

Abstract
The pathogenesis of autoimmune hepatitis (AIH) is complex. However, it is believed that a susceptible individual, owing to his genetic background, sex and age, can develop the disease following exposure to an environmental trigger. Autoimmune hepatitis does not follow a Mendelian pattern of inheritance; hence no single causative genetic locus has been identified. However, several genes, inside and outside the HLA locus, have been linked to an increased susceptibility to AIH. Epidemiological evidence also suggests that the sex and age of the patient plays a role in AIH pathogenesis as the disease onset occurs mainly in the two first decades of life and a higher disease incidence is observed in females. No environmental trigger has been identified, but several have been proposed, mainly viruses and xenobiotics. This article aims at reviewing the current knowledge on susceptibility factors leading to AIH and putative triggers, emphasizing fundamental mechanisms responsible for the break of liver immunological tolerance.
Keywords: Autoimmune hepatitis, Genetic, Enviro-nment, Sex, Virus
INTRODUCTION
Autoimmune hepatitis (AIH) was described more than 50 years ago by Jan Gösta Waldenström and Henry George Kunkel[1] and patients were referred to as “Kunkel-Waldenström girls”. In 1959, following the observation by Ian Mackay of lupus erythematosus (LE) cells in AIH patients, the disease was referred to as “lupoid hepatitis” as it was believed to be a form of lupus erythematosus[1]. As knowledge of symptoms, natural course of disease, pathogenesis and treatment progressed, the name “chronic active hepatitis” was chosen and finally “autoimmune hepatitis” was adopted at the first meeting of the International Autoimmune Hepatitis Group.
It is now believed that this autoimmune disease results from the progressive destruction of the hepatic parenchyma through a loss of immune tolerance towards hepatocytes. While the origin of the immune system dysregulation is still unknown, recent fundamental and clinical research have shed some light on predisposing factors and immune mechanisms involved. The current hypothesis for AIH pathogenesis is that this immune dysregulation is a consequence of an environmental triggering event in a genetically predisposed individual of a particular sex and age (Figure 1).
Figure 1.
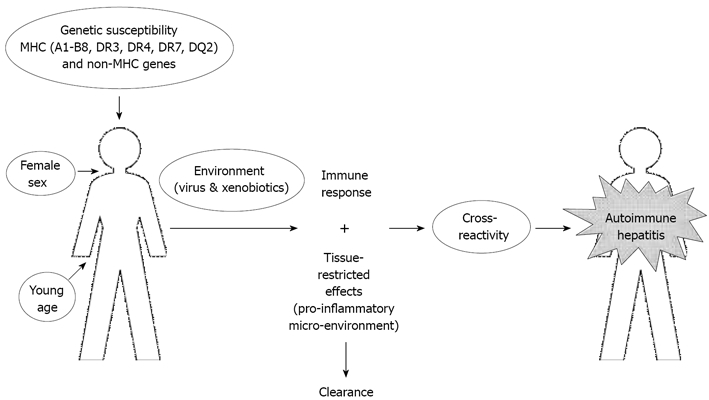
Pathogenesis of autoimmune hepatitis (molecular mimicry hypothesis). AIH can occur in an individual of a particular sex and age with a genetic background of susceptibility. To develop AIH, this individual must encounter an environmental trigger such as an infection or exposure to a xenobiotic. This will induce an immune response and secretion of pro-inflammatory molecules resulting in the elimination of the pathogen. However, this response may also lead to an immune cross-reactivity with liver proteins with an efficient activation of autoreactive cells leading to a break of immunological tolerance towards the liver.
AUTOIMMUNE HEPATITIS: AN OVERVIEW
Autoimmune hepatitis is a disease with a chronic, but fluctuating course. Although primarily a female pediatric disease, autoimmune hepatitis is not limited to patients of a particular sex, age or ethnic group. Epidemiologic studies estimate the prevalence of AIH to be between 50 to 200 cases per million in Caucasian populations of Europe and North-America[2,3]. It is characterized by hypergammaglobulinemia, circulating autoantibodies, low levels of complement factor 4a (C4A) and a high prevalence of a HLA B8, DR3 and DR4 haplotype[4]. AIH clinical presentation can vary, from an acute to chronic hepatitis. Nonspecific symptoms include fatigue, anorexia and weight loss. Most AIH patients respond well to immunosuppressive treatments. The diagnosis of AIH is made according to a scoring system established by the International Autoimmune Hepatitis Group[4,5] which encompasses several clinical parameters and predisposing factors.
Liver histology in autoimmune hepatitis patients is usually characterized by portal and periportal inflammation (interface hepatitis) and lobular hepatitis. Approximately 50% of biopsies show some degree of bridging necrosis[4]. Infiltrates are mainly composed of a mononuclear cells with an abundance of plasma cells. Ten to twenty percents of liver biopsies also show multinucleated giant hepatocytes[4]. At diagnosis, portal fibrosis can be present ranging from an enlargement of the portal tract to cirrhosis.
Autoimmune hepatitis has been classified into two types according to circulating autoantibodies present in patients’ sera. Type 1 AIH is characterized by the presence of anti-nuclear antibodies (ANA) and/or anti-smooth muscle antibodies (SMA) and/or anti-soluble liver antigen (SLA)[6,7]. Type 2 AIH patients are defined by the presence of circulating anti-liver-kidney microsome antibodies (LKM1) and/or anti-liver cytosol 1 antibodies (LC1)[8,9]. Anti-LKM1 antibodies recognize the cytochrome P4502D6 (CYP2D6)[10] and anti-LC1 antibodies react against the formiminotransferase cyclodeaminase (FTCD)[9]. Both CYP2D6 and FTCD are mainly expressed by hepatocytes. The incidence ratio of type 1 to type 2 AIH is 1.5 to 2:1 in Europe and reaches 7:1 in North and South America and Japan[11]. Several clinical differences exist between type 1 and 2 AIH (Table 1). The mean age of onset is 10 years in type 1 AIH and 6.5 years in type 2 AIH, and the female to male ratio is higher in type 2 AIH[6,12]. Type 2 AIH patients frequently present a more severe disease course and are more likely to be resistant to treatment[13]. In addition, associated extrahepatic autoimmune diseases are different in type 1 and 2 AIH. For example, type 1 AIH is more frequently associated with inflammatory bowel diseases and sclerosing cholangitis, diseases that are never observed in type 2 AIH[6,12].
Table 1.
Clinical and biochemical characteristics of type 1 and type 2 autoimmune hepatitis
| Characteristics | |
| Type 1 AIH | Anti-SMA, anti-ANA and/or anti-SLA |
| Mean age of onset: 10 yr old | |
| Female:Male ratio 3:1 | |
| Higher incidence | |
| Frequently associated with IBD and sclerosing cholangitis | |
| Type 2 AIH | Anti-LKM1 and/or anti-LC1 |
| Mean age of onset: 6.5 yr old | |
| Female:Male ratio 9:1 | |
| Associated with HLA DR3, DR7 and DQB1*0201 | |
| More severe disease | |
| More likely to be resistant to treatment |
GENETIC SUSCEPTIBILITY
Autoimmune hepatitis does not follow a Mendelian pattern of inheritance and no single genetic locus has been identified as responsible for the disease. It is generally believed that one or more genes, acting alone or in concert, reduce or increase susceptibility to AIH.
The strongest association between genes and autoimmune hepatitis has been found at the human leukocyte antigen (HLA) locus on chromosome 6. Susceptibility alleles have been identified in several populations (Table 2). In North America and Europe, HLA-A1-B8, HLA-DRB1*0301 and HLA-DRB1*0401 (DR3 & DR4) have been associated with a susceptibility to AIH[14,15]. Through a linkage disequilibrium study in families of AIH patients, HLA-DRB1*03 (DR3) and DRB1*1301 (DR13) as well as HLA-DQB1*0201 were found to be preferentially transmitted to patients compared to unaffected siblings in type 1 and type 2 AIH, respectively[16]. Another genetic study proposed that HLA-DR13 could be a risk factor in the absence of HLA-DR3 or HLA-DR4[17]. However, the size of the population studied did not allow reaching statistically significant conclusions. Hence this association needs confirmation.
Table 2.
