New species of Gymnotus (Gymnotiformes-Teleostei) from southeastern Brazil: Toward the deconstruction of Gymnotus carapo. Copeia. 1999;2:410–421. [Google Scholar]
- Fernandes-Matioli FMC, Almeida-Toledo LF. A Molecular phylogenetic analysis in Gymnotus species (Pisces: Gymnotiformes) with inferences on chromosome evolution. Caryologia. 2001;54:23–30. [Google Scholar]
- Margarido VP, Bellafronte E, Moreira-Filho O. Cytogenetic analysis of three sympatric Gymnotus (Gymnotiformes, Gymnotidae) species verifies invasive species in the Upper Paraná River basin, Brazil. Journal of Fish Biology. 2007;70:155–164. [Google Scholar]
- Lacerda MCV, Maistro EL. Cytogenetic Analysis of Three Sympatric Gymnotus Species (Teleostei: Gymnotidae) from the Fundo Stream, MG, Brazil. Cytologia. 2007;72:89–93. [Google Scholar]
- Fernandes FMC, Albert JS, Daniel-Silva MFZ, Lopes CE, Crampton WGR, Almeida-Toledo LF. A New Gymnotus (Teleostei: Gymnotiformes: Gymnotidae) from the Pantanal Matogrossense of Brazil and adjacent drainages: continued documentation of a cryptic fauna. Zootaxa. 2005;933:1–14. [Google Scholar]
- Fernandes-Matioli FMC, Almeida-Toledo LF, Toledo-Filho SA. Extensive nucleolus organizer region polymorphism in Gymnotus carapo (Gymnotoidei, Gymnotidae) Cytogenet Cell Genet. 1997;78:236–239. doi: 10.1159/000134665. [DOI] [PubMed] [Google Scholar]
- Fernandes-Matioli FMC, Almeida-Toledo LF, Toledo-Filho SA. Natural triploidy in the Neotropical species Gymnotus carapo (Pisces: Gymnotiformes) Caryologia. 1998;51:319–322. [Google Scholar]
- Nascimento AL, Souza ACP, Feldberg E, Carvalho JR, Barros RMS, Pieczarka JC, Nagamachi CY. Cytogenetic analysis on Pterophyllum scalare (Perciformes, Cichlidae) from Jari River, Pará state. Caryologia. 2006;59:138–143. [Google Scholar]
- King M. Species Evolution: The role of chromosome change. Cambridge: University Pres; 1993. [Google Scholar]
- Crampton WGR. Evolution of electric signal diversity in gymnotiform fishes. II. Signal design. In: Ladich F, Collin SP, Moller P, Kapoor BG, editor. Communication in Fishes. Enfield-NH: Science Publishers; 2006. pp. 697–731. [Google Scholar]
- Rodriguez-Cattaneo A, Pereira AC, Aguilera PA, Crampton WGR, Caputi AA. Species-specific diversity of a fixed motor pattern: the electric organ discharge of Gymnotus. PLoS ONE. 2008;3:e2038. doi: 10.1371/journal.pone.0002038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crampton WGR, Davis JK, Lovejoy NR, Pensky M. Multivariate classification of animal communication signals: A simulation-based comparison of alternative signal processing procedures using electric fishes. J Physiol Paris doi: 10.1016/j.jphysparis.2008.10.001. [DOI] [PubMed] [Google Scholar]
- Marchetto MC, Fernandes-Matioli FM, Almeida-Toledo LF. Estudos citogenéticos comparativos em populações de Gymnotus pantherinus (Pisces, Gymnotidae) Genetic and Molecular Biology. 1998;21:65. [Google Scholar]