Specific susceptibility genes in type 1 and type 2 autoimmune hepatitis
| Genes | Population | Type of AIH | Linkage disequilibrium | References |
| MHC genes | ||||
| HLA-A1-B8 | North-America, Europe | 1 | Yes | [14,15] |
| HLA-DRB1*0301 | North-America, UK, Spain, Argentina | 1 and 2 | Yes | [14,15,25] |
| HLA-DRB1*0401 | North-America, Europe | ND | [14,15] | |
| HLA-DRB1*0404 | Mexico | 1 | ND | [18] |
| HLA-DRB1*0405 | Argentina, Japan | 1 | - | [19,20] |
| HLA-DRB1*1301 | North-America, Europe, Brazil, Argentina | 1 | Yes | [16,21] |
| HLA-DRB1-07 | Germany, Brazil, UK | 2 | Yes | [23,24] |
| HLA-DRB3*01 | Brazil | [22,23] | ||
| HLA-DQB1*0201 | North-America, Europe | 2 | Yes | [16] |
| HLA-DQB1*0603 | North-America, Europe | 2 | Yes | [16] |
| Non-MHC genes | ||||
| IgA | Europe | 1 | [28,29] | |
| C4A | Europe, North-America | 1 and 2 | Yes | [30,31] |
| CTLA4 | North-America, Europe | 1 | Yes | [35,36] |
| Fas | Japan, North-America | 1 | ND | [37,38] |
| Vitamin D receptor | Germany | 1 and 2 | ND | [39] |
| TNFa*2 | North-America, UK | 1 | Yes | [43,44] |
ND: Non-determined.
Other HLA alleles have also been described as risk factors for autoimmune hepatitis in other populations (Table 2). Among Mestizo Mexicans, HLA-DRB1*0404 is predominant in adult AIH patients[18]. In Japan and Argentina, HLA-DRB1*0405 has been associated with AIH[19,20] while in Brazil, HLA-DRB1*1301 and DRB3*01 are associated with the disease[19,21,22]. In type 2 autoimmune hepatitis, HLA-DRB1*07 has been associated in German, Brazilian and British populations while HLA-DRB1*03 was found as a risk factor in Spanish patients[23–25]. These differences in susceptibility alleles among various ethnic groups could be explained by the shared motif hypothesis which proposes that multiple alleles can encode for similar motifs within HLA class II. In 94% of type 1 AIH patients, susceptibility alleles encode the LLEQKR or LLEQRR motifs at position 67-72 of class II HLA[15,26]. In contrast, HLA-DB1*1501, which is associated with a reduced risk to develop type 1 AIH, encodes for the ILEQAR motif[15,26]. Substitution of a lysine or arginine to alanine at position 71, which changes both polarity and charge, possibly modifying peptide binding and orientation in the MHC, could influence autoantigen presentation to T cell receptors (TCR).
HLA alleles have also been found to influence the autoantigenic humoral response. In a recent study, HLA-DQB1*0201 was described as the main allele in association with susceptibility to type 2 AIH[27]. DQ2 is in linkage disequilibrium with DR3 or DR7, both associated with type 2 AIH. Interestingly, HLA-DRB1*03 was found associated with type 2 AIH patients which show both LKM1 and LC1 antibodies in their sera, while HLA-DRB1*07 was predominant amongst type 2 AIH patients, whose sole serological marker was anti-LKM1[27]. In addition, children carrying the HLA-DRB1*07 allele developed a more restricted repertoire of anti-LKM1 epitopes compared to those carrying the HLA-DRB1*03 allele[27].
Other genes located at the HLA locus are linked with AIH susceptibility, such as the IgA and complement factor 4A genes[12]. IgA deficiency is common in AIH patients. This deficiency is genetically linked to the MHC locus, especially with HLA susceptibility alleles such as HLA-DR1 and HLA-DR7[28,29]. Also, low levels of C4a are found in 69% of children with AIH[30]. Complement factor 4a (C4a) has also been linked with AIH pathogenesis since deletions in the C4A gene were found in patients who develop AIH at a younger age[31].
Genes outside the HLA locus have also been linked with AIH using single nucleotide polymorphism (SNP) screening techniques. These genes encode proteins which influence either the innate or adaptive immune system. As in Graves’ disease[32], multiple sclerosis[33] and coeliac disease[34], cytotoxic T-lymphocytes antigen 4 (CTLA-4) gene polymorphisms have been found in adult and children with type 1 AIH[35,36]. A linkage disequilibrium was also found in affected children compared to non-affected siblings[36]. A FAS gene promoter polymorphism (position -670) was found to influence susceptibility to AIH[37] and its progression, leading to a more aggressive disease with an early development of cirrhosis[38]. Recently, polymorphisms in the vitamin D receptor was shown to contribute to development of autoimmune liver diseases[39]. This receptor was found to have immunomodulatory functions such as macrophage and monocyte activation, inhibition of Th1 functions and prevention of dendritic cells differentiation[40–42]. Therefore, polymorphisms in the vitamin D receptor could influence the immune response towards autoantigens. Polymorphisms in the tumor necrosis factor-α (TNF-α) gene (TNFA*2) confers a susceptibility to AIH and influences the natural course of the disease. A G to A substitution at position -308 is believed to influence gene transcription and result in higher induced or constitutive levels of circulating TNF-α[43,44]. AIH patients who possess this polymorphism are prone to early disease development, are less likely to enter into remission and more prone to develop liver cirrhosis[44].
Mutations in the autoimmune regulator gene (AIRE) responsible for the development of Autoimmune Polyendocrinopathy-Candidiasis-Ectodermal Dystrophy (APECED) in patients can also lead to AIH in 10% to 20% of cases[45]. The AIRE gene encodes for a transcription factor involved in the thymic negative selection of lymphocytes. Thus, mutations that impair this function could cause multiple autoimmune manifestations. However, studies on known AIRE mutations in patients with autoimmune liver diseases showed that the AIRE gene does not play a major role in their pathogenesis[32,46].
Genetic background influence on the development of an AIH has also been observed in an animal model of type 2 AIH[47]. Xenoimmunisation with plasmid DNA coding for type 2 human autoantigens was performed in three mouse strains which differ in their MHC and/or non-MHC genes[48]. C57BL/6 mice developed a severe AIH while 129/Sv mice, who share the same MHC alleles as with C57BL/6, but on a different non-MHC genetic background, had a mild AIH. In contrast, BALB/c mice, which differ in both MHC and non-MHC genetic background, did not develop AIH. These results highlight the importance of both MHC and non-MHC genes in the initiation and progression of an autoimmune response towards the liver after an environmental triggering event, xenoimmunisation in this case, occurs[48].
INFLUENCE OF SEX AND AGE ON AUTOIMMUNITY
Most autoimmune diseases show a striking sex difference in their incidence, women being affected more frequently than men[49]. Differences in incidence between women and men range from 20:1 in Sjogren’s syndrome to 3:2 in multiple sclerosis[49]. Less frequently, the female to male ratio approaches 1:1, as in ulcerative colitis and diabetes[49].
In AIH, the female to male ratio ranges from 3:1, in type 1, to 9:1, in type 2 AIH[11]. Sex differences in the immune response are also observed in other liver diseases. For example, men are more likely to become chronic carriers of hepatitis B than women[50]. In addition, several studies have investigated the effect of donor:recipient sex matching in the outcome of orthotopic liver transplantation (OLT) and have found that male to female grafts have the most, and female to male the least favourable, outcome, both in terms of patient and graft survival[51].
This gender discrepancy could be the result of existing differences in basic immune responses between females and males. In fact, higher levels of antibodies and stronger T cell activation are observed in women after vaccination[52]. Women have higher absolute numbers of CD4+ T-cells and produce higher levels of Th1 cytokines than men[53]. Interestingly, in vitro oestrogen increases Th1 cytokine production by T lymphocytes, while a decrease is observed in presence of androgen[54].
Age also influences the incidence of several autoimmune diseases suggesting the role of hormones in the pathogenesis of these diseases. AIH is primarily a pediatric disease; 40% of type 1 and 80% of type 2 AIH cases are diagnosed before the age of 18[6,12]. A second peak of incidence of AIH has also been reported in women after menopause[55]. The hormonal status of patients could be related to these prepubertal and post-menopausal peaks of incidence. In fact, sexual hormones are known to directly modulate immune responses and, by doing so, alter the development of autoimmune diseases. 17β-estradiol has been shown to suppress IL-2 secretion by T cells and inhibit IL-2 receptor expression in activated peripheral blood T cells[56]. In vivo, 17β-estradiol (E2) protects C57BL/6 mice from experimental autoimmune encephalomyelitis (EAE)[57]. However, 17β-estradiol was also shown to enhance susceptibility to experimental myasthenia gravis[58], experimental autoimmune uveoretinitis[59] and lupus[60]. Therefore, the effects mediated by estradiol on autoimmunity are diverse and not fully understood. Male sex hormones also affect immune responses. Testosterone can directly affect CD4+ T cells via androgen receptors and induce increased secretion of IL-10, an anti-inflammatory cytokine[61]. Testosterone was also found to protect female SJL mice from developing EAE[62].
Hormonal status of AIH patients during pregnancy can also impact the disease course, with both improvement and exacerbations reported[63,64]. Patients who experience a remission of their disease during pregnancy generally have a disease flare-up after delivery[64]. In some cases, AIH is diagnosed in the first few months of pregnancy or post-partum[65].
Currently, no pathological mechanism and/or direct hormonal effect can explain these observations. Although epidemiological studies show the impact of sex and age on AIH, more research will be needed to understand the interaction of sex, age and autoimmunity.
ENVIRONMENTAL FACTORS
Environmental factors are thought to be the triggerering event for the development of an AIH in genetically predisposed individuals of a particular sex and age. These environmental factors could be drugs, chemicals or viruses. They are believed to initiate the autoimmune response through several means: (1) non-specific activation of resting T cells; (2) modification or release of sequestered proteins; (3) cross-reactivity between virus and self-protein (molecular mimicry); and (4) modulation of gene expression.
Non-specific activation of resting T cells
Non-specific activation of resting T cells has been reported after various virus infections, e.g. Epstein-Barr virus (EBV). It could be speculated that resting autoimmune T cells become activated and proliferate leading to an AIH development. EBV infection preceding the onset of AIH has been reported in some patients[66–69]. While this mechanism could be involved in AIH development, more evidence is needed to confirm its role in AIH pathogenesis.
Xenobiotics could also be a non-specific activator of lymphocytes, as observed in a murine model of immune-mediated hepatic injury induced by injections of Concanavalin A (ConA)[70]. Concanavalin A is a leptin that stimulates the release of various cytokines by lymphocytes, mainly INF-γ and TNF-α[71]. It can also directly stimulate T cells by binding the of MHC and induction of their proliferation[72]. This massive non-specific T cell activation results in hepatitis through a bystander effect mediated by INF-γ and TNF-α[70,71]. Although this murine model does not rely on an autoimmune reaction against the liver per se, it has allowed a better understanding of how xenobiotics could lead to a T cell dependent autoimmune disease.
Modification or release of sequestered protein
An ever growing list of drugs and chemicals has been linked with AIH development in humans. Among these, minocycline, a drug used to treat acne, has been frequently associated with liver autoimmunity[73–75]. Interestingly, when minocycline treatment is stopped, the AIH-like syndrome disappears. Herbal agents such as black cohosh[76], a herbal medicine used to treat menopausal symptoms, and dai-saiko-to[77], a herbal medicine used in Japan, have also been proposed as causative agents for AIH. Recently, a case report of 3 adults and meta-analysis of previous case reports has associated atorvastatin and simvastatin with AIH[78]. This report is significant since statins are amongst the most widely prescribed drugs. However, these patients were also genetically predisposed for AIH, being HLA-DR3, DR4 or DR7[78]. No mechanisms have been proposed to explain the autoimmune effects of these drugs and chemicals. Explanations may lie in the hepatotoxic effect of these chemicals, which could release autoantigens, up-regulate proteins expression (P450s, immunoregulatory proteins) or act as a hapten by modifying the hepatic protein, making them immunogenic.
In experimental models, potential mechanisms for xenobiotics resulting in an immune-mediated liver disease have been described. A model of primary biliary cirrhosis (PBC) in guinea pigs was developed by the injection of 6-bromohexoanate to mimic the lipoate moiety of PDC-E2, the main epitope recognized by anti-mitochondrial antibodies from sera of patients with PBC[79]. In this model, the disease becomes evident 18 mo after being exposed to 6-bromohexoanate. This may suggest that exposition to a xenobiotic and induction of a clinically apparent disease could be a long-term process, and should be taken into consideration in future cause-effect studies on xenobiotic exposition and AIH development.
Another mechanism which could explain the development of an AIH is that a hepatotropic viral infection could result in a release of sequestered autoantigens from hepatocytes within a pro-inflammatory environment. This would lead to an autoimmune reactivity towards hepatic antigens. Also, since hepatocytes can express the MHC class II molecule during the course of a clinical hepatitis[80], a function normally reserved to antigen presenting cells (APC), they acquire the ability to specifically activate CD4+ T cells and induce an immune response[80]. This hypothetic mechanism implies that hepatotropic viruses may trigger an autoimmune reactivity in a non-specific manner by giving activated immune cells access to autoantigens in a pro-inflammatory environment.
However, the liver is involved in the development of immunological tolerance towards oral antigens[81] and thus, has a tolerogenic immune environment resulting in poor immune cell activation and response. Experimental evidence shows that CD4+ T cells activated by hepatocytes are more likely to become CD4+Th2 cells and impair the CD8+ T cell response[82]. Some studies suggest that efficient immune responses are difficult to elaborate in the liver compared to lymph nodes[83]. Furthermore, work from Bowen et al[84] showed that specific T cells directed against a liver antigen are not properly activated if this antigen is uniquely expressed in the liver. A break of tolerance towards liver antigens could only be observed when the liver antigen was also expressed in the periphery[47,84,85]. In light of these results, both hepatotropic and non-hepatotropic viruses should be considered as potential triggering events leading to the development of an AIH, through release of autoantigens in a pro-inflammatory environment and/or by molecular mimicry between viruses and autoantigens.
Molecular mimicry
In many autoimmune disorders, molecular mimicry between a virus and a self-protein has been hypothesized to be the key event leading to the disease. Molecular mimicry occurs when a virus protein sequence, structure or motif is shared with a self-protein. The immune system will mount a response against the virus but, in the process, will cross-react with a homologous self-protein. This immune cross-reactivity could evolve, under certain circumstances, into an autoimmune disease. This hypothesis has been proposed in several autoimmune disorders such as multiple sclerosis where homologies between several infectious agents and the myelin basic protein were found[86].
In two animal models, molecular mimicry was proven to be a possible triggering mechanism for AIH. In the TTR-nucleoprotein (NP) transgenic mouse, which expresses the lymphocytic choriomeningitis virus (LCMV) NP under the control of a liver-specific promoter, DNA vaccination with a plasmid coding for the LCMV-NP led to a liver-specific immune response and a progressive destruction of the hepatic parenchyma[85]. In this case, a molecular identity between the self-protein and the injected antigen was the triggering factor for AIH development. In a model of type 2 AIH, DNA-vaccination of wild-type C57BL/6 mice with a plasmid coding for human type 2 autoantigens, CYP2D6 and FTCD, led to a break of immune tolerance towards the murine homologues of these proteins (the CYP2D9 and murine FTCD). While the initial immune response was directed against the foreign human proteins (CYP2D6 and FTCD), a molecular mimicry with murine homologous proteins led to the development of an autoimmune response against the antigens. These mice developed anti-LKM1 and anti-LC1 autoantibodies and an AIH which shows striking similarities with human type 2 AIH[47]. This murine model of type 2 AIH proved that exposure to a foreign protein can break immunological tolerance against a hepatic self-protein and this, without prior liver damage. The fact that a hepatitis is not necessary to break the immunological tolerance towards liver antigens argues in favor of non-hepatotropic virus(es) being able to trigger AIH in humans.
In patients, links between specific viruses and an autoimmune disease are difficult to establish in part due to the hit-and-run effect. The triggering viral infection could have been cleared months or even years before clinical signs of an autoimmune disorder become apparent. The identification of a virus as a causative agent for AIH must therefore rely on epidemiological studies to establish links between a specific infection and the autoimmune disease. A major obstacle in the elaboration of these studies is the necessity of very large cohorts of patients. Since AIH is a disease of very low prevalence, these studies are very difficult, if not impossible, to perform. Therefore, current candidate viruses as putative causative agent for AIH result from published case reports and homologies between liver autoantigens and virus proteins (Table 3).
Table 3.
Putative candidate viruses as triggering event in autoimmune hepatitis
| Putative virus as AIH trigger event | Evidences | References |
| Hepatitis A virus | Case reports | [94–96] |
| Hepatitis B virus | Case reports | [97,98] |
| Hepatitis C virus | Specific AIH autoantibodies | [87–89] |
| Cross-Reactivity at T and B-cell level | [90–92] | |
| Epstein-Barr virus | Case reports | [66–69] |
| Human herpes virus 6 | Sequence similarities | [99] |
| Herpes simplex virus | Sequence similarities | [102] |
| Cross-reactivity at B-cell level | [102] |
The hepatitis C virus (HCV) is probably the most studied infection present in AIH patients. Five to 10% of HCV infected patients show autoimmune features that are generally associated with AIH such as anti-LKM1, anti-LC1 and/or anti-SLA autoantibodies[87–89]. Purified anti-LKM1 antibodies from HCV-infected patients cross-reacted with the NS3 and NS5a purified proteins suggesting that CYP2D6 and those viral proteins shared similar structures[90]. A molecular mimicry at the B-cell level between a structural motif of CYP2D6 and HCV proteins could explain the production of anti-LKM1 antibodies in HCV-infected patients[90,91]. Another link between P450 and HCV was found at the T cell level by Kammer et al[92] who reported T cell cross-reactivity between P450 and the HCV core protein. Although HCV infection in some patients elicits autoimmune hepatitis-like immune responses through molecular mimicry mechanisms, HCV infection is not the triggering event in AIH patients. In fact, a study showed that very few patients with AIH had specific antibodies against HCV suggesting that most of them had never encountered this virus[93].
Other hepatitis-causing viruses such as hepatitis A and B viruses (HAV, HBV) have been proposed as triggers for AIH[94–98]. The Epstein-Barr virus (EBV), has been associated with AIH in several case reports and in a clinical follow-up of 13 patients[66–69]. These associations occurred in type 1 AIH patients; therefore, no molecular mimicry could be established since no specific autoantigens in type 1 AIH were identified. However, specific markers for these viral infections are not found in the majority of AIH patients.
In type 2 AIH, a putative molecular mimicry was found between two B cell epitopes of FTCD, the target of anti-LC1 autoantibodies, and sequences of the 101K antigenic virion protein and U50 protein from human herpes virus type 6 (HHV-6)[99]. Several homologies were also found between known epitopes of CYP2D6, targeted by anti-LKM1 antibodies, and proteins from HHV-6[99]. HHV-6 hepatotropism and its association with chronic and autoimmune hepatitis in children[100,101] makes this virus a plausible candidate for AIH onset.
A molecular mimicry has also been proposed between the main antigenic site of anti-LKM1 on CYP2D6 and herpes simplex virus (HSV-1) based on similarities between proteins sequences[102]. Cross-reactivity has been found between CYP2D6 and a HSV-1 protein using purified anti-LKM1 autoantibodies from an AIH patient[102]. However, in this study, 4 out 20 patients had not encountered HSV-1 prior to the development of autoimmunity as determined by an antibody assay against HSV-1[102]. These data suggest that HSV-1 could be a trigger\for the development of AIH in some patients, although this remains to be confirmed.
Modulation of gene expression: the role of the innate immune system in autoimmunity
Environmental factors could trigger an autoimmune reaction by creating a pro-inflammatory immunological micro-environment in which autoantigens could be presented. Such a pro-inflammatory micro-environment could result from toll-like receptor (TLR) engagement. Members of the toll-like family of receptors are able to bind pathogen-associated molecular patterns (PAMP) present in most pathogens. By doing so, TLRs can induce a quick and efficient response against those pathogens through the up-regulation of key pro-inflammatory genes, such as type 1 interferons. When the liver undergoes an infection, TLR stimulation could result in the presentation of autoantigens in a pro-inflammatory environment which would result in an efficient activation of specific autoreactive cells.
The role of TLRs in autoimmune liver disease has been studied in PBC. It was shown that patients with PBC have higher levels of TLR3, TLR4 and TLR9 receptors in the liver[103,104]. In vitro stimulation of monocytes from PBC patients with several TLR-binding molecules resulted in higher levels of cytokine secretion[105]. PBMCs from PBC patients, when cultivated with CpG, a TLR9 stimulator, secreted more IgM suggesting a role for TLR9 stimulation in the hyper-IgM observed in PBC patients[106]. Altogether, these studies suggest that TLRs could be involved in the pathogenesis of this autoimmune liver disease.
A break of immune tolerance towards the liver in a mouse model has been achieved by repeated CpG injections into a double-transgenic mouse expressing MHC class I molecule H-2Kb exclusively on hepatocytes and having T cells bearing specific TCR for MHC H-2Kb[107]. CpG injections were sufficient to break immune tolerance and induce a transient AIH, which faded when CpG injections were stopped. These data suggest that TLR9 stimulation is sufficient to activate pre-existing autoreactive T cells and initiate an autoimmune response, but is not sufficient to induce a self-perpetuating autoimmune response. Recently, Lang et al[108] were able to induce a liver inflammation by transferring liver-antigen specific CD8+ T cells in combination with TLR9 and TLR3 stimulation. These results led to the speculation that the immunoprivileged status of the liver could be controlled by TLR signaling[108]. Altogether these data suggest that the innate immune system could be involved in the development of autoimmune processes in the liver possibly through expression up-regulation of pro-inflammatory genes.
CONCLUSION
The pathogenesis of AIH is a complex process. Development of this disease requires a series of events (viral infection and/or chemical exposure) in a suitable environment (genetic background of susceptibility, female sex and young age) (Figure 1). Further research, both clinical and fundamental, will be needed before the pathogenesis of AIH is fully understood. A better comprehension of the disease would allow the development of specific immunotherapies with fewer side effects.
Peer reviewer: Dr. Stefan Wirth, Professor, Children’s Hospital, Heusnerstt. 40, Wuppertal 42349, Germany
S- Editor Li LF L- Editor Negro F E- Editor Ma WH
References
- 1.Reuben A. A sheep in wolf’s clothing. Hepatology. 2003;38:1596–1601. doi: 10.1002/hep.510380643. [DOI] [PubMed] [Google Scholar]
- 2.Manns MP, Vogel A. Autoimmune hepatitis, from mechanisms to therapy. Hepatology. 2006;43:S132–S144. doi: 10.1002/hep.21059. [DOI] [PubMed] [Google Scholar]
- 3.Boberg KM. Prevalence and epidemiology of autoimmune hepatitis. Clin Liver Dis. 2002;6:635–647. doi: 10.1016/s1089-3261(02)00021-1. [DOI] [PubMed] [Google Scholar]
- 4.Alvarez F, Berg PA, Bianchi FB, Bianchi L, Burroughs AK, Cancado EL, Chapman RW, Cooksley WG, Czaja AJ, Desmet VJ, et al. International Autoimmune Hepatitis Group Report: review of criteria for diagnosis of autoimmune hepatitis. J Hepatol. 1999;31:929–938. doi: 10.1016/s0168-8278(99)80297-9. [DOI] [PubMed] [Google Scholar]
- 5.Hennes EM, Zeniya M, Czaja AJ, Pares A, Dalekos GN, Krawitt EL, Bittencourt PL, Porta G, Boberg KM, Hofer H, et al. Simplified criteria for the diagnosis of autoimmune hepatitis. Hepatology. 2008;48:169–176. doi: 10.1002/hep.22322. [DOI] [PubMed] [Google Scholar]
- 6.Maggiore G, Veber F, Bernard O, Hadchouel M, Homberg JC, Alvarez F, Hadchouel P, Alagille D. Autoimmune hepatitis associated with anti-actin antibodies in children and adolescents. J Pediatr Gastroenterol Nutr. 1993;17:376–381. doi: 10.1097/00005176-199311000-00007. [DOI] [PubMed] [Google Scholar]
- 7.Vitozzi S, Djilali-Saiah I, Lapierre P, Alvarez F. Anti-soluble liver antigen/liver-pancreas (SLA/LP) antibodies in pediatric patients with autoimmune hepatitis. Autoimmunity. 2002;35:485–492. doi: 10.1080/0891693021000056712. [DOI] [PubMed] [Google Scholar]
- 8.Maggiore G, Bernard O, Homberg JC, Hadchouel M, Alvarez F, Hadchouel P, Odievre M, Alagille D. Liver disease associated with anti-liver-kidney microsome antibody in children. J Pediatr. 1986;108:399–404. doi: 10.1016/s0022-3476(86)80880-0. [DOI] [PubMed] [Google Scholar]
- 9.Lapierre P, Hajoui O, Homberg JC, Alvarez F. Formimino-transferase cyclodeaminase is an organ-specific autoantigen recognized by sera of patients with autoimmune hepatitis. Gastroenterology. 1999;116:643–649. doi: 10.1016/s0016-5085(99)70186-1. [DOI] [PubMed] [Google Scholar]
- 10.Gueguen M, Yamamoto AM, Bernard O, Alvarez F. Anti-liver-kidney microsome antibody type 1 recognizes human cytochrome P450 db1. Biochem Biophys Res Commun. 1989;159:542–547. doi: 10.1016/0006-291x(89)90027-2. [DOI] [PubMed] [Google Scholar]
- 11.Alvarez F. Autoimmune hepatitis. In: F Suchy, RJ Sokol, W Baliestreri., editors. Liver disease in childhood. Williams and Wilkins: Lippincott; 2001. pp. 429–441. [Google Scholar]
- 12.Gregorio GV, Portmann B, Reid F, Donaldson PT, Doherty DG, McCartney M, Mowat AP, Vergani D, Mieli-Vergani G. Autoimmune hepatitis in childhood: a 20-year experience. Hepatology. 1997;25:541–547. doi: 10.1002/hep.510250308. [DOI] [PubMed] [Google Scholar]
- 13.Krawitt EL. Autoimmune hepatitis. N Engl J Med. 2006;354:54–66. doi: 10.1056/NEJMra050408. [DOI] [PubMed] [Google Scholar]
- 14.Manns MP, Kruger M. Immunogenetics of chronic liver diseases. Gastroenterology. 1994;106:1676–1697. doi: 10.1016/0016-5085(94)90427-8. [DOI] [PubMed] [Google Scholar]
- 15.Doherty DG, Donaldson PT, Underhill JA, Farrant JM, Duthie A, Mieli-Vergani G, McFarlane IG, Johnson PJ, Eddleston AL, Mowat AP. Allelic sequence variation in the HLA class II genes and proteins in patients with autoimmune hepatitis. Hepatology. 1994;19:609–615. doi: 10.1002/hep.1840190311. [DOI] [PubMed] [Google Scholar]
- 16.Djilali-Saiah I, Renous R, Caillat-Zucman S, Debray D, Alvarez F. Linkage disequilibrium between HLA class II region and autoimmune hepatitis in pediatric patients. J Hepatol. 2004;40:904–909. doi: 10.1016/j.jhep.2004.02.023. [DOI] [PubMed] [Google Scholar]
- 17.Czaja AJ, Carpenter HA, Moore SB. Clinical and HLA phenotypes of type 1 autoimmune hepatitis in North American patients outside DR3 and DR4. Liver Int. 2006;26:552–558. doi: 10.1111/j.1478-3231.2006.01249.x. [DOI] [PubMed] [Google Scholar]
- 18.Vazquez-Garcia MN, Alaez C, Olivo A, Debaz H, Perez-Luque E, Burguete A, Cano S, de la Rosa G, Bautista N, Hernandez A, et al. MHC class II sequences of susceptibility and protection in Mexicans with autoimmune hepatitis. J Hepatol. 1998;28:985–990. doi: 10.1016/s0168-8278(98)80347-4. [DOI] [PubMed] [Google Scholar]
- 19.Pando M, Larriba J, Fernandez GC, Fainboim H, Ciocca M, Ramonet M, Badia I, Daruich J, Findor J, Tanno H, et al. Pediatric and adult forms of type 1 autoimmune hepatitis in Argentina: evidence for differential genetic predisposition. Hepatology. 1999;30:1374–1380. doi: 10.1002/hep.510300611. [DOI] [PubMed] [Google Scholar]
- 20.Seki T, Ota M, Furuta S, Fukushima H, Kondo T, Hino K, Mizuki N, Ando A, Tsuji K, Inoko H. HLA class II molecules and autoimmune hepatitis susceptibility in Japanese patients. Gastroenterology. 1992;103:1041–1047. doi: 10.1016/0016-5085(92)90041-v. [DOI] [PubMed] [Google Scholar]
- 21.Fainboim L, Marcos Y, Pando M, Capucchio M, Reyes GB, Galoppo C, Badia I, Remondino G, Ciocca M, Ramonet M. Chronic active autoimmune hepatitis in children. Strong association with a particular HLA-DR6 (DRB1*1301) haplotype. Hum Immunol. 1994;41:146–150. doi: 10.1016/0198-8859(94)90008-6. [DOI] [PubMed] [Google Scholar]
- 22.Czaja AJ, Souto EO, Bittencourt PL, Cancado EL, Porta G, Goldberg AC, Donaldson PT. Clinical distinctions and pathogenic implications of type 1 autoimmune hepatitis in Brazil and the United States. J Hepatol. 2002;37:302–308. doi: 10.1016/s0168-8278(02)00182-4. [DOI] [PubMed] [Google Scholar]
- 23.Jurado A, Cardaba B, Jara P, Cuadrado P, Hierro L, de Andres B, del Pozo V, Cortegano MI, Gallardo S, Camarena C, et al. Autoimmune hepatitis type 2 and hepatitis C virus infection: study of HLA antigens. J Hepatol. 1997;26:983–991. doi: 10.1016/s0168-8278(97)80106-7. [DOI] [PubMed] [Google Scholar]
- 24.Bittencourt PL, Goldberg AC, Cancado EL, Porta G, Carrilho FJ, Farias AQ, Palacios SA, Chiarella JM, Abrantes-Lemos CP, Baggio VL, et al. Genetic heterogeneity in susceptibility to autoimmune hepatitis types 1 and 2. Am J Gastroenterol. 1999;94:1906–1913. doi: 10.1111/j.1572-0241.1999.01229.x. [DOI] [PubMed] [Google Scholar]
- 25.Czaja AJ, Kruger M, Santrach PJ, Moore SB, Manns MP. Genetic distinctions between types 1 and 2 autoimmune hepatitis. Am J Gastroenterol. 1997;92:2197–2200. [PubMed] [Google Scholar]
- 26.Strettell MD, Donaldson PT, Thomson LJ, Santrach PJ, Moore SB, Czaja AJ, Williams R. Allelic basis for HLA-encoded susceptibility to type 1 autoimmune hepatitis. Gastroenterology. 1997;112:2028–2035. doi: 10.1053/gast.1997.v112.pm9178696. [DOI] [PubMed] [Google Scholar]
- 27.Djilali-Saiah I, Fakhfakh A, Louafi H, Caillat-Zucman S, Debray D, Alvarez F. HLA class II influences humoral autoimmunity in patients with type 2 autoimmune hepatitis. J Hepatol. 2006;45:844–850. doi: 10.1016/j.jhep.2006.07.034. [DOI] [PubMed] [Google Scholar]
- 28.De la Concha EG, Fernandez-Arquero M, Gual L, Vigil P, Martinez A, Urcelay E, Ferreira A, Garcia-Rodriguez MC, Fontan G. MHC susceptibility genes to IgA deficiency are located in different regions on different HLA haplotypes. J Immunol. 2002;169:4637–4643. doi: 10.4049/jimmunol.169.8.4637. [DOI] [PubMed] [Google Scholar]
- 29.Vorechovsky I, Webster AD, Plebani A, Hammarstrom L. Genetic linkage of IgA deficiency to the major histocompatibility complex: evidence for allele segregation distortion, parent-of-origin penetrance differences, and the role of anti-IgA antibodies in disease predisposition. Am J Hum Genet. 1999;64:1096–1109. doi: 10.1086/302326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Vergani D, Wells L, Larcher VF, Nasaruddin BA, Davies ET, Mieli-Vergani G, Mowat AP. Genetically determined low C4: a predisposing factor to autoimmune chronic active hepatitis. Lancet. 1985;2:294–298. doi: 10.1016/s0140-6736(85)90348-4. [DOI] [PubMed] [Google Scholar]
- 31.Scully LJ, Toze C, Sengar DP, Goldstein R. Early-onset autoimmune hepatitis is associated with a C4A gene deletion. Gastroenterology. 1993;104:1478–1484. doi: 10.1016/0016-5085(93)90359-k. [DOI] [PubMed] [Google Scholar]
- 32.Djilali-Saiah I, Larger E, Harfouch-Hammoud E, Timsit J, Clerc J, Bertin E, Assan R, Boitard C, Bach JF, Caillat-Zucman S. No major role for the CTLA-4 gene in the association of autoimmune thyroid disease with IDDM. Diabetes. 1998;47:125–127. doi: 10.2337/diab.47.1.125. [DOI] [PubMed] [Google Scholar]
- 33.Fukazawa T, Yanagawa T, Kikuchi S, Yabe I, Sasaki H, Hamada T, Miyasaka K, Gomi K, Tashiro K. CTLA-4 gene polymorphism may modulate disease in Japanese multiple sclerosis patients. J Neurol Sci. 1999;171:49–55. doi: 10.1016/s0022-510x(99)00251-8. [DOI] [PubMed] [Google Scholar]
- 34.Djilali-Saiah I, Schmitz J, Harfouch-Hammoud E, Mougenot JF, Bach JF, Caillat-Zucman S. CTLA-4 gene polymorphism is associated with predisposition to coeliac disease. Gut. 1998;43:187–189. doi: 10.1136/gut.43.2.187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Agarwal K, Czaja AJ, Jones DE, Donaldson PT. Cytotoxic T lymphocyte antigen-4 (CTLA-4) gene polymorphisms and susceptibility to type 1 autoimmune hepatitis. Hepatology. 2000;31:49–53. doi: 10.1002/hep.510310110. [DOI] [PubMed] [Google Scholar]
- 36.Djilali-Saiah I, Ouellette P, Caillat-Zucman S, Debray D, Kohn JI, Alvarez F. CTLA-4/CD 28 region polymorphisms in children from families with autoimmune hepatitis. Hum Immunol. 2001;62:1356–1362. doi: 10.1016/s0198-8859(01)00344-5. [DOI] [PubMed] [Google Scholar]
- 37.Hiraide A, Imazeki F, Yokosuka O, Kanda T, Kojima H, Fukai K, Suzuki Y, Hata A, Saisho H. Fas polymorphisms influence susceptibility to autoimmune hepatitis. Am J Gastroenterol. 2005;100:1322–1329. doi: 10.1111/j.1572-0241.2005.41053.x. [DOI] [PubMed] [Google Scholar]
- 38.Agarwal K, Czaja AJ, Donaldson PT. A functional Fas promoter polymorphism is associated with a severe phenotype in type 1 autoimmune hepatitis characterized by early development of cirrhosis. Tissue Antigens. 2007;69:227–235. doi: 10.1111/j.1399-0039.2006.00794.x. [DOI] [PubMed] [Google Scholar]
- 39.Vogel A, Strassburg CP, Manns MP. Genetic association of vitamin D receptor polymorphisms with primary biliary cirrhosis and autoimmune hepatitis. Hepatology. 2002;35:126–131. doi: 10.1053/jhep.2002.30084. [DOI] [PubMed] [Google Scholar]
- 40.Rook GA, Steele J, Ainsworth M, Champion BR. Activation of macrophages to inhibit proliferation of Mycobacterium tuberculosis: comparison of the effects of recombinant gamma-interferon on human monocytes and murine peritoneal macrophages. Immunology. 1986;59:333–338. [PMC free article] [PubMed] [Google Scholar]
- 41.Lemire JM, Archer DC, Beck L, Spiegelberg HL. Immunosuppressive actions of 1,25-dihydroxyvitamin D3: preferential inhibition of Th1 functions. J Nutr. 1995;125:1704S–1708S. doi: 10.1093/jn/125.suppl_6.1704S. [DOI] [PubMed] [Google Scholar]
- 42.Berer A, Stockl J, Majdic O, Wagner T, Kollars M, Lechner K, Geissler K, Oehler L. 1,25-Dihydroxyvitamin D(3) inhibits dendritic cell differentiation and maturation in vitro. Exp Hematol. 2000;28:575–583. doi: 10.1016/s0301-472x(00)00143-0. [DOI] [PubMed] [Google Scholar]
- 43.Cookson S, Constantini PK, Clare M, Underhill JA, Bernal W, Czaja AJ, Donaldson PT. Frequency and nature of cytokine gene polymorphisms in type 1 autoimmune hepatitis. Hepatology. 1999;30:851–856. doi: 10.1002/hep.510300412. [DOI] [PubMed] [Google Scholar]
- 44.Czaja AJ, Cookson S, Constantini PK, Clare M, Underhill JA, Donaldson PT. Cytokine polymorphisms associated with clinical features and treatment outcome in type 1 autoimmune hepatitis. Gastroenterology. 1999;117:645–652. doi: 10.1016/s0016-5085(99)70458-0. [DOI] [PubMed] [Google Scholar]
- 45.Ahonen P, Myllarniemi S, Sipila I, Perheentupa J. Clinical variation of autoimmune polyendocrinopathy-candidiasis-ectodermal dystrophy (APECED) in a series of 68 patients. N Engl J Med. 1990;322:1829–1836. doi: 10.1056/NEJM199006283222601. [DOI] [PubMed] [Google Scholar]
- 46.Vogel A, Liermann H, Harms A, Strassburg CP, Manns MP, Obermayer-Straub P. Autoimmune regulator AIRE: evidence for genetic differences between autoimmune hepatitis and hepatitis as part of the autoimmune polyglandular syndrome type 1. Hepatology. 2001;33:1047–1052. doi: 10.1053/jhep.2001.24031. [DOI] [PubMed] [Google Scholar]
- 47.Lapierre P, Djilali-Saiah I, Vitozzi S, Alvarez F. A murine model of type 2 autoimmune hepatitis: Xenoimmunization with human antigens. Hepatology. 2004;39:1066–1074. doi: 10.1002/hep.20109. [DOI] [PubMed] [Google Scholar]
- 48.Lapierre P, Beland K, Djilali-Saiah I, Alvarez F. Type 2 autoimmune hepatitis murine model: the influence of genetic background in disease development. J Autoimmun. 2006;26:82–89. doi: 10.1016/j.jaut.2005.11.001. [DOI] [PubMed] [Google Scholar]
- 49.Whitacre CC. Sex differences in autoimmune disease. Nat Immunol. 2001;2:777–780. doi: 10.1038/ni0901-777. [DOI] [PubMed] [Google Scholar]
- 50.Chu CM, Sheen IS, Lin SM, Liaw YF. Sex difference in chronic hepatitis B virus infection: studies of serum HBeAg and alanine aminotransferase levels in 10,431 asymptomatic Chinese HBsAg carriers. Clin Infect Dis. 1993;16:709–713. doi: 10.1093/clind/16.5.709. [DOI] [PubMed] [Google Scholar]
- 51.Brooks BK, Levy MF, Jennings LW, Abbasoglu O, Vodapally M, Goldstein RM, Husberg BS, Gonwa TA, Klintmalm GB. Influence of donor and recipient gender on the outcome of liver transplantation. Transplantation. 1996;62:1784–1787. doi: 10.1097/00007890-199612270-00017. [DOI] [PubMed] [Google Scholar]
- 52.Michaels RM, Rogers KD. A sex difference in immunologic responsiveness. Pediatrics. 1971;47:120–123. [PubMed] [Google Scholar]
- 53.Amadori A, Zamarchi R, De Silvestro G, Forza G, Cavatton G, Danieli GA, Clementi M, Chieco-Bianchi L. Genetic control of the CD4/CD8 T-cell ratio in humans. Nat Med. 1995;1:1279–1283. doi: 10.1038/nm1295-1279. [DOI] [PubMed] [Google Scholar]
- 54.Araneo BA, Dowell T, Diegel M, Daynes RA. Dihydrotesto-sterone exerts a depressive influence on the production of interleukin-4 (IL-4), IL-5, and gamma-interferon, but not IL-2 by activated murine T cells. Blood. 1991;78:688–699. [PubMed] [Google Scholar]
- 55.Keating JJ, O’Brien CJ, Stellon AJ, Portmann BC, Johnson RD, Johnson PJ, Williams R. Influence of aetiology, clinical and histological features on survival in chronic active hepatitis: an analysis of 204 patients. Q J Med. 1987;62:59–66. [PubMed] [Google Scholar]
- 56.McMurray RW, Ndebele K, Hardy KJ, Jenkins JK. 17-beta-estradiol suppresses IL-2 and IL-2 receptor. Cytokine. 2001;14:324–333. doi: 10.1006/cyto.2001.0900. [DOI] [PubMed] [Google Scholar]
- 57.Polanczyk M, Zamora A, Subramanian S, Matejuk A, Hess DL, Blankenhorn EP, Teuscher C, Vandenbark AA, Offner H. The protective effect of 17beta-estradiol on experimental autoimmune encephalomyelitis is mediated through estrogen receptor-alpha. Am J Pathol. 2003;163:1599–1605. doi: 10.1016/s0002-9440(10)63516-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Delpy L, Douin-Echinard V, Garidou L, Bruand C, Saoudi A, Guery JC. Estrogen enhances susceptibility to experimental autoimmune myasthenia gravis by promoting type 1-polarized immune responses. J Immunol. 2005;175:5050–5057. doi: 10.4049/jimmunol.175.8.5050. [DOI] [PubMed] [Google Scholar]
- 59.Buggage RR, Matteson DM, Shen DF, Sun B, Tuaillon N, Chan CC. Effect of sex hormones on experimental autoimmune uveoretinitis (EAU) Immunol Invest. 2003;32:259–273. doi: 10.1081/imm-120025105. [DOI] [PubMed] [Google Scholar]
- 60.Roubinian JR, Talal N, Greenspan JS, Goodman JR, Siiteri PK. Effect of castration and sex hormone treatment on survival, anti-nucleic acid antibodies, and glomerulonephritis in NZB/NZW F1 mice. J Exp Med. 1978;147:1568–1583. doi: 10.1084/jem.147.6.1568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Liva SM, Voskuhl RR. Testosterone acts directly on CD4+ T lymphocytes to increase IL-10 production. J Immunol. 2001;167:2060–2067. doi: 10.4049/jimmunol.167.4.2060. [DOI] [PubMed] [Google Scholar]
- 62.Dalal M, Kim S, Voskuhl RR. Testosterone therapy ameliorates experimental autoimmune encephalomyelitis and induces a T helper 2 bias in the autoantigen-specific T lymphocyte response. J Immunol. 1997;159:3–6. [PubMed] [Google Scholar]
- 63.Heneghan MA, Norris SM, O’Grady JG, Harrison PM, McFarlane IG. Management and outcome of pregnancy in autoimmune hepatitis. Gut. 2001;48:97–102. doi: 10.1136/gut.48.1.97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Buchel E, Van Steenbergen W, Nevens F, Fevery J. Improvement of autoimmune hepatitis during pregnancy followed by flare-up after delivery. Am J Gastroenterol. 2002;97:3160–3165. doi: 10.1111/j.1572-0241.2002.07124.x. [DOI] [PubMed] [Google Scholar]
- 65.Samuel D, Riordan S, Strasser S, Kurtovic J, Singh-Grewel I, Koorey D. Severe autoimmune hepatitis first presenting in the early post partum period. Clin Gastroenterol Hepatol. 2004;2:622–624. doi: 10.1016/s1542-3565(04)00245-9. [DOI] [PubMed] [Google Scholar]
- 66.Aceti A, Mura MS, Babudieri S, Bacciu SA. A young woman with hepatitis after a sore throat. Lancet. 1995;346:1603. doi: 10.1016/s0140-6736(95)91932-5. [DOI] [PubMed] [Google Scholar]
- 67.Vento S, Guella L, Mirandola F, Cainelli F, Di Perri G, Solbiati M, Ferraro T, Concia E. Epstein-Barr virus as a trigger for autoimmune hepatitis in susceptible individuals. Lancet. 1995;346:608–609. doi: 10.1016/s0140-6736(95)91438-2. [DOI] [PubMed] [Google Scholar]
- 68.Kojima K, Nagayama R, Hirama S, Maeda T, Takikawa H, Miyake K, Yamanaka M, Shiga J. Epstein-Barr virus infection resembling autoimmune hepatitis with lactate dehydrogenase and alkaline phosphatase anomaly. J Gastroenterol. 1999;34:706–712. doi: 10.1007/s005350050324. [DOI] [PubMed] [Google Scholar]
- 69.Nobili V, Comparcola D, Sartorelli MR, Devito R, Marcellini M. Autoimmune hepatitis type 1 after Epstein-Barr virus infection. Pediatr Infect Dis J. 2003;22:387. doi: 10.1097/01.inf.0000060825.68086.9c. [DOI] [PubMed] [Google Scholar]
- 70.Tiegs G, Hentschel J, Wendel A. A T cell-dependent experimental liver injury in mice inducible by concanavalin A. J Clin Invest. 1992;90:196–203. doi: 10.1172/JCI115836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Kusters S, Gantner F, Kunstle G, Tiegs G. Interferon gamma plays a critical role in T cell-dependent liver injury in mice initiated by concanavalin A. Gastroenterology. 1996;111:462–471. doi: 10.1053/gast.1996.v111.pm8690213. [DOI] [PubMed] [Google Scholar]
- 72.Keren Z, Berke G. Selective binding of concanavalin A to target cell major histocompatibility antigens is required to induce nonspecific conjugation and lysis by cytolytic T lymphocytes in lectin-dependent cytotoxicity. Cell Immunol. 1984;89:458–477. doi: 10.1016/0008-8749(84)90347-2. [DOI] [PubMed] [Google Scholar]
- 73.Teitelbaum JE, Perez-Atayde AR, Cohen M, Bousvaros A, Jonas MM. Minocycline-related autoimmune hepatitis: case series and literature review. Arch Pediatr Adolesc Med. 1998;152:1132–1136. doi: 10.1001/archpedi.152.11.1132. [DOI] [PubMed] [Google Scholar]
- 74.Gough A, Chapman S, Wagstaff K, Emery P, Elias E. Minocycline induced autoimmune hepatitis and systemic lupus erythematosus-like syndrome. BMJ. 1996;312:169–172. doi: 10.1136/bmj.312.7024.169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Nietsch HH, Libman BS, Pansze TW, Eicher JN, Reeves JR, Krawitt EL. Minocycline-induced hepatitis. Am J Gastroenterol. 2000;95:2993–2995. doi: 10.1111/j.1572-0241.2000.03222.x. [DOI] [PubMed] [Google Scholar]
- 76.Cohen SM, O’Connor AM, Hart J, Merel NH, Te HS. Autoimmune hepatitis associated with the use of black cohosh: a case study. Menopause. 2004;11:575–577. doi: 10.1097/01.gme.0000142914.55849.6a. [DOI] [PubMed] [Google Scholar]
- 77.Kamiyama T, Nouchi T, Kojima S, Murata N, Ikeda T, Sato C. Autoimmune hepatitis triggered by administration of an herbal medicine. Am J Gastroenterol. 1997;92:703–704. [PubMed] [Google Scholar]
- 78.Alla V, Abraham J, Siddiqui J, Raina D, Wu GY, Chalasani NP, Bonkovsky HL. Autoimmune hepatitis triggered by statins. J Clin Gastroenterol. 2006;40:757–761. doi: 10.1097/00004836-200609000-00018. [DOI] [PubMed] [Google Scholar]
- 79.Leung PS, Park O, Tsuneyama K, Kurth MJ, Lam KS, Ansari AA, Coppel RL, Gershwin ME. Induction of primary biliary cirrhosis in guinea pigs following chemical xenobiotic immunization. J Immunol. 2007;179:2651–2657. doi: 10.4049/jimmunol.179.4.2651. [DOI] [PubMed] [Google Scholar]
- 80.Herkel J, Jagemann B, Wiegard C, Lazaro JF, Lueth S, Kanzler S, Blessing M, Schmitt E, Lohse AW. MHC class II-expressing hepatocytes function as antigen-presenting cells and activate specific CD4 T lymphocyutes. Hepatology. 2003;37:1079–1085. doi: 10.1053/jhep.2003.50191. [DOI] [PubMed] [Google Scholar]
- 81.Yang R, Liu Q, Grosfeld JL, Pescovitz MD. Intestinal venous drainage through the liver is a prerequisite for oral tolerance induction. J Pediatr Surg. 1994;29:1145–1148. doi: 10.1016/0022-3468(94)90297-6. [DOI] [PubMed] [Google Scholar]
- 82.Wiegard C, Wolint P, Frenzel C, Cheruti U, Schmitt E, Oxenius A, Lohse AW, Herkel J. Defective T helper response of hepatocyte-stimulated CD4 T cells impairs antiviral CD8 response and viral clearance. Gastroenterology. 2007;133:2010–2018. doi: 10.1053/j.gastro.2007.09.007. [DOI] [PubMed] [Google Scholar]
- 83.Bowen DG, McCaughan GW, Bertolino P. Intrahepatic immunity: a tale of two sites? Trends Immunol. 2005;26:512–517. doi: 10.1016/j.it.2005.08.005. [DOI] [PubMed] [Google Scholar]
- 84.Bowen DG, Zen M, Holz L, Davis T, McCaughan GW, Bertolino P. The site of primary T cell activation is a determinant of the balance between intrahepatic tolerance and immunity. J Clin Invest. 2004;114:701–712. doi: 10.1172/JCI21593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Djilali-Saiah I, Lapierre P, Vittozi S, Alvarez F. DNA vaccination breaks tolerance for a neo-self antigen in liver: a transgenic murine model of autoimmune hepatitis. J Immunol. 2002;169:4889–4896. doi: 10.4049/jimmunol.169.9.4889. [DOI] [PubMed] [Google Scholar]
- 86.Albert LJ, Inman RD. Molecular mimicry and autoimmunity. N Engl J Med. 1999;341:2068–2074. doi: 10.1056/NEJM199912303412707. [DOI] [PubMed] [Google Scholar]
- 87.Lenzi M, Ballardini G, Fusconi M, Cassani F, Selleri L, Volta U, Zauli D, Bianchi FB. Type 2 autoimmune hepatitis and hepatitis C virus infection. Lancet. 1990;335:258–259. doi: 10.1016/0140-6736(90)90070-l. [DOI] [PubMed] [Google Scholar]
- 88.Beland K, Lapierre P, Marceau G, Alvarez F. Anti-LC1 autoantibodies in patients with chronic hepatitis C virus infection. J Autoimmun. 2004;22:159–166. doi: 10.1016/j.jaut.2003.11.001. [DOI] [PubMed] [Google Scholar]
- 89.Vitozzi S, Lapierre P, Djilali-Saiah I, Marceau G, Beland K, Alvarez F. Anti-soluble liver antigen (SLA) antibodies in chronic HCV infection. Autoimmunity. 2004;37:217–222. doi: 10.1080/08916930410001686260. [DOI] [PubMed] [Google Scholar]
- 90.Marceau G, Lapierre P, Beland K, Soudeyns H, Alvarez F. LKM1 autoantibodies in chronic hepatitis C infection: a case of molecular mimicry? Hepatology. 2005;42:675–682. doi: 10.1002/hep.20816. [DOI] [PubMed] [Google Scholar]
- 91.Kerkar N, Choudhuri K, Ma Y, Mahmoud A, Bogdanos DP, Muratori L, Bianchi F, Williams R, Mieli-Vergani G, Vergani D. Cytochrome P4502D6(193-212): a new immunodominant epitope and target of virus/self cross-reactivity in liver kidney microsomal autoantibody type 1-positive liver disease. J Immunol. 2003;170:1481–1489. doi: 10.4049/jimmunol.170.3.1481. [DOI] [PubMed] [Google Scholar]
- 92.Kammer AR, van der Burg SH, Grabscheid B, Hunziker IP, Kwappenberg KM, Reichen J, Melief CJ, Cerny A. Molecular mimicry of human cytochrome P450 by hepatitis C virus at the level of cytotoxic T cell recognition. J Exp Med. 1999;190:169–176. doi: 10.1084/jem.190.2.169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Czaja AJ, Carpenter HA, Santrach PJ, Moore SB, Taswell HF, Homburger HA. Evidence against hepatitis viruses as important causes of severe autoimmune hepatitis in the United States. J Hepatol. 1993;18:342–352. doi: 10.1016/s0168-8278(05)80279-x. [DOI] [PubMed] [Google Scholar]
- 94.Grunhage F, Spengler U, Fischer HP, Sauerbruch T. Autoimmune hepatitis--sequel of a relapsing hepatitis A in a 75-year-old woman. Digestion. 2004;70:187–191. doi: 10.1159/000082253. [DOI] [PubMed] [Google Scholar]
- 95.Huppertz HI, Treichel U, Gassel AM, Jeschke R, Meyer zum Buschenfelde KH. Autoimmune hepatitis following hepatitis A virus infection. J Hepatol. 1995;23:204–208. doi: 10.1016/0168-8278(95)80336-x. [DOI] [PubMed] [Google Scholar]
- 96.Skoog SM, Rivard RE, Batts KP, Smith CI. Autoimmune hepatitis preceded by acute hepatitis A infection. Am J Gastroenterol. 2002;97:1568–1569. doi: 10.1111/j.1572-0241.2002.05751.x. [DOI] [PubMed] [Google Scholar]
- 97.Murakami C, Hino K, Okazaki M, Fujii K, Okuda M, Hanada H, Yamasaki T, Okita K. Hepatitis B virus carrier status linked to autoimmune hepatitis. Intern Med. 1996;35:468–471. doi: 10.2169/internalmedicine.35.468. [DOI] [PubMed] [Google Scholar]
- 98.Maya R, Gershwin ME, Shoenfeld Y. Hepatitis B Virus (HBV) and Autoimmune Disease. Clin Rev Allergy Immunol. 2008;34:85–102. doi: 10.1007/s12016-007-8013-6. [DOI] [PubMed] [Google Scholar]
- 99.Lapierre P, Johanet C, Alvarez F. Characterization of the B cell response of patients with anti-liver cytosol autoantibodies in type 2 autoimmune hepatitis. Eur J Immunol. 2003;33:1869–1878. doi: 10.1002/eji.200323857. [DOI] [PubMed] [Google Scholar]
- 100.Tajiri H, Tanaka-Taya K, Ozaki Y, Okada S, Mushiake S, Yamanishi K. Chronic hepatitis in an infant, in association with human herpesvirus-6 infection. J Pediatr. 1997;131:473–475. doi: 10.1016/s0022-3476(97)80082-0. [DOI] [PubMed] [Google Scholar]
- 101.Schmitt K, Deutsch J, Tulzer G, Meindi R, Aberle S. Autoimmune hepatitis and adrenal insufficiency in an infant with human herpesvirus-6 infection. Lancet. 1996;348:966. doi: 10.1016/s0140-6736(05)65385-8. [DOI] [PubMed] [Google Scholar]
- 102.Manns MP, Griffin KJ, Sullivan KF, Johnson EF. LKM-1 autoantibodies recognize a short linear sequence in P450IID6, a cytochrome P-450 monooxygenase. J Clin Invest. 1991;88:1370–1378. doi: 10.1172/JCI115443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Takii Y, Nakamura M, Ito M, Yokoyama T, Komori A, Shimizu-Yoshida Y, Nakao R, Kusumoto K, Nagaoka S, Yano K, et al. Enhanced expression of type 1 interferon and toll-like receptor-3 in primary biliary cirrhosis. Lab Invest. 2005;85:908–920. doi: 10.1038/labinvest.3700285. [DOI] [PubMed] [Google Scholar]
- 104.Wang AP, Migita K, Ito M, Takii Y, Daikoku M, Yokoyama T, Komori A, Nakamura M, Yatsuhashi H, Ishibashi H. Hepatic expression of toll-like receptor 4 in primary biliary cirrhosis. J Autoimmun. 2005;25:85–91. doi: 10.1016/j.jaut.2005.05.003. [DOI] [PubMed] [Google Scholar]
- 105.Mao TK, Lian ZX, Selmi C, Ichiki Y, Ashwood P, Ansari AA, Coppel RL, Shimoda S, Ishibashi H, Gershwin ME. Altered monocyte responses to defined TLR ligands in patients with primary biliary cirrhosis. Hepatology. 2005;42:802–808. doi: 10.1002/hep.20859. [DOI] [PubMed] [Google Scholar]
- 106.Kikuchi K, Lian ZX, Yang GX, Ansari AA, Ikehara S, Kaplan M, Miyakawa H, Coppel RL, Gershwin ME. Bacterial CpG induces hyper-IgM production in CD27(+) memory B cells in primary biliary cirrhosis. Gastroenterology. 2005;128:304–312. doi: 10.1053/j.gastro.2004.11.005. [DOI] [PubMed] [Google Scholar]
- 107.Sacher T, Knolle P, Nichterlein T, Arnold B, Hammerling GJ, Limmer A. CpG-ODN-induced inflammation is sufficient to cause T-cell-mediated autoaggression against hepatocytes. Eur J Immunol. 2002;32:3628–3637. doi: 10.1002/1521-4141(200212)32:12<3628::AID-IMMU3628>3.0.CO;2-E. [DOI] [PubMed] [Google Scholar]
- 108.Lang KS, Georgiev P, Recher M, Navarini AA, Bergthaler A, Heikenwalder M, Harris NL, Junt T, Odermatt B, Clavien PA, et al. Immunoprivileged status of the liver is controlled by Toll-like receptor 3 signaling. J Clin Invest. 2006;116:2456–2463. doi: 10.1172/JCI28349. [DOI] [PMC free article] [PubMed] [Google Scholar]